不同环境因子对海洋细菌Pseudoalteromonasissachenkonii HZ引导糖基转移酶基因表达的影响
2017-01-13卢晓平许艳蕊范秋苹郝鲁江
卢晓平,许艳蕊,范秋苹,郝鲁江
(齐鲁工业大学山东省微生物工程重点实验室,山东济南 250353)
不同环境因子对海洋细菌PseudoalteromonasissachenkoniiHZ引导糖基转移酶基因表达的影响
卢晓平,许艳蕊,范秋苹,郝鲁江*
(齐鲁工业大学山东省微生物工程重点实验室,山东济南 250353)
为了研究海洋细菌PseudoalteromonasissachenkoniiHZ所产多糖的生物合成基因簇,首先需要克隆到在多糖合成中起关键作用的引导糖基转移酶基因。通过设计引物,采用PCR技术从海洋细菌PseudoalteromonasissachenkoniiHZ成功克隆到该基因,并通过实时荧光定量PCR技术检测了温度、pH及海盐浓度对引导糖基转移酶基因表达量的影响。结果表明:低温有利于引导糖基转移酶的表达,随着温度的上升,引导糖基转移酶基因的表达量先增后减,在20 h时,引导糖基转移酶的表达量急剧上升,15 ℃和35 ℃引导糖基转移酶表达量分别是20 ℃表达量的5.2倍、5.8倍;海盐浓度为4.5%时引导糖基转移酶的表达量为海盐浓度3.5%的2.5倍,海盐浓度为5.5%时引导糖基转移酶的表达量为海盐浓度3.5%的2.0倍;10 h时pH为8、9的培养基中引导糖基转移酶的表达量分别为pH为7的1.64和1.67倍。该结果为探究菌体环境适应性机制提供了一定的基础。
海洋细菌PseudoalteromonasissachenkoniiHZ,引导糖基转移酶,实时荧光定量PCR
海洋环境的特殊性,赋予海洋细菌所产多糖结构的多样性以及功能的特殊性。尽管已有大量文献报道海洋细菌多糖的特殊功能,但人们对于研究生产胞外多糖的海洋细菌的兴趣方兴未艾[1]。近几年,随着对微生物多糖研究的深入,世界上微生物多糖的产量和增长量均在10%以上,而一些新兴多糖年增长量在30%以上,到目前为止已经大量投产的微生物多糖有黄原胶、结冷胶、小核菌葡聚糖、短梗霉多糖等[2]。海洋微生物处于低温、高盐、高压、寡营养、高温度梯度及高毒性浓度的特殊环境,具有产生结构新颖、功能独特的新型活性多糖的潜力[3],是开发多糖类海洋新药的重要资源,一些假交替单胞菌能够分泌胞外多糖(exopolysaccharides,EPS),EPS可以增加产生菌及某些其它海洋生物在特定环境中的存活机会。一株假交替单胞菌S9无论是在液体还是固体培养状态下稳定期都能够产生EPS,促进产生菌定居在无脊椎动物幼虫的表面[4]。Corsaro等报道P.haloplanktisTAC125能够分泌EPS,并且不同的培养温度对EPS的成分也有影响[5]。近年来,已有很多关于海洋微生物代谢产物的研究文献,但有关环境因子调控海洋细菌多糖的合成及相关基因的研究鲜见报道。
目前多种细菌产生的胞外多糖的结构已经被鉴定[6],虽然这些多糖的结构或者活性有所不同,但是多糖的生物合成途径基本相同,甚至于LPS的O抗原和合成途径相似,即通过引导糖基转移酶将单糖从糖核苷酸顺序性转移而装配到脂类载体上形成重复单元,随后是这些重复单元的聚合和输出,形成细胞表面多糖[7-8]。有些基因簇涉及特殊糖核苷酸合成的基因,但所有的基因簇都具有编码特异引导糖基转移酶的基因、参与聚合和输出过程的基因[9]。在多糖合成过程中,催化糖基转移到脂载体上的引导糖基转移酶在G+和G-均有较高的同源性,其也是胞外多糖合成的限速酶之一。因此,根据以上特性,从同源菌中设计相应的引物,从海洋细菌PseudoalteromonasissachenkoniiHZ基因组中扩增出引导糖基转移酶基因。
本研究所用菌株为皱纹盘鲍采苗板上分离得到的高产胞外多糖菌株PesudoalteromonasissachenkoniiHZ,其胞外多糖的合成与环境因子之间是否存在联系以及两者之间的相应关系还不清楚,而对该问题的研究可为阐明胞外多糖在环境适应性中的机制提供新的科学依据。
