不同年生凤丹根际AM真菌与土壤酶活性的相关性分析
2018-03-08李志雅杨安娜汤冬梅汪建中
李志雅, 杨安娜, 汤冬梅, 汪建中, 杨 帆
(安徽师范大学 生命科学学院 生物环境与生态安全安徽省高校省级重点实验室, 芜湖 241000)
凤丹(Paeoniaostii)是牡丹野生种杨山牡丹的栽培品种,主要应用于牡丹丹皮和籽油的生产,是一种很好的药食兼备植物[1]。其根皮有镇痛、解热、抗过敏、免疫等药用,而其籽油作为一种新型木本植物油,具有抗肿瘤、抗炎、改善心血管和调节免疫等医疗保健功能[2-5]。凤丹在安徽南陵已有1600多年的栽培历史,因其质地优良,逐渐成为优良道地药材,为药用牡丹中的主流品种[6],并享受地理标志产品保护。
AM是AM真菌和植物根系一起形成的互惠共生体,大约80%的陆生植物都能与AM真菌共生形成菌根,AM是自然界中分布最广泛的内生菌根[7]。丛枝是AM最重要的结构之一,是AM真菌与植物之间进行物质交换的场所。研究表明,AM真菌能够促进植物幼苗生长[8-10]和对矿质营养尤其是P的吸收,减少施肥量[11-14];提高药用植物有效成分的含量[15-19];增强药用植物的抗环境胁迫能力和抗病性,促进药用植物健康生长[20]。
土壤酶是土壤生态系统的重要组分之一,通常认为土壤酶在很大程度上起源于土壤微生物, 也有一部分来源于植物和土壤动物, 这些生物通过生理代谢向土壤分泌酶,并且它们的残体亦可溶出胞内酶进入土壤[21-22]。有研究表明,接种AM真菌后可以增强植物根围土壤磷酸酶、脲酶和蛋白酶活性[23-24],各种酶活性的增加幅度与宿主菌根侵染率显著相关。因此,根际土壤酶活性是土壤因子被植物有效利用的关键,是AM真菌活力的重要体现,研究凤丹根际AM真菌与土壤酶活性之间的关系,在一定程度上可以从根土界面揭示AM真菌对宿主营养吸收的贡献,为凤丹的栽培研究和道地保护提供理论依据。
