龙眼漆酶基因(DlLac)的克隆及表达分析
2017-07-31赵昱清廖玲燕宋慕波朱东建段振华吴振先韩冬梅
帅 良,赵昱清,+,廖玲燕,宋慕波,朱东建,蔡 文,段振华,吴振先,韩冬梅
(1.贺州学院食品科学与工程技术研究院,广西贺州 542800;2.华南农业大学园艺学院,广东省果蔬保鲜重点实验室,广东广州 510642;3.广东省农业科学院果树研究所,农业部南亚热带果树生物学与遗传资源利用重点实验室,广东广州 510640)
龙眼漆酶基因(DlLac)的克隆及表达分析
帅 良1,2,赵昱清1,2,+,廖玲燕1,宋慕波1,朱东建1,蔡 文1,段振华1,吴振先2,*,韩冬梅3,*
(1.贺州学院食品科学与工程技术研究院,广西贺州 542800;2.华南农业大学园艺学院,广东省果蔬保鲜重点实验室,广东广州 510642;3.广东省农业科学院果树研究所,农业部南亚热带果树生物学与遗传资源利用重点实验室,广东广州 510640)
以‘石硖’龙眼为试材,采用RT-PCR结合RACE技术成功克隆一个龙眼漆酶基因全长cDNA序列,命名为DlLac,NCBI登录号为KY051551。DlLac基因序列全长1898 bp,编码576个氨基酸,NCBI比对结果显示其与荔枝漆酶氨基酸序列同源性最高,高达94%;进化树结果显示龙眼漆酶氨基酸序列与荔枝漆酶氨基酸序列具有高度同源性。氨基酸保守序列结果表明龙眼漆酶氨基酸序列含有漆酶的3个典型保守结构域,分别为:Cu-oxidase-3、Cu-oxidase和Cu-oxidase-2。结合龙眼果实在常温、低温贮藏条件下,果皮褐变与DlLac的表达关系,推测DlLac的上调表达可能对龙眼果皮褐变起促进作用。
龙眼,漆酶,果皮褐变,基因克隆,表达
龙眼(DimocarpuslonganLour.)是我国南方重要的优势特色水果之一,主产于福建、广东、广西和台湾等省。果实成熟于盛夏高温季节,采收后果实生理代谢旺盛,极易发生果皮褐变而严重影响产品的贮运和商品价值[1-2]。研究表明,果皮酶促褐变是引起龙眼果实褐变的最主要原因,其中酚类物质氧化酶在酶促褐变中起着至关重要的作用[3]。酚类氧化酶大体上可以分成儿茶酚氧化酶和漆酶两大类,儿茶酚氧化酶又称为多酚氧化酶。它与漆酶的主要区别在于,儿茶酚氧化酶可以将多种酚类物质氧化成醌类,而漆酶仅能将对苯二酚氧化成对苯醌[4]。
漆酶(laccase,LAC),最初是从日本漆树的汁液中分离得到,因而得名漆酶,是一种含铜的多酚氧化酶,属于多铜氧化酶家族和铜蓝氧化酶蛋白家族中的一员,广泛存在于真菌、细菌、昆虫和植物中[5-6]。研究人员已经在植物、真菌、昆虫和细菌中都发现了漆酶[7-10]。目前,研究最多的是真菌漆酶,其在真菌的形态发生、病原体-宿主互作、逆境防御及木质素降解等过程中起着重要作用[11]。植物漆酶的研究相对较少,人们认为植物漆酶在参与细胞壁的形成、酚类物质的氧化、木质素化和去木质素化等过程起着重要作用[12]。刘保华等在荔枝果皮上的研究发现,荔枝漆酶基因在果实采后贮藏过程中前期呈上调表达,推测其可能起促进褐变的作用[7]。Fang等通过对荔枝的研究发现,荔枝漆酶在果皮褐变中起着降解花色素苷的作用,由此推知,漆酶在荔枝果皮褐变中起促进作用[13]。而有关龙眼漆酶与龙眼果皮褐变之间的关系至今未见报道。本研究通过设计兼并引物,以龙眼果皮为材料,克隆龙眼漆酶基因,并对其序列特征进行分析,探讨龙眼果皮褐变与漆酶基因表达之间的关系,为进一步揭示龙眼果皮褐变奠定基础。
1 材料与方法
1.1 材料与仪器
‘石硖’龙眼 广东省农业科学院果树研究所种质资源圃。采收后,选择成熟度均匀一致,无病、虫、伤、褐的龙眼果实于施保克溶液中浸泡3 min,取出晾干。其中常温(20±1) ℃贮藏用塑料小托盘包装封膜,每盘装20个果,每次取3盘作为三个重复,每2 d取样一次;低温贮藏(4±1) ℃ 用0.03 mm聚乙烯袋包装贮藏在冷库,每袋装25个果,每次取3袋作为三个重复,每8 d取样一次。取样时,将果皮和果肉分开,液氮冷冻,锡箔纸包好,置于-80 ℃超低温冰箱中保存。
