不同水分环境下小麦株高性状QTL定位分析
2016-10-14任艳云王世充邵敏敏孙雷明黄玲赵凯徐兴科王继峰冯维营王霖
任艳云 王世充 邵敏敏 孙雷明 黄玲 赵凯 徐兴科 王继峰 冯维营 王霖
摘要:为了发掘控制小麦抗旱节水相关性状的基因位点,为抗旱节水品种选育提供指导,本研究利用洛旱2号/潍麦8号的 F8∶9群体的302 份材料,在6个不同水分环境条件下进行试验,对小麦抗旱相关株高性状进行了 QTL 分析。结果显示,在所有环境中共检测到 17个控制株高的加性QTL,主要分布在 2B、3A、3B、4A、5A、5D、6B、7A 和 7B 染色体上,加性效应值为1.9196~5.3828 cm, 可解释3.3160%~26.9489%的表型变异。在充分灌溉条件下的E1、E2和E3三个环境中共有14个QTL位点,25次被检测到;在干旱胁迫环境下的E4、E5和E6三个环境中共有5个QTL位点,7次被检测到。在所有检测到的17个QTL位点中,有12个位点只在灌溉环境下被检测到,有3个位点只在干旱胁迫环境下被检测到,只有2个位点在灌溉和干旱胁迫环境下同时被检测到。表明基因的表达受环境条件的影响较大,在QTL水平上表明基因与环境之間存在互作。定位在7A染色体上位于Xbarc049和Xgwm273之间的qPh-WL-7A.3,在三个水分胁迫环境下稳定表达,能增加株高2.4815~3.6972 cm,加性效应贡献率在8.6426%~10.0711%,是一个与水分高效利用密切相关的QTL,可用于小麦抗旱基因改良及分子标记辅助育种。
关键词:小麦;株高;QTL;干旱胁迫
中图分类号:S512.103.2文献标识号:A文章编号:1001-4942(2016)09-0010-07
AbstractPlant height is one of the most important traits that associated with drought resistance of wheat. In order to explore the gene sites associated with drought resistance and water saving, and provide guidance for drought-resistant breeding of wheat, the plant height QTL was evaluated with the population of 302 recombinant inbred lines derived from a cross between drought-tolerant cultivar Luohan 2 and water-sensitive cultivar Weimai 8 in 6 different water regimes. Total 17 additive QTL loci for plant height were identified, and they distributed on chromosomes 2B, 3A, 3B, 4A, 5A, 5D, 6B, 7A and 7B, respectively. These QTL loci could explain the phenotypic variations varied from 3.3160%~26.9489% with the additive effects of 1.9196~5.3828 cm. Of which, 14 QTL loci were detected for 25 times in 3 irrigation environments (E1, E2 and E3), and 5 QTL loci were detected for 7 times in 3 drought stress environments (E4, E5 and E6). Of all the 17 QTL loci for plant height, 12 QTL loci were detected only in irrigation environment, 3 QTL loci were detected only in drought stress environment, and 2 QTL loci were detected in both environments. It suggested that the expression of these genes were greatly influenced by environmental condition, and showed that there was interaction between gene expression and environment on the level of QTL. qPh-WL-7A.3, which was located between Xbarc049 and Xgwm273 on 7A, could stably express in all the three drought stress environments, and increase plant height for 2.4815~3.6972 cm with the contributions of additive effects as 8.6426%~10.0711%. It indicated that the QTL was closely related to water efficient utilization and could be used in the genetic improvement of drought tolerance and molecular marker assisted breeding of wheat.