本实验室前期已对海洋细菌PseudoalteromonasissachenkoniiHZ产多糖的发酵条件进行了优化,将以此为基础研究在发酵过程中引导糖基转移酶基因的表达变化。
1 材料和方法
1.1 材料与仪器
海洋细菌Pseudoalteromonasissachenkonii502A 为实验室保藏菌种;固体培养基Zobell 2216E培养基 蛋白胨5 g/L,酵母膏1 g/L,海盐35 g/L,琼脂20 g/L,调pH7.6~7.8;种子培养基Zobell 2216E培养基 蛋白胨5 g/L,酵母膏1 g/L,海盐35 g/L,调pH7.6~7.8;发酵培养基 葡萄糖45 g/L,硫酸铵2.5 g/L,海盐45 g/L,调pH7.8~8.0;细菌基因组DNA快速抽提试剂盒、柱式细菌总RNA抽提纯化试剂盒、Low MW DNA Marker-A、DNA maker-G、Taq酶、引物 上海生工生物工程有限公司;λ-Hind Ⅲ digest、PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)、SYBR® Premix Ex TaqTMⅡ(Tli RNaseH Plus) 宝生物工程(大连)有限公司;实验所用试剂 均为分析纯。
PCR仪基因扩增仪 Bio-RAD;实时荧光定量PCR仪 Rotor-Gene Q;ZF-1B型三用紫外分析仪 上海播显科学仪器有限公司。
1.2 实验方法
1.2.1 菌种活化 取出甘油管中保藏的海洋细菌PseudoalteromonasissachenkoniiHZ,划线接种于Zobell 2216E固体培养基平板上,25 ℃静置培养24 h长出单菌落后,取单菌落再次划线培养,保存备用。
1.2.2 DNA的提取 将活化好的细菌接种于Zobell 2216E液体培养基中,25 ℃、100 r/min培养8 h。利用细菌基因组DNA快速抽提试剂盒提取DNA,经1%琼脂糖凝胶电泳检测后于-20 ℃保存备用[10]。
1.2.3 引物的设计与合成 利用Primer Primer 5.0,根据GenBank中PseudoalteromonashaloplanktisTAC125(NC_007482.1)的引导糖基转移酶设计一对引物GF(5′-ATGAAAAAAGTGCTGGTAATTGG-3′)和GR(5′-TTAATCTAATTTTTTATTTTTTTCAGCTATCC),由上海生工生物工程有限公司合成。
1.2.4 PCR反应体系与程序 PCR扩增体系为25 μL,具体组成见表1。

表1 PCR扩增体系
扩增程序为:94 ℃、5 min,94 ℃、30 s,44~51 ℃、30 s,72 ℃、2 min,35 cycle;72 ℃、10 min。其中使用温度梯度PCR扩增引导糖基转移酶基因,退火温度的选择参考引物上下游引物退火温度,PCR扩增产物经1%的琼脂糖凝胶电泳检测,取3 μL PCR产物与1 μL 6×上样缓冲液点样,70 V、40 min,利用ZF-1B型三用紫外分析仪进行观察。挑取单一明亮条带的PCR产物送往上海生工有限公司进行测序,得到序列后,将序列输入到NCBI数据库(http://blast.ncbi.nlm.gov)进行序列比对,比较引导糖基转移酶序列的同源性。
1.2.5 温度变化对引导糖基转移酶基因表达的影响 将海洋细菌PseudoalteromonasissachenkoniiHZ接种到种子培养基中,25 ℃、100 r/min培养8 h。按照8%的接种量接种到发酵培养基中,分别在15、20、25、30、35 ℃的恒温培养振荡器中培养,并在10、20、34、44 h分别取样,备用。