Amp、X-gal、IPTG、氯化钠、氯化钙、四硼酸钠、硼酸、磷酸钠、LB培养基等试剂 上海生物工程有限公司;RNAOUT试剂盒 北京华越洋生物科技有限公司;琼脂糖、大肠杆菌感受态细胞、TaKaRa PCR Amplification kit、PMD 20-T Vector kit、TaKaRa 3′-Full RACE Core Set Ver.2.0、TaKaRa Gel DNA Purification Kit Ver2.0 TaKaRa公司;M-MLV逆转录酶 美国Life公司;SMARTTM RACE cDNA Amplification Kit试剂盒 Clontech公司;荧光定量试剂盒 德国Roche公司。
RS232C型分光光度计、5810D型台式高速离心机 德国Eppendorf公司;BS224S型电子分析天平 德国Sartorious公司;Milli-Q-B超存水系统 美国Millipoe公司;PTC-200型PCR仪 美国MJ Research公司;罗氏LightCycler® 480荧光定量PCR仪 德国Roche公司;702Rell超低温冰箱 美国Thermo Fisher公司;T2A型凝胶成像系统 美国BIO-RAD公司。
1.2 实验方法
1.2.1 内果皮褐变指数的评定 参照林河通等的方法,略有改动[14]。每次随机取20个果,根据内果皮褐变程度分为5个等级。0级果:内果皮无明显褐斑;1级果:褐变总面积小于1/4;2级果:褐变总面积达果皮总面积1/4~1/2;3级果:褐变总面积达总面积1/2~3/4;4级果:褐变总面积占内果皮总面积的3/4以上,或者长霉。重复3次。
内果皮褐变指数=∑(褐变级数×该级果数)/总果数
1.2.2 总RNA的提取和第一链cDNA的合成 本实验采用北京华越洋超快型植物RNAOUT试剂盒提取龙眼果皮的总RNA。以纯度合格且已经去除基因组DNA的RNA为模板,按TAKARA cDNA合成试剂盒操作步骤先合成cDNA第一链,反转录酶采用Reverse Transcriptase M-MLV,使用Random Primer,cDNA的合成参照说明书的方法进行,-40 ℃保存,用于漆酶基因cDNA的克隆。
1.2.3 龙眼DlLac基因片段的克隆 依据GenBank上已经报道的其它物种的中性转化酶基因序列,设计简并引物DlLac-For和DlLac-Rev(表1)。以cDNA为模板进行PCR扩增,用1%琼脂糖凝胶电泳检测扩增片段,切胶,回收,连接转化,测序。通过去载体分析所扩增的片段。
1.2.4 龙眼DlLac基因3′-RACE和5′-RACE的克隆 根据所得到的片段序列,利用Primer Premier 5.0软件,参照TaKaRa 3′-Full RACE Core Set Ver.2.0试剂盒要求设计3′-RACE引物3′-RACE-Outter-DlLac和3′-RACE-Inner-DlLac,按照Clontech SMARTTMRACE cDNA Amplification Kit试剂盒要求设计5′-RACE引物5′-Race-DlLac(表1),严格按照说明书进行后续PCR扩增,目的基因片段回收、连接、转化、鉴定、测序及结果分析同上。
1.2.5 龙眼DlLac基因全长的获得及ORF验证 将上述获得的3′-RACE和5′-RACE结果序列和片段序列采用ContigExpress软件进行拼接,获得DlLac基因全长cDNA序列,通过使用NCBI的Blast和SIB的翻译工具(Translate tool)(http://web.expasy.org/translate/)进行序列分析,找出基因全长序列的开放阅读框(ORF),设计ORF序列引物DlLac-ORF-For和DlLac-ORF-Rev,对ORF序列进行克隆,检测,以确定所拼接的序列为正确的基因序列(表1)。
