大泷六线鱼6个野生群体遗传多样性的12S rRNA基因分析
2017-11-17王伟高伟峰张赛赛董安然姜欣彤
王伟 ,高伟峰 ,张赛赛 ,2,董安然 ,姜欣彤
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁 大连 116023;2.天津现代晨辉科技集团有限公司,天津 301800)
大泷六线鱼6个野生群体遗传多样性的12S rRNA基因分析
王伟1,高伟峰1,张赛赛1,2,董安然1,姜欣彤1
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁 大连 116023;2.天津现代晨辉科技集团有限公司,天津 301800)
本文利用PCR技术扩增了6个野生群体大泷六线鱼线粒体12SrRNA基因的部分序列。在获得的582bp碱基序列中,共检测到314个变异位点,占总位点数的53.95%。34尾个体共定义了16种单倍型,6个群体的单倍型多样性指数为0.40~1.00,核苷酸多样性指数为0.00071~0.10441。中性检验Fu's Fs值为1.19782(P<0.01)。分子变异分析表明:Fst=0.88326(P<0.01),群体间和群体内变异分别为88.33%和11.67%。不同个体间的遗传距离构建的NJ和MP系统树表明,东港和旅顺群体亲缘关系较近,构成一个分支,其他群体构成另一分支。
大泷六线鱼;野生群体;线粒体DNA;12SrRNA部分序列
大泷六线鱼Hexagrammos otakii Jordan&Starks也被称为欧氏六线鱼、六线鱼等[1,2],属鲉形目Scorpaeniformes、六线鱼科Hexagrammidae、六线鱼属Hexagrammos[2],俗称黄鱼,广泛分布于我国东部沿海及日本、韩国、朝鲜等国的近海[3,4],在我国主要分布于辽宁和山东等地近海的多岩礁海区。大泷六线鱼味道鲜美,营养丰富,深受人们的青睐,还可用于出口创汇,具有极高的经济价值,在我国北方的海水养殖经济鱼类中占有重要地位。早在20世纪70年代,日本便对其开展人工繁殖研究,我国也从20世纪80年代开始大泷六线鱼的人工育苗[5-8]、营养成分分析[9,10]、生物学指标[11,12]等研究。然而关于大泷六线鱼的遗传多样性的研究较少,目前国内仅见刘奇[13]的遗传多样性研究。
鱼类种群的种质资源评估对渔业资源的合理开发和利用具有重要的导向作用。在鱼类种质资源的相关研究中,检测线粒体DNA(mtDNA)多态性是一种较为普遍方式。线粒体DNA的核苷酸分子较小,易与核DNA分离和纯化,进化速率较快,是其他单拷贝DNA的5~10倍。同一合子来源的个体各组织器官内的mtDNA结构均一,每个鱼类个体中所含的mtDNA具有高度的一致性[14]。遗传中遵循严格的母系遗传规律,通常不会发生重组,容易确定其种质特征,反映群体遗传特征、种群分化和种属关系等特点。因此,作为一种较为常用的遗传分子标记,广泛应用于系统进化和种群遗传研究中[15,16]。12SrRNA基因位于线粒体基因组,具有较高的保守进化性,多用于系统发生及分子进化等研究[17,18]。但该基因较少应用于研究鱼类群体遗传多样性等。本试验分析6个群体大泷六线鱼线粒体DNA中12S rRNA序列的遗传结构、遗传多样性和遗传分化等,有助于了解不同大泷六线鱼群体的遗传多样性及其遗传背景,为合理利用渔业资源以及资源的保护和可持续开发,提供一定的理论基础。