1 材料与方法
1.1 样本采集与处理
所选样地为安徽省芜湖市南陵凤丹种植栽培基地(30°48.18.9′N,118°00.08.3′E,海拔80 m),是生产凤丹的主要地区之一。供试土壤样品选取自然条件基本一致,凤丹种植年限为2、3、4年的3个样田,每个样田采集4个样品,将根样放入有1/2的FAA固定液中,土样去除石子及腐败的落叶,然后封袋,带回实验室。将土样分成二份,一份湿土用于测定土壤的菌丝量,另一份自然风干,用于测定土壤中的孢子密度和土壤酶活性,并放入4℃冰箱中保存。
1.2 测定方法
根样用碱解离-酸性品红染色法[25]进行处理并制成封片,于OLYMPUS-BX51光学显微镜下观察AM真菌在各种植物根系上的侵染情况并估算定殖率[26],并对丛枝菌根典型形态结构如泡囊、丛枝、菌丝圈进行摄影。土样自然风干后,采用湿筛沉淀法[27]收集AM真菌孢子和孢子果,于体视镜下进行计数。将根部的皮剥下,采用HPLC法测定凤丹根部丹皮酚含量。
土壤酶活性测定均采用比色法,酸性磷酸酶、脲酶、蔗糖酶、多酚氧化酶的活性测定分别采用磷酸苯二钠比色法、苯酚—次氯酸钠比色法、3,5-二硝基水杨酸比色法、邻苯三酚比色法[28],在分光光度计下进行测定,读取吸光值。
1.3 数据分析中的参数
定殖率(colonization rate):指AM真菌在植物根中定殖的比率,H(%)是菌丝定殖率,A(%)是丛枝定殖率,HC(%)是菌丝圈定殖率,V(%)是泡囊定殖率,DSE(%)是黑格菌丝定殖率。