1.2.6 海盐浓度对引导糖基转移酶基因表达的影响 将海洋细菌PseudoalteromonasissachenkoniiHZ种子液接种到海盐浓度分别为3.5%、4.5%、5.5%的发酵培养基中,置于25 ℃恒温培养振荡器中培养,并在10、20、34 h分别取样,备用。
1.2.7 pH对引导糖基转移酶基因表达的影响 pH对引导糖基转移酶基因表达的影响 将海洋细菌PseudoalteromonasissachenkoniiHZ种子液接种到pH分别为7、8、9的发酵培养基中,置于25 ℃恒温培养振荡器中培养,并在10、20、34 h分别取样,备用。
1.2.8 RNA的提取及cDNA的反转录合成 使用柱式细菌总RNA提取纯化试剂盒提取细菌的RNA,使用PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)合成cDNA第一链,具体操作参照使用说明书。
1.2.9 荧光定量PCR体系的建立 以测得的引导糖基转移酶序列为靶序列,运用primer primer5.0软件设计一对荧光定量PCR引物SGF(5′-TTTGGCTGGC GTGTAGATAA-3′)和SGR(5′-GCGTTTTGCTATGT CTGAATG-3′)。以16S rDNA为内参基因,引物序列为16S rDNA-F(5′-TGTAGCGGTGAAATGCGTA-3′)和16S rDNA-R(5′-AGGGTATCTAATCCTGTTTGCTC-3′)[11]。并进行引物特异性检测。PCR体系为25 μL,见表2。

表2 荧光定量PCR体系
扩增程序:95 ℃、30 s;95 ℃、5 s,54 ℃、30 s,72 ℃、30 s,50次循环;溶解曲线70~99 ℃。反应在QIAGEN Rotor-GeneQ荧光定量PCR仪上进行,以上反应条件均经过了一系列温度梯度PCR优化[12],每个浓度梯度同时做三个平行重复,以不加模板的反应液为空白对照。
1.2.10 实验数据的处理 使用荧光定量PCR仪附带软件采用2-ΔΔCT算法对实验数据进行分析处理[13],并导出内参基因与目的基因的扩增曲线、溶解曲线。
2 结果与分析
2.1 基因组DNA的提取
利用细菌基因组DNA快速抽提试剂盒提取海洋细菌PseudoalteromonasissachenkoniiHZ基因组DNA,经琼脂糖凝胶电泳,结果如图1所示:位于23 kb处有一明亮条带,可用于后续PCR反应等。

图1 提取的细菌基因组DNA Fig.1 Extracted bacterial genome DNA注:M:λ-Hind Ⅲ digest。
2.2 引导糖基转移酶基因的扩增
以提取的海洋细菌PseudoalteromonasissachenkoniiHZ DNA为模板,以GF和GR为引物进行PCR扩增,结果如图2。
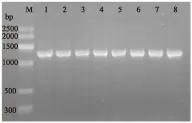
图2 引导糖基转移酶基因的扩增Fig.2 Glycosyltransferase gene amplification注:M为DNA maker-G,泳道1~8为44~51 ℃下所扩增的引导糖基转移酶基因。
图2中泳道8中的PCR产物明亮、条带单一,无引物二聚体,因此将泳道8中的PCR产物送往上海生工生物工程有限公司进行测序。将翻译得到的蛋白质序列在NCBI中进行比对,其与Pseudoalteromonassp.MW4237引导糖基转移酶的蛋白质序列相似度为89%,与Pseudoalteromonassp.P1-11相似序列为88%,与Pseudoalteromonasaqarivorans相似度为87%,推测该序列为引导糖基转移酶序列。
2.3 细菌总RNA的提取
使用柱式细菌总RNA提取纯化试剂盒提取细菌的RNA,电泳结果如图3。由图3可知提取的RNA条带完整,满足下一步实验要求。电泳结束后参照使用说明书合成cDNA第一链。

图3 细菌总RNA的提取Fig.3 Extraction of total RNA from bacteria