1 材料与方法
1.1 样本采集
试验用鱼分别采自青岛、丹东和大连等6个地区,为保证采样群体的准确性,样品均采自各地的采样点码头,样品规格为(20±1.5)cm。样本采集相关信息详见图1和表1。
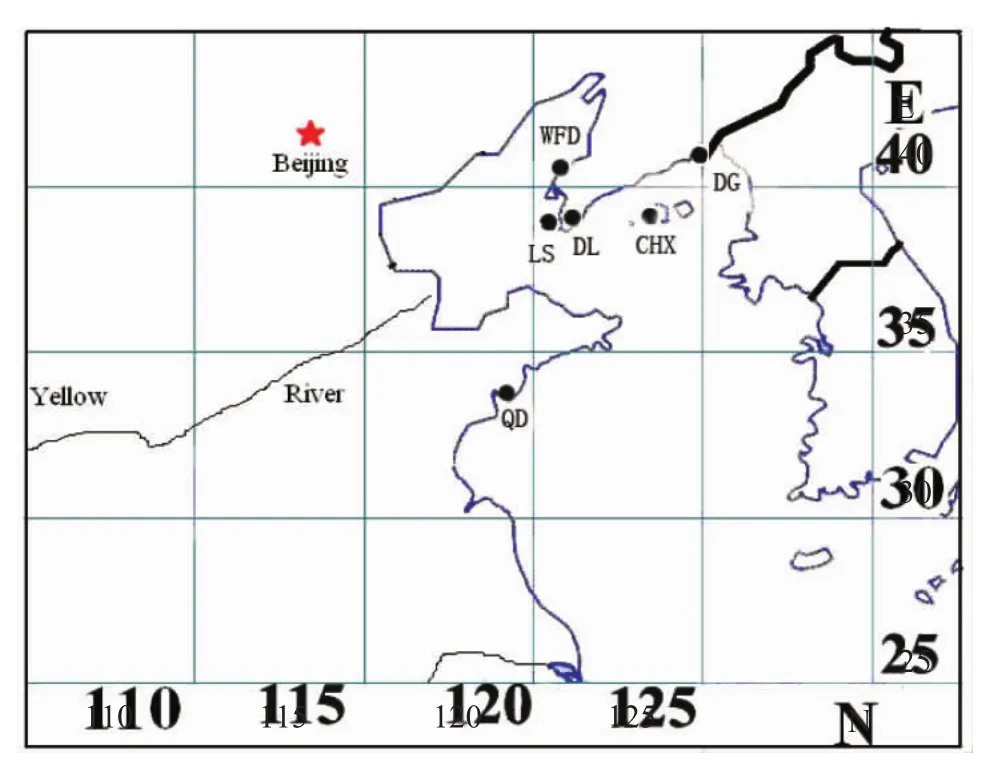
图1 大泷六线鱼采集地点Fig.1 Sampling sites of fat greenling Hexagrammos otakii

表1 大泷六线鱼样品信息Tab.1 Sample information of fat greenling Hexagrammos otakii
1.2 基因组DNA提取
DNA提取采用本实验室常用的一种简化方法进行。从鱼尾部取约50mg肌肉组织样品,充分剪碎,加入蛋白酶K、匀浆液以及SDS,经过大约2h的完全消化,使用C2H3KO2溶液进行抽提,加入无水乙醇形成沉淀,用75%酒精洗涤沉淀物,再用适量TE溶解,于-20℃下保存备用[19]。
1.3 12S rRNA序列的PCR扩增
从NCBI(National Center for Biotechnology Information)的GenBank中检索大泷六线鱼12S rRNA基因同源序列,运用Primer Primer 5软件设计PCR扩增引物、上游引物(5’-CAGAGCGTGGCTAAGATGGA-3’) 和 下 游 引 物(5’-CAGAGCGTGGCTAAGATGGA-3’)。使用约50ng基因组DNA作为PCR反应的模板。反应体系中包含:1.0mmol/LMgCl2、2μL基 因 组 DNA、0.2mmol/L dNTPs、2.5μL 10 ×PCR Buffer、引物各 0.4μmol/L、1U Taq 酶,PCR 反应的总体积为25μL。将扩增体系于95℃下3min预变性,然 后 94℃ 变 性 30s,56℃ 退 火 45s,72℃ 延 伸1min,35个循环结束后,72℃延伸10min。扩增产物采用琼脂糖凝胶试剂盒进行回收纯化。PCR产物委托上海鼎安生物科技有限公司采用双向测序法进行测序,依据峰图对测序的结果进行确认。
1.4 数据分析
采用ClustalX1.83软件对所得的序列进行对位排列,并结合人工校正;采用Mega4.10软件核对碱基组成及相关的变异位点,计算遗传距离,构建NJ和MP分子系统树[20];采用DNASP4.10软件计算核苷酸多样性指数(π)、单倍型多样性指数(Hd)及单倍型数(h);采用Arlequin3.11软件进行中性检验,计算Fu's Fs值推断大泷六线鱼各个群体在历史上发生种群扩张的情况[21];最后结合分子变异分析方法(AMOVA)分析种群间的遗传分化指数Fst,通过排列测验法(permutation test)检测Fst的显著性。
2 结果与分析
2.1 序列特征
PCR扩增所得到的碱基序列长度约为700bp,比对得出34尾大泷六线鱼mtRNA中高度一致的12SrRNA序列,长度约为 582bp。A、T、G、C碱基的平均含量各为30.0%、27.0%、21.0%和22.0%。A+T含量(59.0%)高于G+C含量(41.0%)。旅顺群体T+A含量最高(57.4%),长海县群体最低(56.5%)。G+C含量则相反,长海县群体最高(43.5%),旅顺群体最低(42.6%)。
