大洋桥石藻Gephyrocapsaoceanica对海水酸化与营养盐胁迫的复合响应
2016-06-20蔡小霞于培松张海峰胡佶张海娜潘建明国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室浙江杭州3002
蔡小霞,于培松,张海峰,胡佶,张海娜,潘建明*(.国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州3002)
大洋桥石藻Gephyrocapsaoceanica对海水酸化与营养盐胁迫的复合响应
蔡小霞1,于培松1,张海峰1,胡佶1,张海娜1,潘建明1*
(1.国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州310012)
摘要:本文选取了大洋桥石藻(Gephyrocapsaoceanica)作为实验对象,通过C O2加富实验模拟海水酸化环境,分析了氮充足和氮相对不足条件下海洋颗石藻对海水酸化的生理响应。结果发现在两种营养盐状态下,C O2加富均能一定程度促进大洋桥石藻种群增长与Chla浓度的增加。对比两种营养盐状态,氮相对不足条件下的大洋桥石藻细胞密度和叶绿素含量均有最显著的提高,表明低营养盐浓度和低的氮磷比可能更有利于大洋桥石藻的生长繁殖。电镜观测结果显示酸化对大洋桥石藻的钙化作用具有显著的负影响,并且在氮相对不足条件下,大洋桥石藻的细胞个体变小及比表面积升高。研究结果表明在未来寡营养的大洋上层水体,大气C O2浓度升高会对大洋桥石藻的生理功能产生负面影响,但可能刺激大洋桥石藻的生长。
关键词:海水酸化;C O2加富;营养盐;颗石藻
1 引言
自工业革命以来,化石燃料消耗大增导致了大气C O2浓度的快速升高,而海洋作为一个碳汇,吸收了大气C O2增量的30%~35%[1],这使得目前海水p H值下降了0.1[2—3],相当于海洋中氢离子的浓度上升26%。如果按照目前的C O2排放趋势发展,至21世纪末海水p H可能会下降0.3~0.5[4—5],引发海水酸化问题,对海洋生态环境产生显著影响。
海洋酸化严重影响了海洋生物的钙化过程[2,6—7]。海水碳酸盐系统各参数的浓度与比例的变化会直接或间接地对钙化浮游微藻(颗石藻)的光合作用与钙化作用产生影响,进而可能会改变钙化藻类与非钙化藻类之间的竞争关系,威胁种群的生长,改变群落结构[8—9]。研究表明多数钙化浮游微藻与硅藻及棕囊藻相比,其光合作用速率在目前的大气C O2浓度仍未达到饱和[10—11],表明在大气C O2升高的情况下,钙化藻类的光合作用速率可能对大气C O2浓度的升高产生正反馈过程。此外,也有研究发现大气C O2浓度过高,对颗石藻类的生长与钙化会有抵制作用[10,12],海水碳酸盐系统各参数的浓度与比例的变化会直接或间接地对浮游藻类(尤其是钙化浮游微藻)的钙化作用产生影响[13—14]。目前国内在高浓度C O2对海洋微藻的生理生态影响方面进行了较多的研究[15—19],但针对颗石藻的研究仍然较少。
此外,营养盐水平对海洋浮游植物的生长及生理过程同样都有着重要影响,世界大洋真光层水体营养盐总体表现为氮限制(小于1μmol/L)[20],在氮不足的情况下,颗石藻最终对海洋酸化和C O2浓度升高的趋势产生了哪些反馈?目前,这方面的理解与研究仍然较为薄弱。大洋桥石藻(Gephyrocapsaoceanica)在分类学上属于定鞭金藻门(Haptophyta)颗石藻纲(Coccolithophyceae),是海洋中优势的今生颗石藻。本文选取大洋桥石藻(G.oceanica)作为实验对象,模拟了高氮和低氮的条件以及不同C O2浓度水平下大洋桥石藻的生长情况,探讨了其对大洋桥石藻生长的复合影响,同时着重分析了海水酸化对大洋桥石藻的钙化作用的影响,以期加深了解海洋酸化及营养盐水平对颗石藻及其相关生物地球化学过程的作用。
2 实验设计与分析方法
2.1 实验设计
2.1.1 实验藻种
藻种购自厦门大学近海海洋国家重点实验室海洋藻类保种中心(CC M A),选取颗石藻中较有代表性的种属,大洋桥石藻(G.oceanica),株系CC M BP98。藻类保种时采用的培养液为f/2培养基配方[21],采用的海水取自南大洋贫营养表层海水(盐度约为35),并用0.45μm的醋酸纤维膜过滤,于120℃条件下高压灭菌30 min。