KeywordsWheat; Plant height; QTL; Drought stress
小麦是我国第三大粮食作物,主要分布在我国部分干旱、半干旱地区,水分胁迫一直是影响我国北方小麦生产的最主要因素[1]。小麦茎秆是支撑器官,支撑叶片和穗部器官。合理的叶片着生部位及着生角度,使群体冠层结构合理,有利于光合作用,提高群体生物合成总量。因此,株高影响着小麦的种植密度、株型、冠层结构、抗倒性能和收获指数,是影响小麦产量的最重要因素之一[2]。研究表明,水分环境对小麦株高表型变异具有显著影响,水分亏缺通常导致小麦株高和产量显著降低[3,4]。在干旱胁迫条件下,小麦维持较高株高可增加花前茎秆碳水化合物贮存量和花后茎秆代谢向籽粒转运量,有利于籽粒灌浆和增加粒重,从而有效补偿产量的损失[5],整体表现出小麦株高与产量及其产量构成因素间不同程度的正相关性[5-7]。株高不仅是对小麦基因型本身的反映,也是小麦从出苗到成熟整个生长过程对环境适应性反应的结果,大量研究表明,干旱胁迫下的株高与小麦的抗旱性呈高度正相关,是鉴定小麦抗旱性的可靠指标[8,9]。在干旱环境下,不仅小麦的最终株高(90~105 cm)与抗旱性呈高度正相关,拔节期的苗高也与抗旱性呈极显著正相关。但小麦株高与其抗旱性之间的正相关关系有一定的限制,超过一定限度这种关系就会丧失[10]。因此,株高评价是小麦抗旱性的重要指标,合理株高是旱地小麦获得较高产量的重要结构基础[3]。
大量研究表明,小麦株高属于多基因控制的复杂数量性状,主要受加性、上位性及环境互作效应调控[4,6,7,10-12]。迄今为止,通过突变体结合分子生物学手段已发现 25个矮秆基因(Rht);此外借助分子遗传学方法,发现了大量的控制株高的数量性状位点(QTL),这些 QTL 几乎分布在小麦整个基因组上[2,4,6,7,10-18]。因此利用不同遗传背景的作图群体,借助分子数量遗传学方法,发掘在干旱条件下遗传效应值稳定表达、能够增加株高且在水分充足条件下不表达的数量性状位点,对稳定株高、增加植株的抗旱性、提高产量及分子标记辅助选育抗旱小麦品种具有重要意义[12]。為此,本研究通过抗旱性差异较大的 2 个冬小麦品种洛旱2号(耐旱型) 与 潍麦8号(水分敏感型) 杂交创建的RIL群体为材料,在不同水分环境下对小麦株高进行 QTL 定位剖析,旨在为小麦株高抗旱遗传改良和分子标记辅助选择育种奠定理论基础。
1材料与方法
1.1试验材料
亲本洛旱2号为多穗型抗旱小麦品种,由洛阳市农业科学研究院选育而成;潍麦8号为高产大穗水分敏感型小麦品种,由潍坊市农业科学院选育;以及由二者杂交经多代自交获得的302个重组自交系(RIL,F8∶9)群体,简称WL。
1.2试验设计
将WL群体及其亲本于2010-2011、2011-2012、2013-2014年度种植于济宁市农业科学研究院试验农场(环境1、2、3,E1、E2、E3),采用灌溉模式,冬前浇越冬水,春季浇拔节水和灌浆水,其它管理同一般大田。2012-2013年度种植在济宁市农业科学研究院试验农场和临沂市农业科学研究院试验农场,采用控水模式,足墒播种,出苗后不浇水。其中济宁试点设置两个处理,一个为覆膜(环境4,E4),另一个为不覆膜(环境5,E5),临沂试点不覆膜(环境6,E6),其余非水分管理措施同一般大田。每个家系和亲本种植2行,行距30 cm,株距4 cm,行长2 m,每个环境1次重复,随机排列。