在34尾大泷六线鱼12S rRNA序列中,共检测到314个变异位点,多态位点百分率为53.95%。其中单态位点53个,简约信息位点261个,插入/缺失位点32个。共有转换位点54个,其中C-T有28个位点,A-G有26个位点。颠换位点62个,四种形式均有,其中A-C有19个位点,T-G有12个位点,A-T有17个位点,G-C有14个位点。平均转换与颠换之比为0.87。由表2可知,6个群体的34尾大泷六线鱼共定义16种单倍型。2个单倍型为不同群体所共享,仅占单倍型总数的12.5%,14种单倍型均为单个群体所特有,在单倍型总数中占87.5%,仅有1个为4个种群的共有单倍型,1个为2个种群的共有单倍型。
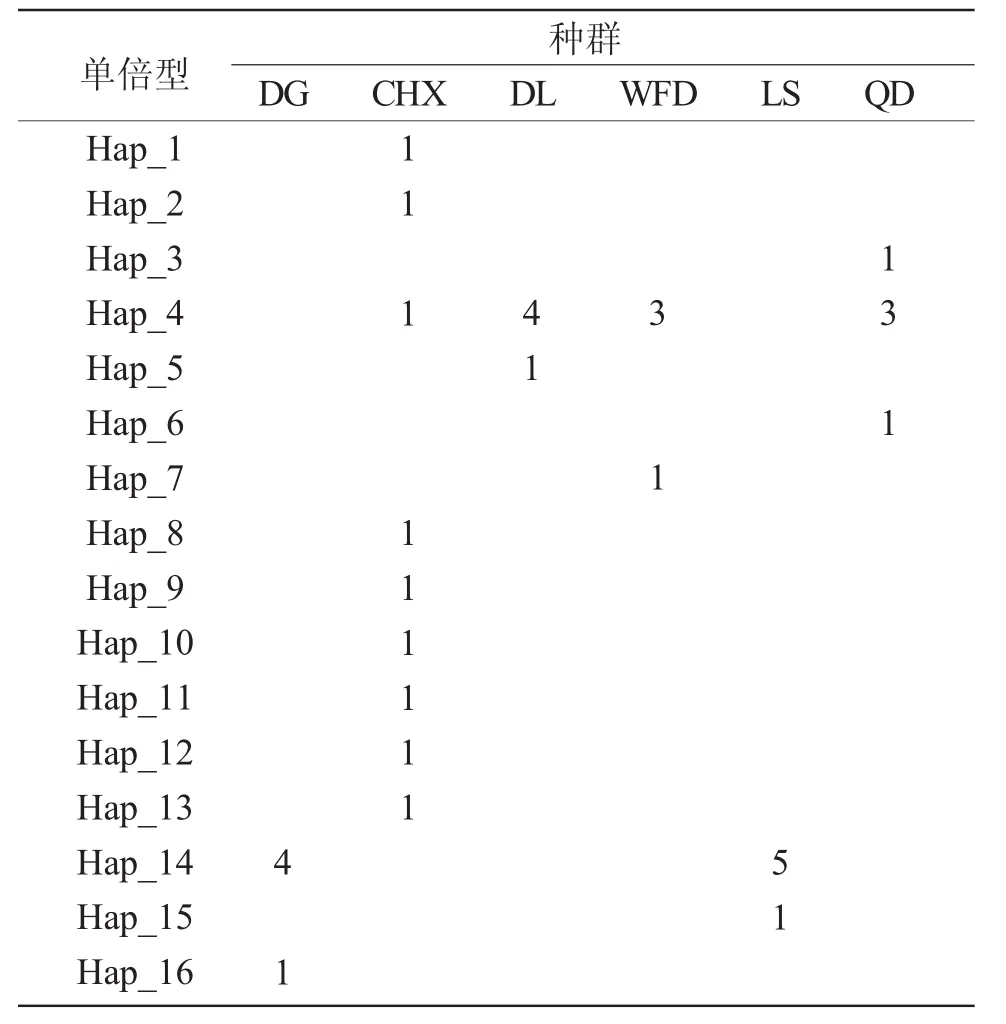
表2 16种单倍型在大泷六线鱼群体中的分布Tab.2 The distribution of the 16 haplotypes in fat greenling Hexagrammos otakii populations
2.2 遗传多样性
群体遗传多样性参数的统计结果(表3):表明各种群遗传多样性差异明显。长海县群体的核苷酸多样性指数(0.10441)和单倍型多样性指数(1.00)最高,而东港和大连群体的单倍型多样性指数最低(0.40),大连群体的核苷酸多样性指数最低(0.00071)。旅顺群体序列Fu's Fs值的计算结果为负值,统计学计算的结果具有显著的差异性,可将此群体的检验结果定义为偏离中性模式。
2.3 遗传结构
Kimura 2-paramter遗传分化指数Fst和遗传距离的计算结果(表4)表明,6个群体间的平均遗传距离为0.00097~0.76765,其中瓦房店群体与东港群体的群体间遗传距离最高,瓦房店群体与大连群体的群体间遗传距离最低。6个群体间的遗传分化指数Fst为0~0.99832,其中大连群体与旅顺群体之间的遗传分化指数最高,东港与旅顺群体之间以及青岛、瓦房店和大连群体之间的分化可以忽略不计。
由表5可知:分子变异分析(AMOVA)的结果为Fst=0.88326(P<0.01)。群体间变异占变异总量的88.33%,群体内变异仅占11.67%,由此推断,6个大泷六线鱼群体间发生了显著的遗传分化。
2.4 分子系统树的构建
外群选用斑头鱼Agrammus agrammus的同源序列(登录号:AB290791.1)进行比对分析,基于Kimura-双参数计算群体间的遗传距离,构建NJ和MP分子系统进化树,通过1 000次bootstrap检验对其节点的支持率进行检验。由图2可知,NJ和MP分子系统进化树具有极高的相似度,6个群体的34尾大泷六线鱼大致呈现出两个分支。其中旅顺和东港群体的亲缘关系较为接近,为一条分支,其他群体皆位于另一条分支。
3 讨论
遗传多样性是物种延续和发展的基础[22],其丰富的种群在复杂多变的自然环境中表现出更强的适应性。近年来,随着渔业资源的开发利用力度加大,鱼类种群遗传多样性减少,对各个鱼类种群的延续能力产生不良影响,严重时甚至会导致濒危物种的灭绝速度加快[23]。在本研究中,共测序得出16种单倍型,各个群体的单倍型多样性指数范围为0.40~1.00,核苷酸多样性指数范围为0.00089~0.10441,各群体具有较为显著的遗传多样性差异。本研究中,旅顺群体的变异位点数最高,达到244个,核苷酸多样性指数(0.10441)最高,说明旅顺群体的遗传多样性非常丰富。