2.1.2 实验装置
实验采用光照培养架(济南凯贝实验仪器有限公司,H T-III),实验设定3个C O2浓度组(380×10-6、800×10-6和1 600×10-6),通过曝气的方式使海水中C O2加富,达到培养实验的酸化条件(图1)。通气装置通过面板式有机玻璃转子流量计(金姚市金泰仪表有限公司,LZ M-4 T)和两通阀控制气体分流与流速的稳定性。
2.1.3 实验条件
在光照培养箱中进行藻细胞的预培养至生长期,取等量生长状况良好的藻种接种至新鲜培养液的玻璃三角瓶中,接种浓度控制在生物量O D665=0.005左右,培养实验条件为25℃、光强7 000 lx、光周期14L∶10 D。各个培养实验组同时进行,历时9 d,实验过程中每天振摇3~4次。实验设置高氮组和低氮组,其中高氮组的硝酸盐初始浓度约为20μmol/L,氮磷比约为12,低氮组的硝酸盐浓度约为6μmol/L,氮磷比约为6,每个C O2浓度组均设3个平行样。
培养实验设定了3个浓度组,即C O2浓度分别为380×10-6、800×10-6和1 600×10-6。其中,380 ×10-6为当前大气中C O2浓度,作为参照组,而800 ×10-6组(实验组1)则以预测的21世纪末、22世纪初大气C O2浓度作为参考浓度,同时设定一个较高浓度的1 600×10-6组(实验组2),模拟远期极端环境条件。

图1 C O2加富培养实验设置(a)及装置示意图(b)Eig.1 The sketch of C O2enrich ment culture experiments(a)and simplified schematic diagram(b)
2.2 分析方法
实验过程中每隔一定时间取样,精确测定培养液的p H值(德国梅特勒实验室p H计,S20P),反映其p H值的细微变化。在营养盐充足和氮相对不足的培养体系中,从第3天开始,3个C O2浓度组的大洋桥石藻培养液的p H值均维持在一个相对稳定的状态,此时培养液中的C O2已达到饱和状态,各组的p H均略有升高,但变化幅度均不超过0.04,表明C O2加富对海水环境的p H值影响基本一致。
同时,取一定体积的藻液,依据《海洋调查规范——海水化学要素观测》[22]进行水样处理与分析,于营养盐自动分析仪(荷兰斯柯拉,san++Skalar)上进行测定,其中,硝酸盐+亚硝酸盐(N O-3+N O-2)、磷酸盐(P O3-4)分别采用镉铜柱还原-重氮偶氮法和磷钼蓝法测定,检测限分别为0.1μmol/L和0.03 μmol/L。
叶绿素a(Chl a)采用萃取荧光法[22]进行过滤与萃取,并于唐纳荧光仪(Turner Designs Eluorometer,M odel 10)上测定,观测其浓度变化。此外,以Lugol碘液试剂固定适量藻液,于生物显微镜(Olympus,C X41)下观察细胞形态变化,同时计数细胞数量,每个样品计数3次,取平均值,计算藻液的细胞密度。
以培养藻液的细胞密度,求出其比生长速率(μ),计算公式(1)如下[23]:
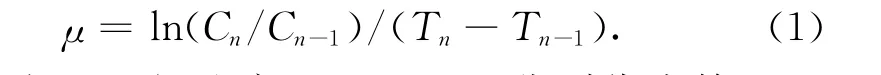
式中,μ为比生长速率,Cn和Cn-1分别代表第n和n-1天的细胞密度度,Tn和Tn-1分别代表培养第n 和n-1天。
实验后期,收集一定量的离心后的藻细胞,用2.5%的戊二醛进行固定,采用离子溅射仪(日本日立公司,IB-3)镀金,之后于高倍扫描电镜(德国里奥电镜有限公司,LE O1530)下进行观察,同时拍摄扫描电镜图片。
取剩余藻液,用玻璃纤维滤膜(W hatman G E/E,孔径0.7μm)进行过滤,富集藻细胞于滤膜上,洗涤烘干称取藻体干质量,之后进行浓酸酸熏24 h处理,除去碳酸盐成分,洗涤烘干得藻体干质量,以此差值计算藻体的无机碳含量(PIC)。同时,将酸处理后的藻细胞,于元素分析仪(德国Elementar,vario EL cube)上进行测定(检出限为0.045 mg),得藻细胞的有机碳含量(P O C)。从而得出细胞的钙化比率占总化学组成的比率,以及在各个培养条件下藻体细胞的钙化作用程度。
3 结果
3.1 营养盐的消耗吸收