孢子密度(spore density,SD):指生物个体在单位面积中的数目,本文指20 g干土样中AM真菌孢子数。
菌丝量(hyphae quantity,HQ):指菌丝在单位面积中的个数,本文指5 g干土样中所含菌丝的比率,单位mm/g。
2 结果
2.1 凤丹根际AM真菌结构
通过对南陵丫山采集的凤丹根样进行染色制片与镜检,发现所采集的根样全部被AM真菌侵染,形成了Arum-type类型的AM结构,即菌丝在宿主皮层细胞间隙生长,产生大量的胞间菌丝,并且侧向分支进入细胞内,其顶端不断分叉而发育成典型的丛枝结构(图1-1、1-2)。在镜下可看到丛枝菌根真菌中的泡囊、菌丝圈、黑隔菌丝和根内孢子,其中当菌-植共生体生长达到顶峰时,菌丝的顶部开始膨大,从而形成了贮存器官泡囊结构(图1-3),泡囊有不同的形状,如卵圆形、椭圆形等;也有菌丝在细胞内或细胞间弯曲闭合形成了大量的菌丝圈结构(图1-6)[29];在个别菌根内还观察到了根内孢子结构(图1-5)。
2.2 凤丹根际黑隔菌丝结构
在光镜下,还可以看到一些菌丝没有被染液染上颜色,而且菌丝还被一小段一小段地隔开,显示出明显的隔断,称它为黑隔菌丝(dark septate endophytes,DSE)(图1-4),DSE是指定居在植物根组织细胞内或细胞间隙,在分类学上较为混杂的一群子囊菌和无性繁殖的真菌[30],它并不属于丛枝结构。
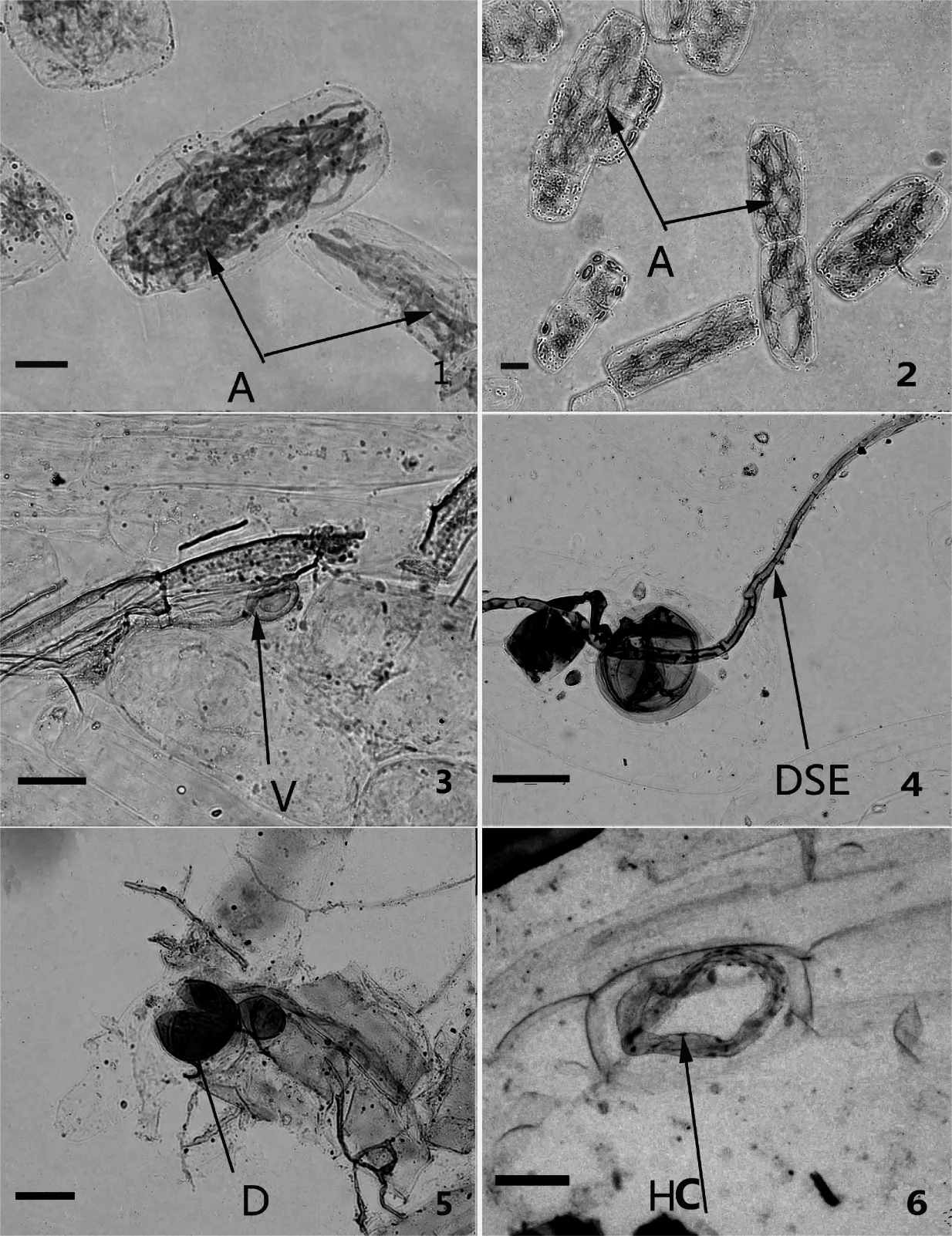
图1 凤丹根皮组织中AM真菌的定殖情况(1 bar = 30 μm)Fig 1 Structure of AMF in roots of Paeonia ostii(1 bar = 30 μm)