1.2.6 龙眼DlLac基因的生物信息学分析 使用NCBI上的ORF(开放阅读框)Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)工具查找基因的ORF;使用SIB(Swiss Institute of Bioinformatics)的(Translate tool)(http://web.expasy.org/translate/)对ORF进行翻译;蛋白质分子量及理化性质分析采用ProtParam tool(http://web.expasy.org/protparam/)工具分析;使用NCBI上的Blastn和Blastx对序列进行比对分析;同源性及序列多重比较分析使用Clustal X 1.83软件并用GeneDoc软件进行美化编辑;运用MEGA 5软件中的Neighbor-Joining(邻位相连法,NJ)法构建系统进化树并进行编辑。

表1 龙眼漆酶基因克隆与表达分析中使用的引物Table 1 Primers used in cloning and expression analysis of DlLac
1.2.7 龙眼DlLac基因表达分析 使用BatchPrimer3在线引物设计工具设计龙眼DlLac基因定量引物DlLac-RT-For和DlLac-RT-Rev。RT-qPCR使用荧光染料为SYBR Green I Master(Roche),仪器为Roche Lightcycler® 480,使用384孔模块。10 μL扩增体系:cDNA模板1 μL,5 μ mol/L的上游引物和下游引物各0.5 μL,SYBR Green I Master(Roche)5 μL,ddH2O 3 μL。使用DlActin和DlEF-1a基因作为双内参进行RT-qPCR分析[15-17]。并用相对定量软件对PCR结果的熔解曲线和标准曲线进行分析、定量。相对表达量计算采用2-ΔΔCT法,由Roche Lightcycler® 480软件系统自动分析得出结果。
1.3 数据处理与作图
使用Microsoft Office 2013软件进行数据处理和分析,使用Origin 8.5作图,并使用Adobe Illustrator CS6软件进行图形编辑。
2 结果与分析
2.1 龙眼果皮褐变指数的变化
在常温贮藏和低温贮藏过程中,龙眼内果皮褐变指数随着贮藏时间的推移而逐渐升高;常温贮藏下,内果皮在前2 d褐变指数较低,贮藏2 d后褐变指数迅速升高;低温贮藏下,内果皮在前16 d中褐变指数较低,表现出较好的贮藏品质,贮藏16 d后褐变指数迅速上升(图1)。由此可知,低温可以抑制龙眼内果皮褐变,延长龙眼果实的贮藏时间。

图1 常温贮藏和低温贮藏下果皮褐变指数的变化Fig.1 Changes of longan pericarp browning index during room temperature(20±1) ℃ and low temperature(4±1) ℃ storage
2.2 龙眼DlLac基因的克隆
经测定提取的龙眼总RNA的OD260/OD280比值均在1.8~2.0之间,OD260/OD230比值均高于2.0[18],说明提取的龙眼总RNA的质量较高,纯度和完整性较好。进行反转录获得cDNA。
以龙眼cDNA为模板,使用简并引物DlLac-For和DlLac-Rev进行PCR扩增,得到一条长度为600 bp左右的片段(图2A),切胶,回收,连接,转化,送测序。NCBI比对结果发现,成功克隆了一条长619 bp的漆酶基因片段,命名为DlLac。
根据DlLac基因测序结果设计3′-RACE-Outter-DlLac和3′-RACE-Inner-DlLac引物(表1),按照TaKaRa 3′-Full RACE Core Set Ver.2.0试剂盒内的PCR条件进行巢式PCR扩增,电泳检测后均得到650 bp左右的目的片段(图2B),片段大小与预期相符合,切胶,回收,连接,转化,送测序,测序结果分析表明,所获得的3′末端序列与扩增的片段序列有相应的高度重复的片段,说明DlLac基因的3′末端序列扩增成功。