分析旅顺的地理位置及其结果发现,旅顺东临黄海,西濒渤海,南与山东半岛隔海相望,中间有水道可与外海相通,对基因的交流提供了极大的便利。而大连群体(0.40,0.00071)与东港群体(0.40,0.00089)的遗传多样性相对贫乏,其原因可能与其所处的地理环境有关。Fu[24]认为,Fs中性检验的结果显著偏离中性,P值具有显著差异且Fs值呈负值,表明该种群在其进化历程中,可能发生过快速的种群扩张。在本试验的1 000次的模拟检测中,旅顺群体的Fs值为负值(P<0.05),推断旅顺的大泷六线鱼群体可能在历史上发生过短时间、大规模的种群扩张,使该地区大泷六线鱼群体的遗传多样性较高。

表3 6个大泷六线鱼群体单倍型参数及遗传多样性比较Tab.3 The comparison of haplotype diversity and genetic diversity among the 6 populations of fat greening Hexagrammos otakii

表4 各种群之间的遗传距离(左下角)及其遗传分化指数Fst(右上角)Tab.4 Genetic distance(below diagonal)and Fst(above diagonal)among different populations of fat greening Hexagrammos otakii

表5 大泷六线鱼群体的AMOVA分析Tab.5 The AMOVA analysis of the populations of fat greening Hexagrammos otakii

图2 基于DNA 12S rRNA的16个单倍型构建的NJ和MP分子系统树Fig.2 Neighbor-joining and Maximum Parsimony tree of fat greening Hexagrammos otakii based on the 16 haplotypes in mtDNA 12S rRNA sequences
群体间的遗传距离以及遗传分化指数是衡量群体分化程度的重要指标,二者的数值与群体分化程度为线性关系。东港群体以及旅顺群体与其他群体间Fst值都大于0.8,接近1;长海县群体与青岛、瓦房店和大连群体之间的值也大于0.3,其他三个群体间的遗传多样性分化程度可忽略不计。由于各种群间的亲缘关系与其相对遗传距离呈明显的正相关[25],因此,青岛及旅顺群体与其他群体之间的遗传距离(0.68805~0.76765)明显高于旅顺与东港群体之间的遗传距离(0.00209)及其他三个群体之间的遗传距离(0.00097~0.09305),表明青岛及旅顺群体的亲缘关系较为接近。AMOVA分析结果表明,群体间变异占变异总量的88.33%,6个样点的大泷六线鱼群体间差异遗传较大,这与程起群等(2007)[26]、李莹(2012)等[19]研究结果一致。NJ和MP分子系统树的结果与遗传分化指数的结果一致,这与张源真等研究结果一致[27]。东港群体和旅顺群体为一个分支,长海县、瓦房店、大连和青岛群体共同组成另一大分支。mtDNA属母系遗传,由此可推断,这6个大泷六线鱼群体在进化过程中,曾由共同祖先分化为两个进化分支,继而分别展开进一步的分化。