A:丛枝(arbuscules);V:泡囊(vesicules);DSE:黑隔菌丝(dark septate endophytes);D:孢子(spores);HC:菌丝圈(hyphal coil)
2.3 凤丹根际AM真菌的定殖状况
从定殖上看,不同年生的凤丹全部被侵染,但是定殖程度不同,其中两年生凤丹的丛枝定殖率最高,达32%;三年生凤丹的丛枝定殖率最低,为14.5%,丛枝定殖率在不同年生凤丹中的差异不明显(表1)。
2.4 凤丹根际土中孢子密度
实验结果发现,所有的样品中均有孢子产生,但不同年生凤丹根际土中孢子密度不同。从高到低为:2年生凤丹、4年生凤丹、3年生凤丹,且不同年生孢子密度之间差异不显著(表1)。
2.5 凤丹根际土壤中的菌丝含量
在研究的样品中,实验结果为3年生凤丹根际土的菌丝量最多,4年生的凤丹菌丝量最低,并且2年生、3年生、4年生的凤丹根际土中菌丝量没有差异(表1)。
2.6 凤丹根部丹皮酚含量
如图2所示,凤丹根部丹皮酚含量为3年生<4年生<2年生,其中2年生丹皮酚含量为0.174mg,且不同年生之间无显著差异。
2.7 凤丹根际土壤中酶活性变化
由实验可知,不同年生凤丹根际多酚氧化酶活性存在显著差异,其中,4年生凤丹活性最强,2年生凤丹活性最弱,表明其根际多酚氧化酶活性呈逐年增长的趋势(图3-b);不同年生凤丹根际蔗糖酶活性无规律性变化,2年生凤丹活性最强(图3-d);土壤酸性磷酸酶的活性随着凤丹种植时间的增加活性逐渐降低,且差异达到了显著水平(图3-a);在土壤脲酶活性测定中,3年生凤丹根际土壤活性最高,2年生最低,酶活在不同年生凤丹中无规律性变化,且4年生凤丹土壤脲酶活性在P<0.05水平上差异显著(图3-c)。