在氮充足的培养体系中,随着培养实验的进行,培养液中的硝酸盐和磷酸盐浓度逐渐降低,硝酸盐浓度在第9天相对第1天平均降低4.1μmol/L,磷酸盐则降低了0.54μmol/L(图2a),消耗利用的氮和磷的比值接近于7.6∶1,低于Redfield比值(16∶1)。在整个实验周期中,3个浓度组(380×10-6、800×10-6及1 600×10-6)硝酸盐的消耗百分比分别为33.9%、27.8%及31.8%,磷酸盐的消耗百分比分别为40.1%、39.9%及41.2%。在低氮组的培养体系中硝酸盐浓度在第9天相对第1天平均降低了6.2μmol/L,磷酸盐则降低了0.54μmol/L,3个浓度组的硝酸盐消耗百分比分别为53.4%、69.5%、64.7%,磷酸盐的消耗百分比分别为38.2%、43.8%、37.3%。低氮组消耗利用的氮磷比值平均为11∶1,明显高于氮充足的实验组,显然,在氮相对不足的条件下大洋桥石藻对无机氮具有过度吸收的营养盐策略。
3.2 大洋桥石藻的生长状况
在高氮的培养体系中,大洋桥石藻的细胞数在3种不同的C O2浓度加富条件下均有所增加,平均从0.37×103个/m L增长到第9天的平均4.05×103个/m L。如图3a所示,在培养初期,各组的比生长速率分别为0.25 d-1、0.13 d-1、0.07 d-1。在培养中期(第7天),800×10-6组具有最高的细胞增长,其次是1 600×10-6组,380×10-6组最低(图3a)。Chla含量变化与细胞数相似,培养初期3个浓度组的Chl a含量基本一致并缓慢增加,从第5天开始增长加快,各组之间也表现出了明显的差异,以800×10-6组增长最为迅速,至第8天达到顶峰(3.69μg/L),1 600× 10-6组的增长幅度次之,最高浓度可达2.20μg/L。显然,C O2浓度加富对大洋桥石藻种群增长与Chl a含量有较为显著的促进作用,并且在C O2浓度为800 ×10-6时表现最为明显。
对于低氮的培养体系,大洋桥石藻的细胞数在培养初期增长缓慢,3个浓度组之间并无差异,从第6天开始,不同的C O2浓度加富的影响才表现出差异性,并且随着时间的推移,差异显著增大(图3c)。与参照组相比,低氮培养组800×10-6组和1 600×10-6组的细胞数较为迅速,并且其细胞数明显高于高氮培养组。800×10-6组的最大比生长速率为1.34 d-1,其在第7天达到种群数量最大值(2.17×104cells/m L),为3个浓度组中最高。1 600×10-6组的最大比生长速率为0.85 d-1,其在第8天达到种群数量最大值,1.47×104cells/m L,略高于参照组。Chl a含量变化与细胞数的增长相似(图3d),其含量增长总体水平由低到高依次为:380×10-6组、1 600× 10-6组、800×10-6组。可以看出,在低氮的培养条件下,C O2浓度加富对细胞数的增长与Chl a含量的增加均有促进作用,并且这种促进作用以800×10-6组最为显著,这与氮充足的培养条件相一致,但低氮培养下其种群增长明显强于高氮培养。
3 .3 大洋桥石藻在不同CO2浓度水平的电镜观察结果
扫描电镜观察结果显示低氮培养体系下,参照组(380×10-6)大洋桥石藻细胞形态结构较为完整,其外层的钙壳清晰可见,仅有少数细胞存在着外层钙壳脱落的现象(图4a)。在C O2浓度倍增的条件下,细胞形态结构表现为形态不完整,外层钙壳脱落,在所观察的样本上随处可见散落的钙壳(图4b),表明培养液C O2浓度倍增,p H降低对颗石藻钙化作用具有显著的负影响。对于C O2浓度为1 600×10-6的实验组,大部分细胞因钙壳脱落,表面破碎,出现明显的缺失现象,细胞形态不规则。不同C O2浓度加富培养下的细胞个体大小有所差异,由大到小依次为380× 10-6组、800×10-6组、参照组(380×10-6),表明低氮条件下海水酸化可能引起大洋桥石藻的细胞体积的降低和比表面积的升高。