2.4 引物特异性分析
以合成的cDNA为模板,以SGF和SGR为引导糖基转移酶保守片段上下游引物,以16S rDNA-F和16S rDNA-R为内参基因上下游引物,进行普通PCR扩增,检测引物特异性。结果见图4,可知产物单一,无引物二聚体,可进行下一步实验。
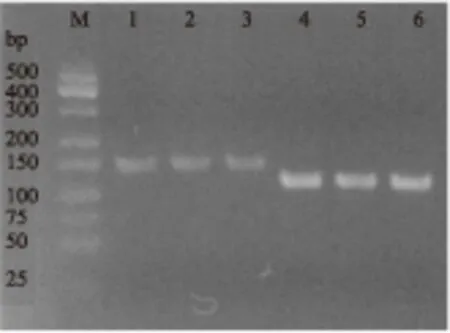
图4 普通PCR引物特异性分析Fig.4 Normal PCR primers specific analysis注:M-Low MW DNA Marker-A;1~3泳道为引导糖基转移酶保守片段;4~6泳道为内参16S rRNA片段。
由图4可知,SGF/SGR和16S rDNA-F/16S rDNA-R均能扩增出单一条带且无引物二聚体,引物特异性良好,使用该两对引物进行荧光定量PCR实验,溶解曲线(图5和图6)显示只有单一吸收峰,表明引物特异性良好。
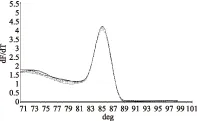
图5 引导糖基转移酶溶解曲线Fig.5 Glycosyltransferase dissolution profile注:不同形状的曲线表示各孔中引导糖基转移酶基因的扩增。

图6 16S rRNA溶解曲线Fig.6 16S rRNA dissolution profile注:不同形状的曲线表示各孔中16S rRNA基因的扩增。
2.5 引导糖基转移酶基因对温度变化的响应
以16S rDNA为内参基因,引导糖基转移酶为目的基因,以10 h取样的25 ℃作为对照组,采用2-ΔΔCT算法计算不同温度下目的基因的相对表达量。结果如图7所示。

图7 温度变化对引导糖基转移酶的影响Fig.7 Temperature changes affect the glycosyltransferase
PseudoalteromonasissachenkoniiHZ在不同温度下培养20 h后,引导糖基转移酶的表达量随温度的上升而增加。在20 h时,引导糖基转移酶的表达量急剧上升,15 ℃和35 ℃引导糖基转移酶表达量分别是20 ℃表达量的5.2倍、5.8倍。34 h和44 h时引导糖基转移酶的表达量差异不大。
2.6 引导糖基转移酶对海盐浓度变化的响应
以16S rDNA为内参基因,引导糖基转移酶为目的基因,以10 h取样的海盐浓度4.5%作为对照组,采用2-ΔΔCT算法计算不同海盐浓度下目的基因的相对表达量,结果如图8所示。
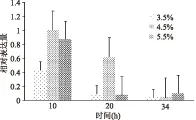
图8 海盐浓度对引导糖基转移酶表达的影响Fig.8 Salt concentration changes affect the glycosyltransferase
在10 h时,海盐浓度为4.5%引导糖基转移酶的表达量为海盐浓度3.5%的2.5倍,海盐浓度为5.5%引导糖基转移酶的表达量为海盐浓度3.5%的2.0倍,10 h之后引导糖基转移酶的表达量都减少,34 h后不同海盐浓度中引导糖基转移酶的表达量无明显变化。此外引导糖基转移酶的表达情况也与前期发酵培养基优化参数相吻合,从分子角度证明了最优培养基中海盐浓度的选择,即当海盐浓度为4.5%时,培养基中多糖产量最大。
2.7 引导糖基转移酶对pH变化的响应
以16S rDNA为内参基因,引导糖基转移酶为目的基因,以10 h取样的pH8作为对照组,采用2-ΔΔCT算法计算不同pH条件下目的基因的相对表达量。pH变化对引导糖基转移酶表达的影响如图9所示。

图9 pH变化对引导糖基转移酶表达的影响Fig.9 pH changes affect the glycosyltransferase
pH对引导糖基转移酶的影响与海盐浓度对引导糖基转移酶的影响相似,即在10 h时pH为8和pH为9的培养基中引导糖基转移酶的表达量分别为pH为7的1.64和1.67倍。随着培养时间的延长,引导糖基转移酶的表达量都降低。