根据DlLac基因测序结果设计5′-RACE引物5′-Race-DlLac(表1),按照Clontech SMARTTMRACE cDNA Amplification Kit试剂盒内PCR条件进行扩增,电泳检测后1100 bp左右的目的片段(图2C),片段大小与预期相符合,切胶,回收,连接,转化,送测序,测序结果分析表明,所获得的5′末端序列与扩增的片段序列有相应的高度重复的片段,说明DlLac基因的5′末端序列扩增成功。
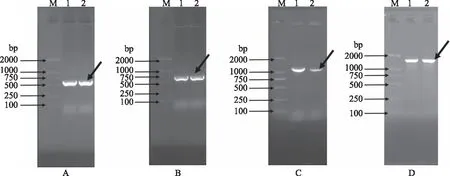
图2 龙眼DlLac基因序列扩增结果Fig.2 PCR product of DlLac gene from longan注:M:DL2000;1,2号孔道为扩增产物;A:中间片段扩增结果; B:3′端片段扩增结果;C:5′端片段扩增结果;D:ORF序列扩增结果。
根据扩增的DlLac基因片段以及3′RACE和5′RACE结果,使用ContigExpress软件对片段序列进行拼接,获得DlLac基因的全长cDNA序列,通过使用ORF Finder对序列ORF进行查找,根据ORF序列设计ORF全长引物DlLac-ORF-For和DlLac-ORF-Rev(表1),用于验证拼接全长序列的准确性,使用LA Taq酶进行PCR扩增,经电泳检测后均发现与预测片段大小相一致的条带,DlLac基因的ORF扩增产物约1800 bp左右(图2D),切胶,回收,连接,转化后送测序,测序结果去载体后经Blast比对分析发现,DlLac基因条带的ORF序列正确,结果表明成功获得了DlLac基因全长cDNA序列。

图3 龙眼漆酶氨基酸序列系统进化树Fig.3 Phylogenetic tree of laccase in different plants
2.3 龙眼DlLac基因的生物信息学分析
DlLac基因序列全长1898 bp,包括一个长度为2 bp的5′UTR和165 bp的3′UTR,ORF长度为1731 bp,编码576个氨基酸,蛋白质的分子质量为63.87 kDa,等电点为4.87,蛋白质的不稳定系数为34.87,脂溶指数为74.81,其疏水性平均值为-0.164,表明该蛋白为亲水不稳定脂溶蛋白,NCBI的登录号为KY051551,经Blast比对后发现与荔枝漆酶基因(EU527187.2)核苷酸序列同源性最高,达到了94%;氨基酸比对结果显示,与荔枝(ACB22018.2)漆酶基因推导的氨基酸序列同源性最高,达到了90%。聚类分析结果表明,龙眼漆酶氨基酸序列与荔枝漆酶氨基酸序列聚为一枝(图3)。由此可知所获得的序列确为龙眼漆酶基因序列。
通过选取荔枝(ACB22018.2)、美国梧桐(AAB09228.1)和可可(XP_007020214.1)漆酶的氨基酸序列进行保守序列分析,结果发现,龙眼漆酶氨基酸残基序列含有漆酶的3个典型保守结构域,分别为:Cu-oxidase-3、Cu-oxidase和Cu-oxidase-2(图4)。

图4 不同植物漆酶氨基酸序列比对Fig.4 Alignment of different plant laccase amino acid sequences
2.4DlLac在贮藏过程中的表达
由图5可知,在龙眼果实常温贮藏过程中,DlLac表达量随着贮藏时间的延长呈上升趋势,在贮藏的第8 d达到表达量的最大值。低温贮藏过程中,DlLac表达量随着贮藏时间的延长,呈现先下降后上升的趋势,在贮藏第32 d时表达量达到低温时的最大值。
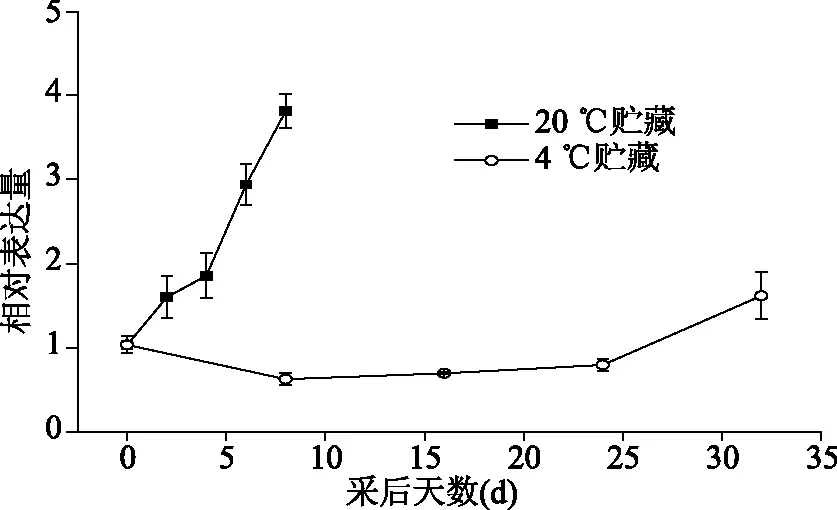
图5 DlLac在贮藏过程中的表达变化Fig.5 Changes of DlLac expression in pericarp during storage
3 结论与讨论
漆酶是一种结合多个铜离子的蛋白,是一种含铜的多酚氧化酶,它能催化许多化合物的氧化反应,底物比较广泛,包括许多与对二酚结构类似的化合物[8]。龙眼果皮中含有大量的酚类物质,且种类繁多,主要包括单宁、没食子酸、水杨酸、儿茶素、表儿茶素等[19-20]。这些酚类物质是多酚氧化酶的底物,同时也是漆酶的氧化底物。研究表明,植物漆酶能够氧化表儿茶素、单宁酸及没食子酸等多酚类物质[21]。由此可知,龙眼果皮中的漆酶能够导致龙眼果皮褐变。