图2 不同C O2浓度加富条件下(380×10-6、800×10-6、1 600×10-6),培养液中硝酸盐(N O-3)和磷酸盐(P O3-4)浓度随时间的变化Eig.2 The time series change of nitrate concentration and phosphate concentration in the C O2enrich ment(380×10-6;800×10-6and 1 600×10-6)culture experiments a.高氮组硝酸盐浓度的变化(μmol/L);b.高氮组磷酸盐浓度的变化(μmol/L);c.低氮组硝酸盐浓度的变化(μmol/L);d.低氮组磷酸盐浓度的变化(μmol/L)a.Nitrate concentration(μmol/L)under sufficient nutrients;b.phosphate concentration(μmol/L)under sufficient nutrients;c.nitrate concentration(μmol/L)under nitrate deficiency;d.phosphate concentration(μmol/L)under nitrate deficiency
3.4 细胞有机碳与无机碳合成
表1所示为氮充足与氮不足两种条件下,大洋桥石藻经不同C O2浓度加富培养后,其细胞中的有机碳与无机碳含量,及各自占细胞总干质量比。结果显示,800×10-6组的有机碳合成比重远低于其他2个浓度组,但其无机碳合成比重却是3个浓度组中最高的。可见,当C O2浓度加倍(800×10-6)时,光合作用合成有机碳量减少,但钙化作用合成无机碳量却在增加。而当C O2浓度达到1 600×10-6时,光合作用过程略有增强,钙化作用反而有所下降。表明大洋桥石藻对C O2加富的响应呈现相反的趋势。就细胞的无机碳与有机碳比而言,3个浓度组分别为77.7∶1、329.4∶1、66.7∶1,进一步体现了800×10-6组显著的钙化作用。
在氮相对不足条件下,大洋桥石藻的有机碳与无机碳合成量也受到不同C O2浓度水平的影响。800 ×10-6组的无机碳含量占细胞总干质量比在3个浓度组中最高,而有机碳含量占细胞总干质量比仅仅略高于参照组。1 600×10-6组的有机碳与无机碳的合成比重均高于参照组,但与800×10-6组相比,则有机碳比重较高而无机碳比重较低。可见,在氮不足条件下,当C O2浓度加倍(800×10-6)时,大洋桥石藻的有机碳与无机碳合成量均有所增加,其中无机碳的增加更为显著。就细胞的无机碳与有机碳比而言,3个浓度组分别为26.6∶1、27.5∶1、24.2∶1,800×10-6组的光合作用与钙化作用同步增强,三者的比值没有显著差异。
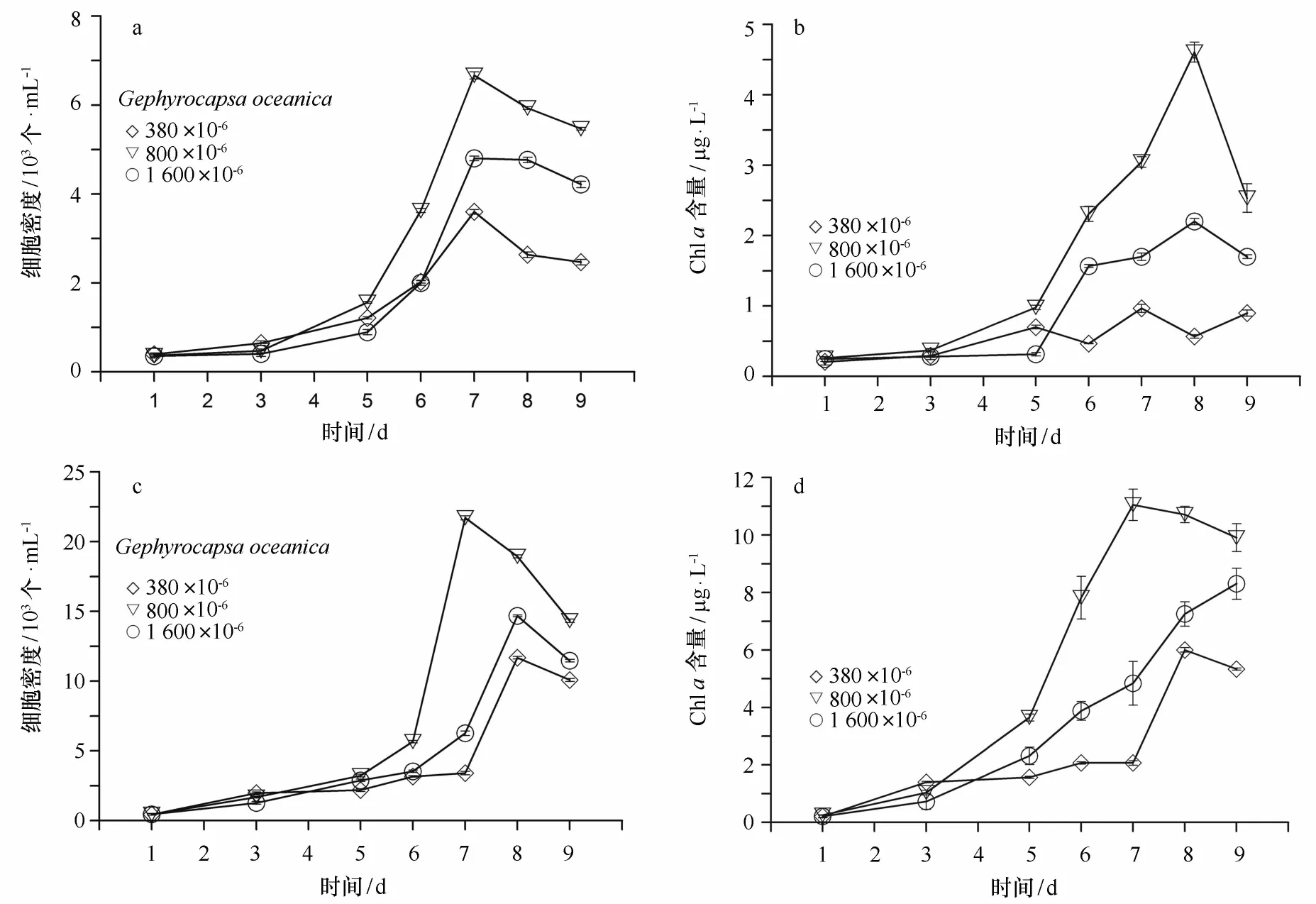
图3 不同C O2浓度加富条件下(380×10-6、800×10-6及1 600×10-6),细胞数和Chl a含量随时间的变化Eig.3 The time series change of cell density and chlorophyll a in the C O2enrich ment(380×10-6,800×10-6and1 600×10-6)culture experiments a.高氮组细胞数的变化(103cells/m L);b.高氮组Chl a含量的变化(μg/L);c.低氮组细胞数的变化(103cells/m L);d.低氮组Chl a浓度的变化(μg/L)a.The cell nu mber(103cells/m L)under sufficient nutrients;b.chlorophyll a(μg/L)under sufficient nutrients;c.the cell nu mber (103cells/m L)under nitrate deficiency;d.chlorophyll a(μg/L)under nitrate deficiency

表1 高氮组和低氮组中细胞中有机碳及无机碳含量及比例Tab .1 The concentration and proportion of organic carbon and inorganic carbon under condition of sufficient nutrients and nitrate limitation