1.3表型性状调查和分析
于小麦开花15 d后,用直尺量取两个亲本及RIL家系的平均高度(从地面到穗的顶部,不连芒),5点平均作为平均株高。成熟后每个亲本及各RIL家系随机取5个单株,量取主茎高度(从分蘖节到穗的顶部,不连芒),取平均值作为主茎株高。
1.4遗传图谱的构建及QTL分析方法
应用MapMaker/EXP 3.0软件构建连锁图谱,该图谱包含348个位点,分布在23个连锁群上,基因组全长3 132.2 cM,标记间的平均距离为9.0 cM。A基组长1 086.1 cM、B基组长1 170.8 cM,D基组长875.2 cM。单个染色体上的标记数目相差较大,以7B染色体上标记最多,为47个,3D上最少,只有2个。染色体长度以7B最长,为226.6 cM,3D最短为36.7 cM。7B染色体标记间的平均距离最短,为3.2 cM,6D染色体标记间的平均距离最长为19.9 cM。4D和6D各包含1个连锁断点[19]。
采用 SPSS 13.0软件对表型数据进行正态分布分析。应 用 完 备 区 间 作图软件IciMapping v3.0 (http://www.isbreeding.net/) 进行 QTL 分析。LOD 阈值设定为 2.5,步长为 1 cM,进行1 000次置换检测。对6个环境的平均株高和4个环境的主茎株高进行了QTL分析。
2结果与分析
2.1小麦平均株高及主茎高度的表型变异
RIL群体及其亲本的株高参数如表1所示。在不同环境条件下,洛旱2号的株高显著低于潍麦8号,RIL群体的株高平均值显著高于双亲平均值,明显的表现为倾高亲遗传。RIL群体的株高峰度系数和偏度系数绝对值都小于1,表现为
2.2株高QTL鉴定
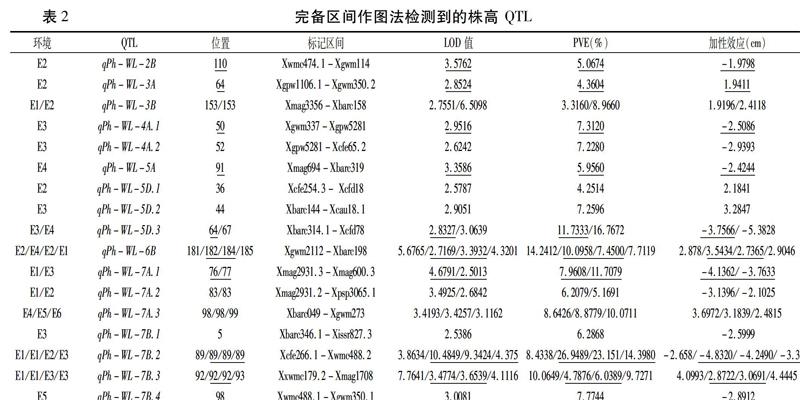
在不同环境条件下,从小麦RIL群体中共检测到 17个控制株高的加性QTL,主要分布在 2B、3A、3B、4A、5A、5D、6B、7A 和 7B 染色体上(表 2、图 1) 。有 7个加性QTL 位点的加性效应来自于高值亲本潍麦8号,具有增加株高的效应,加性效应值为 1.9196~4.4445 cm,单个QTL可解释3.3160%~14.2412%的表型变异; 其余10 个加性QTL 来自于低值亲本洛旱2号,具有降低株高的效应,加性效应值为 1.9798~5.3828 cm,可解释5.0674%~26.9489%的表型变异。在6个环境共检测到的32个点次加性 QTL中,有 10点次(涉及到6个QTL位点)可解释超过 10% 的表型变异, 18点次(涉及到14个QTL位点)可解释5%~10% 的表型变异。
2.3平均株高和主茎株高QTL分析
在E1、E2、E3和E4四个环境中共有15个株高QTL位点,28点次被检测到(E5、E6两个环境没检测主茎株高QTL)。其中有9个主茎株高QTL位点, 14点次被检测到,分别位于2B、3A、4A、5A、5D、6B、7A、7B染色体上;有10个平均株高QTL位点,14点次被检测到,分别位于3B、4A、5D、6B、7A、7B染色体上。检测到的两类QTL位点数及总次数大体相当。在这些QTL位点中,有4个被同时检测到,占总数的26.7%,有5个位点只在主茎株高中被检测到,占总数的33.3%,有6个位点只在平均株高中被检测到,占总数的40.0%。