表1凤丹AM真菌的定殖率
Table1 Colonization an of AMF ofPaeoniaostii
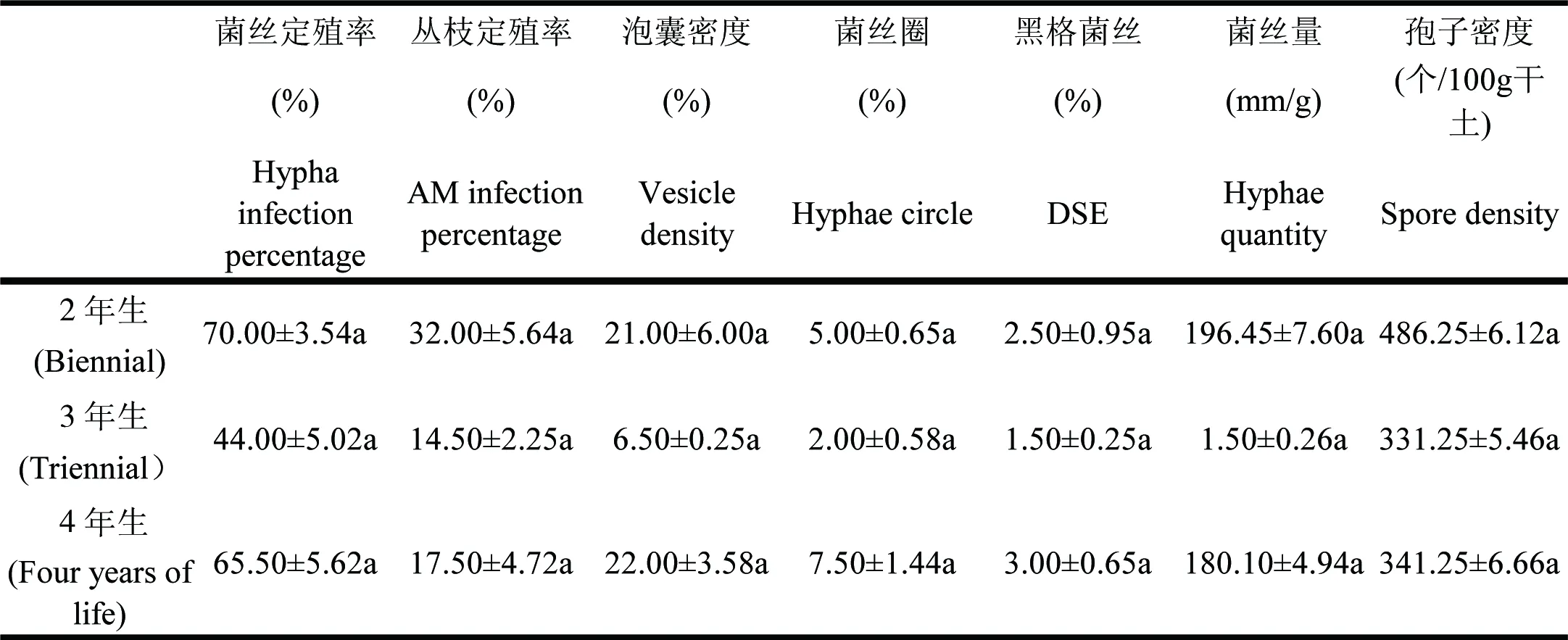
数值为平均值,±为标准差;表中的字母代表在不同年生时P<0.05的水平上差异是否显著
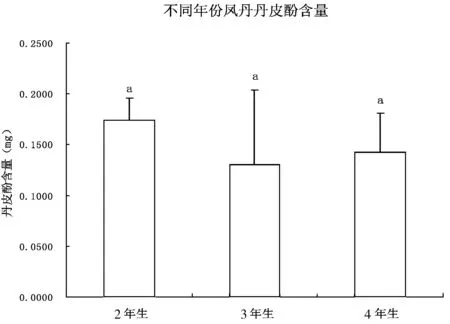
图 2 不同年生凤丹根部丹皮酚含量变化
注:同一表格中的不同字母表示在P<0.05水平上差异显著
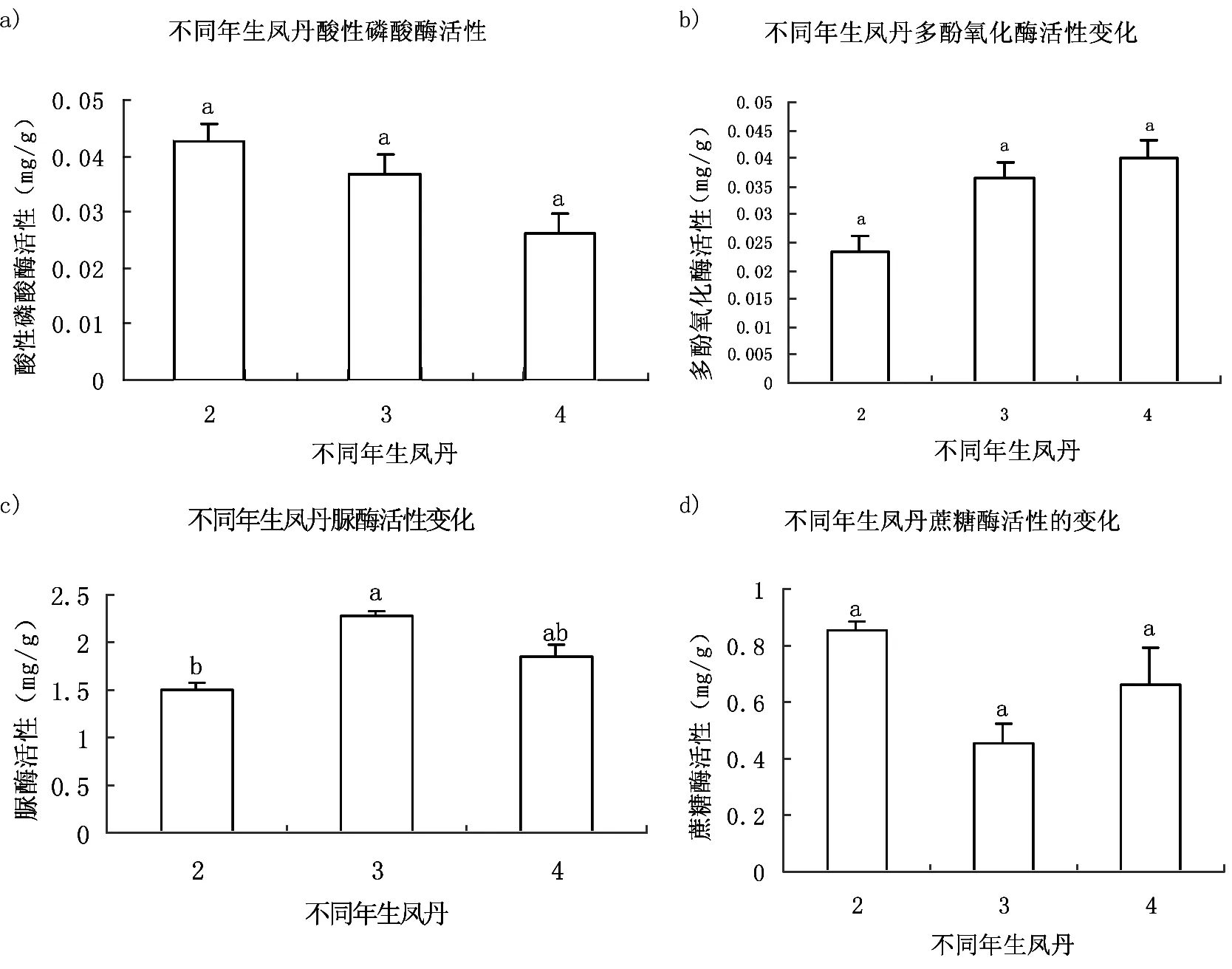
图 3 不同年生凤丹根际土壤酶的活性变化
同一表格中的不同字母表示在P<0.05水平上差异显著
2.8 凤丹根际AM真菌的参数与土壤酶活性的冗余分析
为进一步验证凤丹根际AM真菌的参数与土壤酶活性之间的关系,把AM真菌的参数与土壤酶活性进行了冗余分析(图4)。该图由两部分组成一部分是黑色的向量,为AM真菌及黑隔菌丝的参数(HC,SD,H,V,A,HQ,DSE),另一部分是红色的向量为土壤酶活参数(PPO,PCA,S,U)。向量越长代表各参数越重要,AM真菌的参数与土壤酶活的夹角的余弦值代表他们之间的相关系数,两者之间的向量同方向代表有较强的正相关反方向代表有较强的负相关,垂直表明不相关。由图4可以看出,酸性磷酸酶、脲酶、蔗糖酶、多酚氧化酶与土壤菌丝量呈正相关,酸性磷酸酶、脲酶、蔗糖酶、多酚氧化酶与泡囊密度呈负相关,孢子密度和菌丝圈与酸性磷酸酶、蔗糖酶、多酚氧化酶呈正相关,脲酶呈负相关;菌丝与AM真菌与酸性磷酸酶呈正相关,与脲酶、蔗糖酶、多酚氧化酶呈负相关。