本研究基于线粒体控制区的对比发现,大泷六线鱼的分布与其基因多样性的地理位置分布存在一定差异。青岛群体与大连群体间的亲缘关系反而较为接近,这可能基因交流所形成,青岛群体形成的时间较短,分化速度较快,而12S rRNA序列比较保守,在短时间内无法形成足够的有效信息含量[28]。同时也可能是进化树的构建过程中,受多方面因素影响的结果,例如分析软件的不同,所选择的分子标记、物种差异以及计算方法的不同等,目前由于上述因素缺乏统一的标准,都有可能影响分析结果。但是通过本试验,可以发现除东港群体与旅顺群体外,其他群体间依然具有较大的遗传分化,这种分化的结果与其地理位置的分布具有较高的一致性。因此结合本次针对两种分子标记方法的研究结果,建议按照不同的地理区域对大泷六线鱼种群资源展开划区域、多层次的保护,使渔业资源能够得到更加合理的开发,达到可持续利用的目标。
[1]刘蝉馨,秦克静.辽宁动物志(鱼类)[M].沈阳:辽宁科学技术出版社,1987:393-396.
[2]成庆泰,郑宝珊.中国鱼类系统检索[M].北京:科学出版社,1987.
[3]辽宁省海洋局.辽宁省海岛资源综合调查研究报告[M].北京:海洋出版社,1996:357.
[4]唐启升,叶懋中.山东近海渔业资源开发与保护[M].北京:农业出版社,1990:80-102.
[5]Fukuhara O and Fushimi T.Development and early live history of the greenlings Hexagrammos otakii(spices:Hexagrammidae)reared in the laboratory[J].Bulletin of the Japanese SocietyofScientific Fisheries,1983,49(12):1843-1848.
[6]吴立新,秦克静,姜志强,等.大泷六线鱼(Hexagrammos otakii)人工育苗初步试验[J].海洋科学,1996(4):32-34.
[7]于鸿仙,庄虔增,徐春华,等.六线鱼人工育苗技术研究[J].齐鲁渔业,1998,15(5):21-24.
[8]庄虔增,于鸿仙,刘岗,等.六线鱼育种生产技术的研究[J].中国水产科学,1999,6(1):103-106.
[9]康斌,武云飞.大泷六线鱼的营养成分分析[J].海洋科学,1999(6):23-25.
[10]Lee S M,Cho S H and Kim K D.Effect of dietary protein and energy levels on growth and body composition of juvenileflounderParalichthys olivaceus[J].WorldAquaSoc,2000,31(3):306-315.
[11]王书磊,姜志强,苗治欧.大连海区大泷六线鱼生物学指标的季节变化[J].水产科学,2005,24(5):1-3.
[12]刘奇,王亮,高天翔,等.北黄海大泷六线鱼主要生物学特征比较研究[J].中国海洋大学学报:自然科学版,2009,39(S1):13-18.
[13]刘奇.大泷六线鱼(Hexagrammos otakii)生物学特征与遗传多样性研究[D].青岛:中国海洋大学,2010.
[14]郑光明,朱新平,张跃.鱼类种质鉴定技术与渔业管理[J].中国水产科学,1996,6(2):108-111.