3 讨论与结论
在海冰和海水界面存在着大量产生胞外多糖的细菌,高浓度的胞外多糖使得细菌在恶劣的寒冬条件和高盐度中生存,并对冰晶对细菌的伤害起到缓冲作用[14]。Mancuso等从南极海冰和海水中分离得到两株海洋细菌CAM025和CAM036,经过16S rDNA鉴定为Pseudoalteromonas属,对其胞外多糖研究发现,其在-2、10 ℃时胞外多糖的产量是20 ℃的30倍[15]。李江等考察了温度对多糖合成关键酶基因ugd的影响,也发现低温有利于胞外多糖的合成,其在2、10 ℃时基因的表达量是20 ℃的4倍[11]。这些研究结果支持了胞外多糖在低温下对微生物区系具有重要作用。
胞外多糖在细菌极端环境适应性中具有重要作用,并引起了越来越多学者的关注。细菌胞外多糖的生物合成的产量受外部因素的影响,因此通过荧光定量PCR检测了温度、pH、海盐浓度对引导糖基转移酶表达的影响,发现低温有利于引导糖基转移酶的表达,在不同培养温度下培养20 h时,引导糖基转移酶的表达量急剧上升,15 ℃引导糖基转移酶表达量是20 ℃表达量的5.2倍,由此可知细菌在低温环境下会通过大量分泌胞外多糖来适应环境,免受冰晶的伤害,盐度和pH的改变也会影响引导糖基转移酶的表达,通过提高引导糖基转移酶基因的表达量来促进细菌胞外多糖的合成量,以缓冲盐度和pH对菌体的胁迫作用,通过控制胞外多糖的合成可能是菌体环境适应性的策略之一。
[1]Chi Zhenming,FangYan.Exopolysaccharides from Marine
Bacteria[J].Journal of Ocean University of China,2005(4):67-74.
[2]崔艳红,黄现清.微生物胞外多糖研究进展[J].生物技术通报,2006(2):25-29.
[3]孙红梅,张星星,白玉,等.海洋假单胞菌胞外多糖的抗氧化性[J].大连工业大学学报,2012(31):395-398.
[4]Holmström C,Kjelleberg S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents[J]. FEMS Microbiology Ecology,1999,30:285-293.
[5]Corsaro MM,Lanzetta R,Parrill iE,et al. Influence of growth temperature on lipid and phosphate contents of surface polysaccharides from the antarctic bacterium Pseudoalteromonas haloplanktis TAC 125[J].Journal of Bacteriology,2004,186(1):29-34.
[6]张珊珊,王长云,魏晓蕾,等.海洋微生物胞外多糖结构与生物活性研究进展[J].微生物学通报,2007(34):153-156.
[7]Katsuhide Miyake,shinji Iijima.Bacterial capsular polysaccharide and sugar transferases[J].Adv Biochem Engin/Biotechnol,2004,90:89-111.
[8]Kolkman MA,van der Zeijst BA,Nuijten PJ.Function analysis of glycosyltransferases encoded by the capsular polysaccharide biosynthesis locus of streptococcus pneumoniae serotype 14[J]. Journal of Biological Chemistry,1997,272(31):19502-19508.