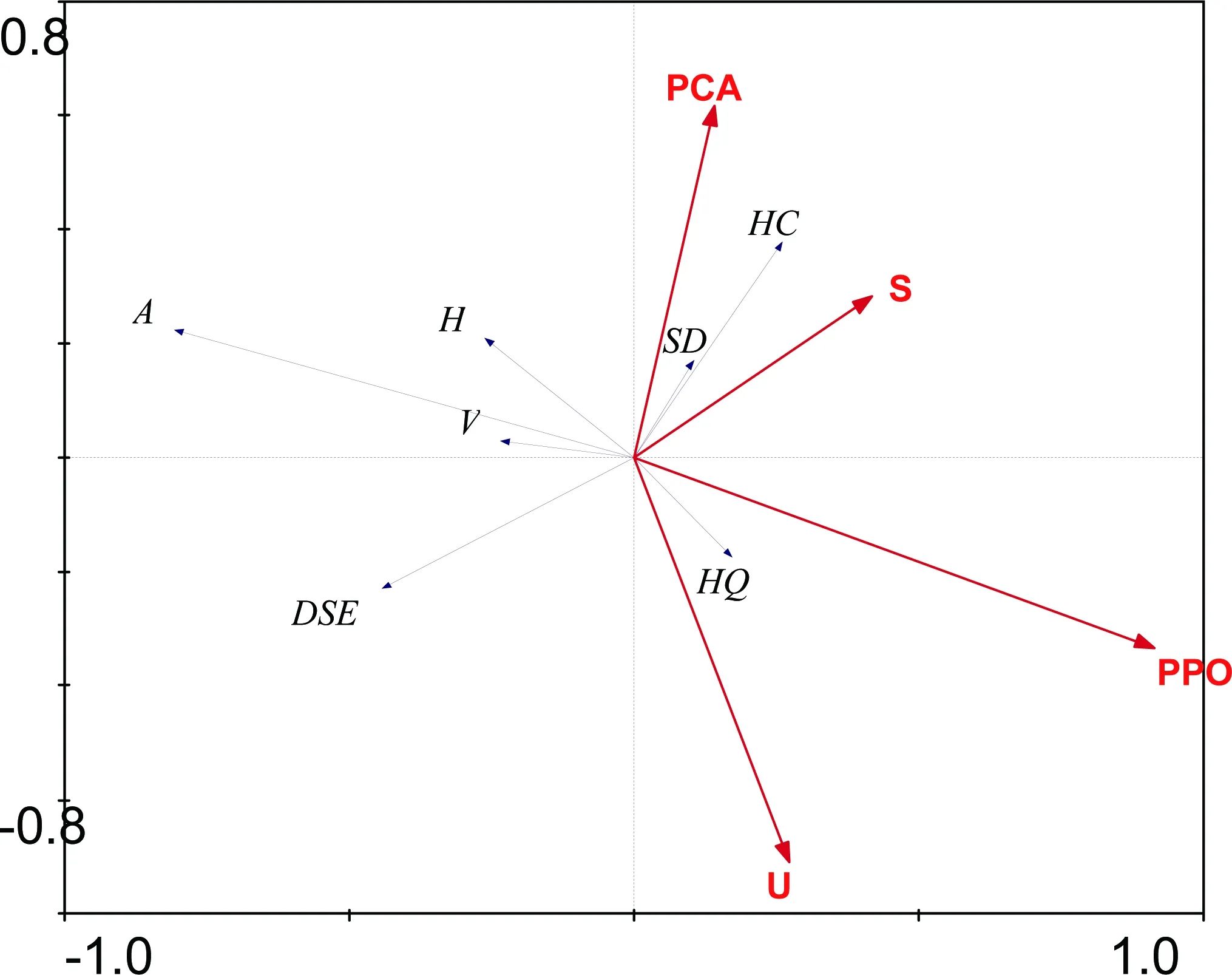
图 4 凤丹AM真菌参数与土壤酶活性的冗余分析
图中字母HQ为菌丝量;HC为菌丝圈;SD为孢子密度;V为泡囊密度;H为菌丝定殖率;A为丛枝定殖率;DSE为黑隔菌丝;PPO为多酚氧化酶;PCA为酸性磷酸酶;S为蔗糖酶;U为脲酶
3 讨论
研究发现凤丹与AM真菌的共生现象十分普遍,所采取样地的全部根样都被AMF不同程度的侵染,说明凤丹能与AM真菌形成良好的共生关系。凤丹与AM真菌都能形成良好的共生关系可能与其是多年生木本及其遗传背景复杂的生物学特性相关。根内形成典型的Arum-type类型的AM结构,这与贺学礼等[31]研究的沙棘根围与AM真菌共生形成的AM结构类型相同。在不同年生的凤丹中, AM真菌侵染率、菌丝侵染率与孢子密度均表现为2年生>4年生>3年生,说明三者在不同年生间的变化趋势相同,它们在生态系统中是相互促进的。由于丛枝定殖率、孢子密度和菌丝定殖率在不同年生间无显著差异性,说明它们在不同年生时侵染能力较稳定。
实验结果表明土壤中菌丝量为180~200 mm/g(表1),大量的根外菌丝对促进植物养分和水分的吸收有重要作用。凤丹通过形成良好的菌根定殖率和大量的根外菌丝对土壤贫瘠地区凤丹的种植具有重要的意义。
凤丹根部丹皮酚含量随不同年生产生的变化趋势与根际AM真菌在不同年生的侵染趋势相同,说明当丛枝定殖率增高时,丹皮酚的平均含量也会增加,可能是当侵染能力增强时,会提高宿主植物对土壤中矿质元素及水分的吸收,使凤丹皮中活性成分之一的丹皮酚平均含量增加。