[15]Lu G and Li S.Advances in the study and application of fishmitochondrialDNApolymorphism[J].JournalofFisherySciences ofChina,1998,3:94-103.
[16]李春枝,张邦杰,李本旺,等.尖塘鳢属鱼类线粒体12S rRNA 基因序列分析[J].生态科学,2006,25(5):433-436.
[17]Honda M,Otah K,Kobayashi M,et al.Phylogenetic relationships ofAustralian skinks ofthe Mabuya group(Reptilia:Scincidae)inferred from mitochondrial DNA sequences[J].Genes Genet Syst,1999,74(4):135-139.
[18]Medina Mand Walsh P J.Molecular systematics ofthe order Anaspida based on mitochondrial DNA sequences(12S,16S,and COI)[J].Molecular Phylogenetics and Evolution,2000,15(1):41-58.
[19]李莹,王伟,孟凡平,等.利用线粒体DNA控制区部分序列分析不同地理群体大泷六线鱼遗传多样性[J].海洋科学,2012,36(8):40-46.
[20]Kumar S,Tamura K and Nei M.MEGA3:integrated software for molecular evolutionary genetics analysis and sequence alignment[J].Briefings in Bioinformatics,2004(5):150-163.
[21]Rozas J,Sánchez-DelBarrio J C,Messeguer X,et al.DnaSPDNA polymorphismanalyses bythe coalescent and other methods[J].Bioinformatics,2003,19(18):2496-2497.
[22]郭新红,刘少军,颜金鹏,等.异源四倍体鲫鲤、三倍体湘云鲫和它们父母本线粒体DNA 12S rRNA基因遗传变异的分析[J].遗传,2004,26(6):875-880.
[23]季维智,宿兵.遗传多样性研究的原理与方法[M].杭州:浙江科学技术出版社,1999.
[24]Fu Y X.Statistical tests of neutrality of mutations against population growth,hitchhiking and background selection[J].Genetics,1997,147(2):915-925.
[25]任岗,章群.中国沙塘鳢属鱼类线粒体12S rRNA基因序列分析[J].水生生物学报,2007,31(4):473-478.
[26]程起群,马春艳,缪炯,等.基于线粒体12S rRNA序列研究凤鲚两个野生群体的遗传多样性[J].大连水产学院学报,2007,22(5):387-391.
[27]张源真,王伟,姜志强,等.基于16S rRNA部分序列探讨部分鳚亚目鱼类的分子系统进化关系[J].海洋科学,2012,36(9):89-95.
[28]邵爱华,朱江,史全良,等.暗纹东方鲀线粒体12S rRNA基因的克隆及序列分析[J].苏州科技学院学报:自然科学版,2008,25(4):39-44.
Genetic Diversity Analysis of Six Wild Populations of Fat Greenling Hexagrammos otakii by Mitochondrial DNA 12S rRNA Partial Sequence
WANG Wei1,GAO Wei-feng1,ZHANG Sai-sai1,2,DONG An-ran1,JIANG Xin-tong1
(1.Key Laboratory of Mariculture&Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China;2.Tianjin Chen Hui Modern Technology Group Co.,Ltd.,Tianjin 301800,China)
Nucleotide sequences of mitochondrial DNA 12S rRNA partial sequence were amplified in six wild populations of fat greenling Hexagrammos otakii using PCR techniques.A total of 314 variable sites were detected among the aligned sequences of 582 bp with mutation rate of 53.95%.Sixteen haplotypes were identified from 34 individuals according to the determined sequences,and the six populations had haplotype diversity of 0.40~1.00,nucleotide diversity of 0.00071~0.10441 and Fu's Fs of 1.19782 in neutrality tests(P<0.01).AMOVA analysis demonstrated that:there were Fst=0.88326(P<0.01),variance of 88.33%among six populations and variance of 11.67%within the six populations.The NJ and MP molecular phylogenetic trees constructed by the distances among different individuals revealed that the phylogenetic trees were all divided into two branches,one in Donggang and Lvshun populations and the others in the other populations.
Hexagrammos otakii;wild population;mitochondrial DNA;12S rRNA partial sequence
S917
A
1005-3832(2017)05-0007-06
2017-04-18
大连市科技计划项目(2015B11NC072);大连市科技之星计划项目(2015R081).
王伟(1978-),男,理学博士,研究方向为鱼类群体遗传学.E-mail:wangwei@dlou.edu.cn