本研究通过采用RT-PCR结合RACE技术成功克隆了龙眼漆酶基因全长cDNA序列,命名为DlLac,NCBI登录号为KY051551。DlLac基因序列全长1898 bp,编码576个氨基酸,比对结果发现与荔枝漆酶基因的氨基酸序列同源性最高,达到了94%。系统进化树分析发现荔枝漆酶氨基酸序列和龙眼漆酶氨基酸序列具有高度同源性。氨基酸保守序列分析发现,龙眼漆酶氨基酸序列含有漆酶的3个典型保守结构域,分别为:Cu-oxidase-3、Cu-oxidase和Cu-oxidase-2。在常温贮藏下,随着果皮中DlLac表达量上升,龙眼果皮褐变指数也呈直线上升(图1和图5),推测可能原因是随着DlLac表达上升导致龙眼果皮中漆酶总量及活性上升,使果皮中底物氧化量增加,促进龙眼果皮褐变;低温贮藏条件下,DlLac表达量先下降后上升(图5),造成这一现象的可能原因是龙眼果实在刚进入低温环境时,DlLac表达受到了低温的抑制,但是随着贮藏过程的延长,DlLac基因适应了低温环境,导致在贮藏后期DlLac表达量又呈上升趋势,从而促进龙眼果皮褐变。结合荔枝漆酶基因在果皮褐变中所起作用和研究结果[7],推测龙眼果实贮藏过程中DlLac基因的上调表达可能促进了龙眼果皮褐变。
[1]孙健,李丽,游向荣,等. 荔枝、龙眼采后酶促褐变反应化学机理研究进展[J]. 南方农业学报,2012,43(10):1561-1568.
[2]陈艺晖,林艺芬,林河通,等. 龙眼采后果皮褐变因素及防褐保鲜技术研究进展[J]. 包装与食品机械,2010,28(3):27-31.
[3]Jiang Y,Duan X,Joyce D,et al. Advances in understanding of enzymatic browning in harvested litchi fruit[J]. Food Chemistry,2004,88(3):443-446.
[4]田奇琳,林玉玲,郑庆游,等. 龙眼DlLac7的克隆及其表达调控分析[J]. 果树学报,2016(10):1185-1193.
[5]万云洋,杜予民. 漆酶结构与催化机理[J]. 化学通报,2007(9):662-670.
[6]Yun-Yang W,Du Y,Fang-Xing Y,et al. Purification and characterization of hydrosoluble components from the sap of Chinese lacquer tree Rhus vernicifera[J]. International Journal of Biological Macromolecules,2006,38(3-5):232-240.
[7]刘保华,肖茜,冯超,等. 荔枝漆酶基因LcLac的克隆与表达分析[J]. 园艺学报,2012,39(5):853-860.
[8]王国栋,陈晓亚. 漆酶的性质、功能、催化机理和应用[J]. 植物学通报,2003,20(4):469-475.
[9]赵敏,魏兴东,汪春蕾,等. 细菌漆酶的研究进展[J]. 中国造纸学报,2008,23(3):107-114.
[10]左斌,王海斌,葛俊,等. 真菌漆酶基因研究进展[J]. 微生物学免疫学进展,2009,37(1):72-76.
[11]Baldrian P. Fungal laccases-occurrence and properties[J]. FEMS Microbiol Rev,2006,30(2):215-242.
[12]Ranocha P,Chabannes M,Chamayou S,et al. Laccase down-regulation causes alterations in phenolic metabolism and cell wall structure in poplar[J]. Plant Physiol,2002,129(1):145-155.