黑格菌丝作为一种内生真菌并不属于AM真菌,但是在凤丹根部却同时看到了这两类真菌,说明这两类真菌可以同时生存,但是二者之间的关系还有待研究。
土壤酶与AM真菌结构有着密切关系。酸性磷酸酶、脲酶、蔗糖酶、多酚氧化酶的活性与土壤菌丝含量呈正相关,说明这4种酶对土壤中微生物的活动有着促进的作用;酸性磷酸酶、脲酶、蔗糖酶、多酚氧化酶的活性与泡囊定殖率呈负相关,可能是由于这4种酶参与土壤微生物与植物间的营养循环而使AM真菌没有形成贮藏营养的泡囊;菌丝定殖率、AM真菌定殖率与酸性磷酸酶的活性呈正相关,与脲酶、蔗糖酶、多酚氧化酶的活性呈负相关,可能是P对AM真菌的侵染有促进作用,N、糖类与酚类物质对AM真菌的侵染有抑制作用,还有待进一步研究。
[1]韩雪源, 张延龙, 牛立新, 等. 不同产地‘凤丹’牡丹籽油主要脂肪酸成分分析[J].食品科学, 2014, 35(22): 181-184.
[2]韩宏毅, 王 剑. 多不饱和脂肪酸及其生理功能[J].中国临床研究, 2010, 23(6): 523-525.
[3]刘立新. ω-3脂肪酸对高血压合并颈动脉粥样硬化患者血管内皮功能的影响[J].中国临床药理学杂志, 2010, 26(5): 330-333.
[4]杨振晶. 植物生长物质对‘凤丹’牡丹生根率及幼苗生长的影响[D].聊城:聊城大学, 2016.
[5]吴国成. 牡丹籽油的提取工艺及主要脂肪酸组成[J] . 中国化工贸易, 2015,7(33): 388.
[6]潘法柱. 南陵凤丹生物产业基地升为“国家级”[J].安徽林业科技, 2014(2):8.
[7]刘润进, 陈应龙. 菌根学[M]. 北京: 科学出版社, 2007: 1-447.