图4 低氮培养下大洋桥石藻细胞的典型扫描电镜照片Eig.4 The typical scanning electron micrograph of G.oceanic cell under nitrate deficiency a.C O2浓度为380×10-6的培养条件下,比例尺为5μm;b.C O2浓度为800×10-6的培养条件下,比例尺为10μm;c.C O2浓度为1 600×10-6的培养条件下,比例尺为5μm a.The set concentration of C O2is 380×10-6,the scaleis 5μm;b.C O2is 800×10-6,the scale is 10μm;c.C O2is 1 600×10-6,the scale is 5 μm
4 讨论
4.1 海水酸化与营养盐不足的复合响应
为了模拟大洋桥石藻对营养盐相对不足和海水酸化的复合响应,设置了高氮组和低氮组两种不同的营养盐状况以及3组不同的C O2浓度加富(380× 10-6、800×10-6及1 600×10-6),其中低氮组硝酸盐的初始浓度约为6μmol/L,高氮组的硝酸盐初始浓度约为20μmol/L。对大洋桥石藻在3组C O2浓度加富下的Chl a生物量(最高Chl a浓度)进行基于IB M SPSS Statistics 20.0的单因素方差分析及多重比较(显著性水平为0.05),结果显示Chl a生物量在3个浓度组间有统计学差异(P值为0.01),表明C O2加富对大洋桥石藻的生长起到了促进作用;多重比较分析结果则显示380×10-6与800×10-6组有显著差异(P值为0.01),但与1 600×10-6组差异不显著(P值为0.06),主要由于C O2浓度1 600×10-6已达到抑制藻类的阈值,显然800×10-6组最有利于大洋桥石藻的生长繁殖,钙化藻类大洋桥石藻的光合作用在当前大气C O2浓度仍未达到饱和,与前人的研究一致[10]。
此外,进一步对大洋桥石藻在高氮和低氮环境下的Chl a生物量(最高Chl a浓度)及种群增长(最大细胞密度)进行单因素分析(显著性水平为0.05),结果显示藻类生物量及种群增长在不同营养盐状态下差异十分显著(P值均为0.01)。如图5所示,营养盐充足条件下,不同C O2加富组的细胞数普遍不高。而在氮不足和低氮磷比的条件下,大洋桥石藻的细胞数很高,与营养盐充足时相比,高了1个数量级,并且由高到低依次为800×10-6组、1 600×10-6组、380 ×10-6组。大洋桥石藻对氮不足和C O2加富表现出相反的响应机制。对比两种营养盐状态,氮不足下的大洋桥石藻各项生理指标进一步提高,显然,作为培养液的唯一变量,营养盐浓度和营养盐结构起到了重要的作用,低氮磷比可能促进了大洋桥石藻的生长繁殖,同时细胞对营养盐的利用效率增强,其与C O2加富相结合则使这种作用进一步增强。
4.2 氮不足和酸化耦合下颗石藻的生理生态响应
海水中的DIC浓度较高,但大洋桥石藻的光合固碳却远远没有饱和[24—27],自然条件下的DIC条件对颗石藻细胞的生长并不会起到限制作用,因此,C O2或DIC的增加通常并不能促进其生长[28—30]。在营养盐方面,研究表明光合作用与氮代谢是两个紧密相连的过程[31]。Nimer等[32]的研究则表明,在营养盐限制时,p H降低(pC O2700×10-6)不影响颗石藻的生长。在氮限制培养时发现[31],氮限制可诱导小球藻的C O2浓缩机制(CC M)形成,同时提高微藻细胞的氮利用率,使细胞的C O2固定速率维持在合理水平,以减少对氮的需求。而在本文实验中,无论是在氮充足或氮不足条件下,当C O2浓度增加时,大洋桥石藻的有机碳含量均略有增加,细胞的光合固碳作用有所增强,并且在氮不足条件下大洋桥石藻细胞的有机碳比重均远大于氮充足条件,暗示在海水酸化下,寡营养盐条件可能会促进细胞内有机碳含量的增加。

图5 氮充足(蓝色)与氮不足(红色)以及不同C O2加富浓度(380×10-6、800×10-6、1 600×10-6)条件下,培养过程的细胞数最高值的对比Eig.5 The comparison of cell density maxima in different C O2enrich ment culture experiments(380×10-6,800× 10-6,1 600×10-6)under condition of sufficient nutrients(blue)and nitrate limitation(red)