[13]Fang F,Zhang X,Luo H,et al. An Intracellular Laccase is Responsible for the Epicatechin Mediated Anthocyanin Degradation in Litchi Fruit Pericarp[J]. Plant Physiology,2015,169(4):359-2015.
[14]林河通,席玙芳,陈绍军. 龙眼果实采后失水果皮褐变与活性氧及酚类代谢的关系[J]. 植物生理与分子生物学学报,2005,31(3):287-297.
[15]帅良,李静,韩冬梅,等. 龙眼亚硫酸盐氧化酶(DlSO)基因的克隆及表达分析[J]. 果树学报,2015(1):43-50.
[16]帅良,李静,韩冬梅,等. 龙眼己糖激酶基因的克隆及原核表达[J]. 华南农业大学学报,2015,36(3):91-97.
[17]Shuai L,Li J,Niu J J,et al. Sucrose-metabolizing enzymes and their genes in the arils of two Dimocarpus longan cultivars[J]. Biologia Plantarum,2016,60(4):741-748.
[18]尤超,赵大球,梁乘榜,等. 银杏叶片RNA提取方法的研究[J]. 中国农学通报,2011,27(19):23-27.
[19]王志远.龙眼壳与龙眼核多酚的分离纯化、结构鉴定及抗氧化活性[D]. 厦门:厦门大学,2008.
[20]林河通,席玙芳,陈绍军. 龙眼果实采后失水果皮褐变与活性氧及酚类代谢的关系[J]. 植物生理与分子生物学学报,2005,31(3):287-297.
[21]罗耀红,左莹,苏钰琦,等. 漆酶对酚类化合物生物催化氧化动力学研究——以儿茶酚和表儿茶素为模式底物[J]. 林产化学与工业,2008,28(3):13-17.
Cloning and expression analysis ofthe laccase gene(DlLac)fromDimocarpuslongan
SHUAI Liang1,2,ZHAO Yu-qing1,2,+,LIAO Ling-yan1,SONG Mu-bo1,ZHU Dong-jian1,CAI Wen1,DUAN Zhen-hua1,WU Zhen-xian2,*,HAN Dong-mei3,*
(1.Institute of Food Science and Engineering Technology,Hezhou University,Hezhou 542800,China;2.College of Horticulture of South China Agricultural University,Guangdong Key Lab ofPostharvest Science of Fruit and Vegetable,Guangzhou 510642,China;3.Institute of Tree Research Guangdong Academy of Agriculture Sciences,Key Laboratory ofSouth Subtropical Fruit Biology and Genetic Resource Utilization,Ministry of Agriculture,Guangzhou 510640,China)
In the present study,laccase gene cDNA was cloned and sequenced from ‘shixia’ longan cultivar using RT-PCR and RACE,which was named asDlLac. The NCBI numbers were KY051551. TheDlLacgene consists of 1898 bp encoding a polypeptide of 576 amino acids.DlLacgene had the highest homology with the Litchi chinensis(94%)through the NCBI blastn. Amino acids of laccase gene sequence had 3 highly conservative domain structure:Cu-oxidase-3,Cu-oxidase and Cu-oxidase-2. During the normal and cold storage,combining the relationship of pericarp browning andDlLacexpression,it suggested that the laccase gene in pericarp may contribute to the postharvest longan pericarp browning.
Dimocarpuslongan;laccase;pericarp browning;gene cloning;expression
2016-12-02 +并列第一作者
帅良(1986-),男,博士,讲师,研究方向:果品采后生理,E-mail:shuailiang1212@163.com。 赵昱清(1988-),男,硕士,研究方向:果品采后生理,E-mail:zyq208@163.com。
*通讯作者:吴振先(1971-),男,博士,教授,研究方向:果品采后生理,E-mail:litchi2008@126.com。 韩冬梅(1973-),女,硕士,研究员,研究方向:果品采后生理,E-mail:handm2009@qq.com。
国家现代农业产业技术体系(荔枝龙眼)(CARS-33-14)资助;贺州学院博士启动基金(HZUBS201510);贺州市科技开发项目(贺科攻1541008);广西中青年教师基础能力提升项目(KY2016YB457);广西特聘专家专项经费;广西果蔬保鲜和深加工研究人才小高地开放课题(2016XGDSHFW02)。
TS201.3
A
1002-0306(2017)13-0095-06
10.13386/j.issn1002-0306.2017.13.018