[8]白 祯. 基质和AM真菌对酸枣幼苗生长及营养的影响[D]. 重庆:西南大学,2013.
[9]乔亚君. AM真菌与施硒量对丹参幼苗生长及品质的影响[D].保定: 河北大学,2015.
[10]SURYANARAYANAN T S, MURALI T S, VENKATESAN G. Occurrence and distrbution of fungal endphytes in tropical for ests across a rain fall gradient [J]. Can J Bot, 2002, 80(8): 818-826.
[11]于二敏, 李 聪, 贺超兴. AM真菌对大棚芦笋矿质元素吸收和营养品质的影响[C]. 乌鲁木齐:中国设施园艺学术年会,2014.
[12]贺 超, 陈伟燕, 贺学礼, 等. 不同水肥因子与AM真菌对黄芩生长和营养成分的交互效应[J] .生态学报, 2016, 36(10): 2798-2806.
[13]陈伟燕. 不同水肥条件下AM真菌对黄芩生长的影响[D].保定: 河北大学, 2014.
[14]ABDESSAMAD T A, LINGUA G, AVROVA A. A cullin gene is induced in tomato roots forming arbuscular mycorrhizae [J]. Can J Bot, 2002, 80(6): 607-616.
[15]鲍长余. 三叉苦化学成分和其他两种药用植物有效成分含量测定的研究[D] . 海口: 海南南师范大学, 2012.
[16]仇惠君, 曾富兰, 曾燕红, 等. AM真菌对黄花蒿养分吸收和有效成分积累的效应[C]. 南昌:中国植物学会会员代表大会暨八十周年学术年会, 2013.
[17]黄京华, 谭钜发, 揭红科, 等. 丛枝菌根真菌对黄花蒿生长及药效成分的影响[J].应用生态学报, 2011, 22(6): 1443-1449.
[18]李建恒. 不同因素对丹参等药用植物化学成分的影响研究[D].保定: 河北大学, 2014.
[19]王 平, 贺学礼, 马 丽, 等. 3种杀真菌剂对AM真菌侵染和黄芩生长的影响[C]. 重庆:全国菌根学术研讨会, 2011.
[20]贺学礼, 王凌云, 马 晶, 等. 河北省安国地区丹参根围AM真菌多样性[J].生物多样性, 2010, 18(2): 175-181.
[21]秦 燕, 何 峰, 仝宗永, 等. 刈割对羊草草原土壤酶活性和养分含量的影响[J].草业学报, 2016, 25(4): 55-62.
[22]LI Y P, FENG Y L, CHEN Y J, et al. Soil microbes alleviate allelopathy of invasive plants [J]. Science Bulletin, 2015, 60(12):1083-1091.
[23]欧 洪, 郭冬琴, 林俊杰, 等. AM真菌对滇重楼根际土壤微生物数量及酶活性的影响[J].中药材, 2016, 39(5): 948-955.
[24]许庆龙, 刘晓敏, 徐小兵, 等. 4种丛枝菌根真菌对南高丛蓝莓抗旱性的影响[J]. 浙江大学学报(农业与生命科学版), 2016, 42(4): 427-434.
[25]BERCH S M, KENDRICK B. Vesicular-arbuscular mycorrhizae of southern ontario ferns and fern-allies [J]. Mycologia, 1982, 74: 769-776.
[26]MCGONIGLE T P, MILLER M H, EVANS D G. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi [J]. New Phytologist, 1990, 115(3): 495-501.
[27]KOSKE R E, WALKER C. Gigaspora erytyropa, a new species forming arbuscular mycorrhizae[J]. Mycologia, 1984, 76(2): 250-255.
[28]关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986: 45-47.
[29]杨安娜, 李凌飞, 赵之伟. 滑桃木的丛枝菌根[J].云南大学学报(自然科学版), 2004, 26(1): 85-87.
[30]JUMPONEN A. Dark septate endophytes-Are they mycorrhizal [J].Mycorrhiza,2001, 11(4): 207-211.
[31]贺学礼, 陈 程, 何 博. 北方两省农牧交错带沙棘根围 AM 真菌与球囊霉素空间分布[J].生态学报, 2011, 31(6): 1653-1661.