电镜观察结果显示,海洋酸化严重削弱了大洋桥石藻的钙化作用。从PIC/P O C值的变化结果来看,C O2浓度加富下均呈降低趋势,不管是氮不足还是氮充足条件。在野外的围隔实验研究中,实验条件的差异以及颗石藻种类的不同可能使研究结果有所不同,随着p H值的降低,PIC与P O C的变化总结起来基本为以下3种情况:(1)PIC减少,PIC/P O C值降低;(2)P O C增加,PIC/P O C值降低;(3)PIC、P O C均减少,PIC/P O C值几乎不变[33—35]。可见,p H的降低,PIC/P O C值也随之降低,这在室内培养实验与野外围隔实验中是基本一致,而不同的实验条件对PIC与P O C的变化影响则略有不同。
5 结论
在氮充足和氮不足的条件下,C O2加富均能一定程度上促进大洋桥石藻种群增长(细胞数和比生产速率)和Chl a含量的提高,并且这个促进作用在C O2浓度为800×10-6时表现最为明显。对比两种营养盐状态,氮相对不足的营养盐状态下的大洋桥石藻各项生理指标均有显著提高,显然,作为培养液的唯一变量,营养盐浓度和营养盐结构起到了重要的作用,低氮磷比可能促进了大洋桥石藻的生长繁殖。电镜观测结果表明海水酸化对钙化作用具有显著的负影响,此外PIC/P O C值在C O2浓度加富下呈一定的降低趋势。海水酸化会使大洋桥石藻表面的颗石粒发生畸变,或者缺角不完整颗石层的厚度变薄,以及使得细胞个体的变小和比表面积的升高。大洋桥石藻对海水酸化及氮相对不足的复合响应,暗示在大洋寡营养盐环境下,未来大气C O2浓度的升高一定程度上可能刺激大洋桥石藻的生长。
致谢:本所江志兵博士协助进行SPSS方差统计分析,两位审稿专家提出了严谨细致的意见,在此一并致谢。
参考文献:
[1] Quay P D,Tibrook B,W ong C S.Oceanic uptake of fossilfuel C O2:carbon13 evidence[J].Science,1992,256(5053):74-79.
[2] Orr J C,Eabry V J,Au mont O,et al.Anthropogenic ocean acidification over the twenty-first century and itsimpact on calcifying organisms[J].Nature,2005,437(7059):681-686.
[3] Doney S C,Eabry V J,Eeely R A,et al.Ocean Acidification:the other C O2problem[J].Annu Rev M ar Sci,2009,1:169-192.
[4] Caldeira K,Wickett M E.Oceanography:anthropogenic carbon and ocean p H[J].Nature,2003,425(6956):365,doi:10.1038/425365a.
[5] Raven J A.Ocean acidification due to increasing atmospheric carbon dioxide[M].London,U K:Royal Society,2005.
[6] Eeely R A,Sabine C L,Lee K,et al.Impact of anthropogenic C O2on the CaC O3system in the oceans[J].Science,2004,305(5682):362-366.
[7] Zondervan I,Zeebe R E,Rost B,et al.Decreasing marine biogenic calcification:a negative feedback on rising atmospheric pC O2[J].Global Biogeochemical Cycles,2001,15(2):507-516.
[8] Rochelle-Newall E,Delille B,Erankignoulle M,et al.Chromophoric dissolved organic matterin experimental mesocosms maintained under different pC O2levels [J].M arine Ecology Progress Series,2004,272:25-31.