在某一特定环境中检测到的主茎株高QTL和平均株高QTL的数目和位置不尽相同,大多数不能在相同位点被同时检测到,只有少数主茎株高QTL和平均株高QTL被同时定位在染色体上相同位置。在那些没被同时检测到的QTL中,有些可以在其他环境下相同的位点被检测到。如在E1环境中检测到的5个平均株高QTL中,有2个(qPh-WL-7B.2 和qPh-WL-7B.3)在同环境下与检测到的控制主茎高度的QTL定位在相同区间,有2个(qPh-WL-7A.2 和qPh-WL-3B)在E2环境中被检测到,另有1个(qPh-WL-6B)在E2和E4环境中作为主茎株高被定位在相同的区间。
这说明株高QTL调控株高发育是比较多样性的,一部分QTL同时对主茎和分蘖的发育都起一定的调控作用,另外一部分QTL对主茎调控作用强,还有一部分QTL对分蘖的调控作用强,且受环境条件的影响。在一定环境条件下二者同时表达,控制株高发育,但当环境条件改变后,原来对主茎调控作用强的QTL可能变弱,甚至不能检测到,而对分蘖的调控作用变强;原来对分蘖调控作用强的部分QTL可能变弱,而对主茎的调控作用变强。同时也有某些QTL只对主茎或者分蘖的发育起调控作用。这部分QTL解释了为什么有的小麦品种穗层整齐,主茎和分蘖高度相差較小,有的品种穗层不齐,主茎和分蘖高度差异较大的原因。
2.4小麦株高QTL在基因组中分布的不均匀性及其与水分环境互作
由图 1 看出,在不同水分环境条件下,控制小麦株高的加性QTL在不同染色体间和同一染色体内的不同标记区段上呈现出明显的不均匀分布,其中在 5D、7A 和7B上分布最多,均达3~4个。这些QTL在特定标记区间的聚集分布形成了 QTL 热点区域,如 5D染色体Xcfe254-Xcfd78,7A 染色体Xmag2931-Xgwm273 和 7B 染色体 Xcfe266-Xgwm350 区间。说明,小麦株高QTL在基因组中并不是均匀分布,在某些特定区间可能集中携带大量控制株高的QTL。
在所有的17个加性QTL中,有3个位点被4次定位在相同的区间,有1个位点被重复检测到3次,有4个位点被重复检测到2次,其余QTL 均为环境特异表达。在被检测到4次的3个QTL中,有的位于染色体相同的位置,有的位置稍有不同。如在7B染色体上Xcfe266.1-Xwmc488.2 区间的QTL,4次定位的位置完全相同,但同在同条染色体上Xwmc179.2-Xmag1708 区间定位到的QTL,有3次定位在相同的位置,另外1次定位在相差1 cM的位置。这些位点因环境不同,对株高表型变异的贡献率也有显著差异,如qPh-WL-7B.2,对株高表型变异的贡献率为8.4338%~26.9489%,说明这些重要位点尽管在多环境中能稳定表达,但其表达形式和强度仍存在较强的环境依赖性。
从表2中可以看出,不同环境中检测到的QTL数目也不相同,6个环境中检测到的QTL 数量差异较大。在 E1、E2和E3环境中分别有8个QTL 被检测到,而在E4、E5 和E6环境中分别只检测到4、2个和1个 QTL,同样表明基因的表达
受环境条件的影响较大,这在QTL水平上表明基因与环境之间存在互作。在控水和充分灌溉两种环境下检测到的QTL位点数和次数也有明显的差异。在控水环境下检测到的株高QTL明显少于充分灌溉环境下的数量。在充分灌溉条件下,E1、E2和E3三个环境中共有14个QTL位点,25次被检测到;在限制水分环境下的E4、E5和E6三个环境中共有5个位点,7次被检测到。在所有检测到的17个位点中,有12个位点只在灌溉环境下被检测到,有3个位点只在限制水分环境下被检测到,只有2个位点在充分灌溉和水分胁迫环境下同时被检测到。