[9]Taylor WH,Juni E.Pathways for biosynthesis of a bacterial capsular polysaccharide I characterization of the organism and polysaccharide[J].Journal of Acteriology,1961,81(5):688-693.
[10]姆布鲁克等著.黄培堂等译.分子克隆实验指南(上册)/(美)[M]. 3版. 北京:科学出版社,2002:385-387.
[11]李江,沈继红,何培青,等.环境因子对南极菌Pseudoalteromonassp. S-15-13多糖合成关键酶UGD基因表达的影响[J].海洋与湖沼,2010,41(6):824-828.
[12]Torben LS,Niels BR,Carola H,et al.Real-time quantitative PCR for assessment of abundance of Pseudoalteromonas species in marine samples[J].Applied and Environmental Microbiology,2004,70(4):2373-2382.
[13]Kenneth JL,Thomas DS.Analysis of Relative Gene Expression Data Using Real-time Quantitative PCR and the 2-ΔΔCTMethod[J].METHODS,2001(25):402-408.
[14]Krembs C,Eicken H,Junge K,et al.High concentrations of exopolymeric substances in Arctic winter sea ice:implications for the polar ocean carbon cycle and cryoprotection of diatoms[J]. Deep Sea Research,2002,49(12):2163-2181.
[15]Mancuso CA,Garon S,Bowman JP,et al.Production of exopolysaccharides by Antarctic marine bacterial isolates[J]. Journal of Applied Microbiology,2004,96(5):1057-1066.
The impact of environmental factors on marine bacteriaPseudoalteromonasissachenkoniiHZ glycosyltransferase gene expression
LU Xiao-ping,XU Yan-rui,FAN Qiu-ping,HAO Lu-jiang*
(Qilu University of Technology,Shandong Provincial Key Laboratory of Microbial Engineering,Jinan 250353,China)
To study the biosynthesis gene cluster of the polysaccharide produced by marine bacteriumPseudoalteromonasissachenkoniiHZ,and to clone the gene which plays a key role in the synthesis of polysaccharide. Glycosyltransferase gene was amplified by designing primers using PCR technology from marine bacteriaPseudoalteromonasissachenkoniiHZ,and examined the effect of temperature,pH and salt concentration of glycosyltransferase gene expression by real-time fluorescence quantitative PCR. The results showed that the low temperature was in favor of the expression of the glycosyltransferase. With the increase of the temperature,the expression of the gene of the glycosyltransferase increased first and then decreased. It was showed that glycosyltransferase gene expression level increased rapidly,and the glycosyltransferase gene expression level ofPseudoalteromonasissachenkoniiHZ grown at 15 and 35 ℃ were 5.2,5.8 times higher than grown at 20 ℃,respectively. 4.5% salt concentration could increase the expression level of glycosyltransferase gene 2.5 times compared to that of 3.5% salt concentration,while 5.5% salt concentration glycosyltransferase gene expression level was 2 times compared to 3.5% salt concentration. The glycosyltransferase gene expression level ofPseudoalteromonasissachenkoniiHZ grown at pH8 and 9 were 1.64、1.67 times higher than grown at pH7,respectively. The research on glycosyltransferase gene ofPseudoalteromonasissachenkoniiHZ laid a foundation for exploring the mechanism of adaptation to the environment.
marine bacteriaPseudoalteromonasissachenkoniiHZ;glycosyltransferase;real-time PCR
2016-04-14
卢晓平(1992-),男,硕士研究生,研究方向:微生物酶资源的开发与利用,E-mail:15288863082@163.com。
*通讯作者:郝鲁江(1972-),男,博士,副教授,研究方向:海洋细菌活性物质的筛选,微生物酶资源的开发,E-mail:lujiang_hao@163.com。
山东省自然科学基金(ZR2012CM019)。
TS201.1
B
1002-0306(2016)22-0212-05
10.13386/j.issn1002-0306.2016.22.033