[9] 阮祚禧.海洋酸化对钙化浮游植物颗石藻的影响[D].汕头:汕头大学,2008.Ruan Zuoxi.Effects of ocean acidification on the calcifying phytoplankton coccolithophore[D].Shantou:Shantou U niversity,2008.
[10] Riebesell U,Zondervan I,Rost B,et al.Reduced calcification of marine plankton in response to increased atmospheric C O2[J].Nature,2000,407 (6802):364-367.
[11] Rost B,Riebesell U,Burkhardt S,et al.Carbon acquisition of bloom-forming marine phytoplankton[J].Limnol Oceanogr,2003,48(1):55-67.
[12] 蔡小霞,潘建明,张海娜,等.C O2加富对颗石藻Emilianiahuxleyi生长及钙化作用的影响[J].海洋环境科学,2013,32(2):249-253.Cai Xiaoxia,Pan Jian ming,Zhang Haina,et al.Effects of elevated C O2concentration on growth and calcification of the coccolithes Emilianiahuxleyi[J].M arine Environ mental Science,2013,32(2):249-253.
[13] Raven J A,Crawfurd K.Environ mental controls on coccolithophore calcification[J].M arine Ecology Progress Series,2012,470:137-166.
[14] Eeng Y,W arner M E,Zhang Y,et al.Interactive effects of increased pC O2,temperature and irradiance on the marine coccolithophore Emiliania huxleyi(Prymnesiophyceae)[J].European Journal of Phycology,2008,43(1):87-98.
[15] 夏建荣,高坤山.高浓度C O2对极大螺旋藻生长和光合作用的影响[J].水生生物学报,2001,25(5):474-480.Xia Jianrong,Gao Kunshan.Effects of high C O2concentration on growth and photosynthesis of Spirulina maxima[J].Acta H ydrobiologica Sinica,2001,25(5):474-480.
[16] 夏建荣,高坤山.C O2浓度升高对斜生栅藻生长和光合作用的影响[J].植物生理学通讯,2002,38(5):431-433.Xia Jianrong,Gao Kunshan.Effect of elevated C O2concentration on growth and photosynthesis of Scenedesmusobliquus[J].Plant Physiology Com munications,2002,38(5):431-433.
[17] 夏建荣.大气C O2浓度升高对海洋浮游植物影响的研究进展[J].湛江海洋大学学报,2006,26(3):106-110.Xia Jianrong.Advancesin the study ofthe effects of elevaed atmospheric C O2concentrations on marine phytoplankton[J].Journal of Zhanjiang O-cean U niversity,2006,26(3):106-110.
[18] 邹定辉,高坤山.高C O2浓度对大型海藻光合作用及有关过程的影响[J].生态学报,2002,22(10):1750-1757.Zou Dinghui,Gao Kunshan.Effects of elevated C O2concentration on the photosynthesis and related physiological processes in marine macroalgae [J].Acta Ecologica Sinica,2002,22(10):1750-1757.
[19] 徐智广,邹定辉,张鑫,等.C O2和硝氮加富对龙须菜(Gracilarialemaneiformis)生长、生化组分和营养盐吸收的影响[J].生态学报,2008,28 (8):3752-3759.Xu Zhiguang,Zou Dinghui,Zhang Xin,et al.Effects ofincreased atmospheric C O2and N supply on growth,biochemicalcompositions and uptake of nutrients in Gracilarialemaneiformis(Rhodophyta)[J].Acta Ecologica Sinica,2008,28(8):3752-3759.
[20] Ealkowski P G.Evolution ofthe nitrogen cycle and itsinfluence on the biological sequestration of C O2in the ocean[J].Nature,1997,387(3872):272-275.
[21] 程兆第,高亚辉,刘师成.福建沿岸微型硅藻[M].北京:海洋出版社,1993:1-125.Cheng Zhaodi,Gao Yahui,Liu Shicheng.Nano-planktonic diatoms in Eujian coast[M].Beijing:China Ocean Press,1993:1-125.
[22] 国家海洋标准计量中心,国家海洋局第一海洋研究所.海洋调查规范G B/T 12763-2007[S].北京:中国标准出版社,2008.National Technical Supervision Bureau,Eirst Institute of Oceanography,State Oceanic Administration.Specificationsfor oceanographic survey G B/T 12763-2007[S].Beijing:Chinese Standard Press,2008.
[23] 陈雄文,高坤山.C O2浓度对中肋骨条藻的光合无机碳吸收和胞外碳酸酐酶活性的影响[J].科学通报,2003,48(21):2275-2279.Chen Xiongwen,Gao Kunshan.Effects of C O2concentration on the photosynthetic inorganic carbon uptake and extracellular carbonic anhydrase ctiveity of Skeletonemacostatum[J].Chinese Science Bulletin,2003,48(21):2275-2279.