3讨论与结论
小麦抗旱性是由多基因控制的复杂数量性状,受到许多形态结构性状、生长发育性状、产量构成性状和生理生化性状的直接或间接影响,利用普通的细胞遗传和数量遗传学方法难以进行深入研究[18]。近年来,随着分子生物技术的快速发展和不断完善,分子遗传学与传统数量遗传学的交叉结合、各类作物分子标记遗传连锁图谱的构建、比较基因组学及相关的分子遗传学研究为作物抗旱性改良提供了新的机遇。通过对与抗旱相关基因紧密连锁的分子标记的鉴定分析,可以把这一复杂的数量性状分解成为单个的孟德尔因子进行分析和选择。一旦找到与抗旱节水相关基因紧密连锁的标记,就可以在育种工作中进行标记辅助选择,大大提高选择效率[20]。
大量研究表明,小麦株高是小麦抗旱性的一个重要指标,通过不同遗传背景和不同环境条件下的QTL定位分析,可以为分子标记辅助育种提供理论支撑[12,21]。本研究通过在不同水分环境下的株高QTL定位分析发现,水分对株高基因的表达影响很大,部分在水分充足条件下效应值较高的QTL在水分匮缺的环境下效应值变小,甚至不表达,这也许是水分匮缺条件下株高明显降低的主要原因之一。但株高过矮,不利于小麦群体的建成,不利于群体光合作用和有机物的积累,不利于产量的提高,所以,在抗旱育种中那些在水分充足和匮乏环境下同时表达的QTL对干旱条件下小麦株高的建成具有较重要的意义,特别是那些在水分充足时未被检测到,只在干旱条件下被检测到的QTL,对在干旱胁迫下小麦株高的建成、增加碳水化合物的积累、提高产量具有非常重要的意义。
本研究定位到7A染色体上Xbarc049和Xgwm273之间的qPh-WL-7A.3,在三个水分匮乏的环境下稳定表达,株高增加2.4815~3.6972 cm,加性效应贡献率都在8.6426%~10.0711%,是一个与水分高效利用密切相关的QTL。梁子英等[22]以小麦品系“0911-46”与品系“42”杂交获得的F2及其衍生F2∶3群体为材料,应用SSR标记构建连锁图谱,在7A染色体Xbarc336-Xwmc607区间检测到控制株高的QTL。叶亚琼等[12]以冬小麦重组近交系群体[陇鉴19(耐旱)×Q9086(水分敏感)]采用条件复合区间作图法对4个环境不同水分条件下的株高进行QTL定位分析,在7A上发现了1个在多环境表达的加性QTL。张正斌等[23]关于小麦水分利用效率(WUE)改良的生理遗传研究表明,小麦7AS 染色体臂上载有高WUE 基因。本研究定位到的这个QTL与这些QTL处于相似位置,有可能是同一个QTL,进一步相互验证了试验结果的真实性。该研究结果为进一步进行重要抗旱基因的精细定位、克隆及 QTL 分子标记辅助育种奠定了基础。
参考文献:
[1]吴振录,黄光宏,樊哲儒,等. 小麦水分高效利用种质的筛选方法探讨[J]. 麦类作物学报,2005,25(5):143-146.
[2]刘兆晔,于经川,辛庆国,等. 小麦株高问题的探讨[J].山东农业科学,2014,46(3):130-134
[3]武仙山,王正航,昌小平,等.用株高旱胁迫系数分析小麦发育中的抗旱性动态[J].作物学报,2008,34(11):2010-2018.
[4]Wang Z H,Wu X S,Ren Q,et al.QTL mapping for developmental behavior of plant height in wheat (Triticum aestivum L.) [J].Euphytica,2010,174(3):447-458.
[5]Yang J,Zhang J,Liu L,et al.Water deficit-induced senescence and its relationship to the remobilization of prestored carbon in wheat during grain filling [J].Agronomy Journal,2001,93(1):196-206.
[6]Wu X,Wang Z,Chang X,et al. Genetic dissection of the developmental behavior of plant height in wheat under diverse water regimes [J].Journal of Experimental Botany,2010,61(11):2923-2937.
[7]Cui F,Li J,Ding A,et al. Conditional QTL mapping for plant height with respect to the length of the spike and internode in two mapping populations of wheat [J]. Theoretical and Applied Genetics,2011,122(8):1517-1536.
[8]王晨阳,毛凤梧,周继泽,等.不同土壤水分含量对小麦籽粒灌浆的影响[J]. 河南职技师院学报,1992, 20(14):17-22.
[9]周桂莲.小麦抗旱性鉴定的形态学指标及其分析评价[J]. 陕西农业科学, 1996(4):33-34
[10]王彦梅,张正斌,刘昌明,等. 河北省抗旱节水小麦生产现状及育种对策[J]. 中国生态农业学报,2004,12(4):142-144.
[11]Cadalen T,Sourdille P,Charmet G,et al.Molecular markers linked to genes affecting plant height in wheat using a doubled-haploid population [J].Theoretical and Applied Genetics,1998,96(6):933-940.
[12]叶亚琼,栗孟飞,刘媛,等. 小麦株高QTL定位及其水分环境互作遗传分析 [J]. 华北农学报,2015,30(5):83-91.
[13]Sourdille P,Cadalen T,Guyomarch H, et al. An update of the Courtot×Chinese Spring intervarietal molecular marker linkage map for the QTL detection of agronomic traits in wheat [J].Theoretical and Applied Genetics,2003,106 (3) : 530-538.
[14]周淼平,黄益洪,任丽娟,等.利用重组自交系检测小麦株高的QTL [J].江苏农业学报,2004, 20(4): 201-206.
[15]刘冬成,高睦枪,关荣霞,等.小麦株高性状的QTL分析[J].遗传学报,2002,29(8) : 706-711.
[16]Zhang K,Tian J,Zhao L,et al.Mapping QTLs with epistatic effects and QTL×environment interactions for plant height using a doubled haploid population in cultivated wheat [J].Journal of Genetics and Genomics,2008,35(2):119-127.
[17]Wang L, Cui F, Ding A M, et al. Length of internode of and spike:how do they contribute to plant height of wheat at an individual QTL level? [J]. Cereal Research Communications, 41(1): 54-65.
[18]张瑞霞. 小麦抗旱相关农艺性状的QTL分析 [D]. 保定:河北农业大学,2009.
[19]王霖,冯维营,黄玲,等. 小麦容重 QTL 定位 [J]. 山东农业科学, 2014,46(4):24-27.
[20]黄清华,曹连莆,张新忠,等. 数量性状定位及作物抗旱性分析研究进展[J]. 安徽农业科学, 2007, 35(21):6386-6388.
[21]丁安明,崔法,李君,等. 小麦单株产量与株高的QTL分析 [J]. 中国农业科学,2011,44(14):2857-2867.
[22]梁子英,李美霞,王竹林,等.小麦株高相关性状的QTL分析 [J]. 西北农业学报,2014,23(6): 64-72.
[23]张正斌,山仑,徐旗. 控制小麦种属旗叶水分利用效率的染色体背景分析 [J]. 遗传学报,2000, 27(3):240-246. (上接第9頁)
[6]司爱君, 阮孟斌, 谢宗铭. 改良热酚法快速提取棉花不同组织RNA[J]. 安徽农业科学, 2012, 40(16):8830-8832.