[24] Paasche E.A tracer study oftheinorganic carbon uptake during coccolith formation and photosynthesisin the coccolithophorid Coccolithushuxleyi [J].Physiol Plantaru m,1964,3(suppl.):1-82.
[25] Nielsen M V.Photosynthetic characteristics of the coccolithophorid Emiliania huxleyi(Prymnesiophyceae)exposed to elevated concentrations of dissolved inorganic carbon[J].J Phycol,1995,31(5):715-719.
[26] Berry L,Taylor A R,Lucken U,et al.Calcification and inorganic carbon acquisition in coccolithophores[J].Eunct Plant Biol,2002,29(2/3):289 -299.
[27] Zondervan I,Rost B,Riebesell U.Effect of C O2concentration on the PIC/P O C ratio in the coccolithophore Emilianiahuxleyi grown underlightlimiting conditions and different daylengths[J].J Exp M ar Biol Ecol,2002,272(1):55-70.
[28] Paasche E,Brubak S,SkattebØl S,et al.Growth and calcification in the coccolithophorid Emiliania huxleyi(Haptophyceae)at low salinities[J].Phycologia,1996,35(5):394-403.
[29] Buitenhuis E T,De Baar H J W,Veldhuis M J W.Photosynthesis and calcification by Emilianiahuxleyi(Prymnesiophyceae)as a function ofinorganic carbon species[J].J Phycol,1999,35(5):949-959.
[30] Clark D R,Elynn K J.The relationship between the dissolved inorganic carbon concentration and growth ratein marine phytoplankton[J].Proc R Soc Lond B,2000,267:953-959.
[31] Turpin D H,W eger H G.Steady-state chlorophyll a fluorescence transients during am moniu m assimilation by the N-limited green alga Selenastrum minutum[J].Plant Physiol,1988,88(1):97-101.
[32] Nimer N A,Brownlee C,M errett M J.Carbon dioxide availability,intracellular p H and growth rate of the coccolithophore Emilianiahuxleyi[J].M ar Ecol Prog Ser,1994,109:257-262.
[33] Beardall J,Griffiths H,Raven J A.Carbon isotope discrimination and the C O2accu mulating mechanism in Chlorellaemersonii[J].J Exp Bot,1982,33(4):729-737.
[34] Delille B,Harlay J,Zondervan I,et al.Response of primary production and calcification to changes of pC O2during experimental blooms ofthe coccolithophorid Emilianiahuxleyi[J].Global Biogeochem Cy,2005,19(2):G B2023.
[35] Engel A,Zondervan I,Aerts K,et al.Testing the direct effect of C O2concentration on a bloom ofthe coccolithophorid Emilianiahuxleyiin mesocosm experiments[J].Limnol Oceanogr,2005,50(2):493-507.
Combined effects of seawater acidification and nitrogen deficiency on Gephyrocapsaoceanica
Cai Xiaoxia1,Yu Peisong1,Zhang Haifeng1,H u Ji1,Zhang Haina1,Pan Jian ming1
(1.Laboratoryof Marine Ecosystem and Biogeochemistry,Second Institute of Oceanography,State Oceanic Administration,Hangzhou 310012,China)
Abstract:Combined influences of nutrients status and seawater C O2concentration on microalgae growth and physiology of Gephyrocapsaoceanic was investigated.It was shown that the abundance of G.oceanic significantly increased during C O2enrich ment experiments at different nutrient levels(nitrogen-deficiency vs.nutrient-rich).H owever,the strongestresponse was observed in cell density and chlorophylla concentration of G.oceanic when nitrogen was deficiency,suggesting low nutrients concentration and low N/P ratio might be favor to the cell growth.Besides,electron microscopy resultsindicated that seawater acidification had significant negativeimpact on the calcification and cell size distribution(became smaller)of G.oceanic when nitrogen was deficiency.H owever,in future oligotrophic open ocean,increasing C O2concentration might stimulate G.oceanic thrive.
Key words:seawater acidification;C O2enrich ment;nutrients;Coccolithophores
*通信作者:潘建明,男,研究员,长期从事海洋环境化学和生物地球化学研究。E-mail:jmpan@sio.org.cn
作者简介:蔡小霞(1987—),女,福建省漳州市人,助理研究员,从事近海海洋环境化学研究。E-mail:caixx1987@163.com
基金项目:国家海洋局第二海洋研究所基本科研业务费专项资助(JG1406);国家海洋局全球变化与海气相互作用专项(G ASI-03-01-03-02);浙江省自然科学基金(L Y14 D060008)。
收稿日期:2015-07-21;
修订日期:2016-01-09。
中图分类号:Q948.8
文献标志码:A
文章编号:0253-4193(2016)04-0130-09
