吐温-20和吐温-80对稻瘟病菌孢子萌发的影响
2016-03-21朱紫薇许有嫔李沛华陈学伟李伟滔
冉 莉,朱紫薇,杨 超,许有嫔,李沛华,贺 闽,王 静,陈学伟,李伟滔
(四川农业大学水稻研究所,四川 成都 611130)
吐温-20和吐温-80对稻瘟病菌孢子萌发的影响
冉 莉,朱紫薇*,杨 超,许有嫔,李沛华,贺 闽,王 静,陈学伟,李伟滔**
(四川农业大学水稻研究所,四川 成都 611130)
为了研究吐温-20和吐温-80对稻瘟病菌孢子萌发的影响,从而明确稻瘟病菌喷雾接菌时孢悬液的配置方法和保存时长。本试验通过配置不同浓度的吐温-20和吐温-80的孢悬液,将其放置不同的时间,然后通过显微镜观察吐温-20和吐温-80对体型不同的稻瘟病菌孢子萌发的影响。结果发现稻瘟病菌孢子在水、吐温-20和吐温-80的溶液中均可萌发,且在吐温-20和吐温-80的溶液可以生成两条或多条芽管。吐温-20和吐温-80均能抑制稻瘟病菌孢子的萌发,但与吐温-80相比,吐温-20对孢子萌发的抑制作用较小。与孢子体型较大的稻瘟病菌Zhong10-8-14相比,体型较小的Guy11的孢子对吐温-20和吐温-80的抑制作用更敏感。由此表明,吐温-20更适合用来制作稻瘟病菌的孢悬液。新鲜配置的孢悬液,只要吐温-20的浓度不高于0.50 %,且放置时间不超过2 h,孢子的萌发率不会显著降低,依然可以用来进行喷雾接菌试验。
稻瘟病菌;萌发;吐温-20;吐温-80
稻瘟病是由子囊菌(Magnaportheoryzae)引发的水稻重大病害之一[1]。它能够侵染从幼苗期到成熟期各个时期的水稻植株,从而引发苗瘟、叶瘟、茎瘟和穗瘟等,严重影响水稻产量和品质[2-3]。而水稻是世界重要的粮食作物,因此,为了保障粮食安全,水稻稻瘟病的研究一直以来被病理学家和育种家所重视。
水稻稻瘟病菌致病性的症状鉴定与分析是研究稻瘟病的基础[4],而稻瘟病菌孢子的萌发是稻瘟病菌致病的前提。在实验条件下,喷洒在水稻叶片上的孢子均匀度和孢子萌发率严重影响了分子生物学实验的精确度[5-6]。因此,为了使孢子均匀分布在孢悬液中,并在喷洒稻瘟病菌孢子后能够使其很好的粘附于叶片上。例如在制作白僵菌(Beauveriabassiana)和绿僵菌(Metarhiziumanisopliae)孢子的孢悬液时通常添加吐温-80,当吐温-80的浓度高于0.10 %时,白僵菌孢子的萌发率显著降低[9]。而当高于0.20 %时,绿僵菌孢子的萌发率才显著降低[7]。在稻瘟病菌的接菌试验中,通常加入吐温-20和吐温-80等表面活性剂[4,7-10]。但是,目前尚未有报道吐温-20和吐温-80等表面活性剂是否影响稻瘟病菌孢子的萌发率。此外,在制作稻瘟病菌孢悬液后,通常情况下会及时喷洒在水稻叶片上,以防止稻瘟病菌孢悬液放久后影响接菌效果。但是,配置的新鲜的孢悬液具体能够存放多久,也未有相关的报道。
本研究中使用吐温-20和吐温-80两种试剂,研究它们对稻瘟病菌孢子萌发的影响。由于明胶的熔点较低,因此,本研究并未使用明胶进行相关研究。通过本研究,意在通过试验明确吐温-20和吐温-80是否影响稻瘟病菌孢子的萌发,并比对两者中哪种试剂更适宜配置孢悬液。同时,明确新鲜孢悬液的保存时间,为稻瘟病菌的喷菌试验提供可靠的技术指导。
1 材料与方法
1.1 稻瘟病菌生理小种
10个稻瘟病菌生理小种:B15、B13、Guy11、A25、HN02、HN01、FJ10、ZB25、HN41和Zhong10-8-14,均由四川农业大学水稻研究所保存。
1.2 孢悬液的制备与显微观察
稻瘟病菌生理小种使用滤纸片保存法保存在-20 ℃冰箱中,使用前剪取一小片滤纸,放置在配置好的完全培养基上(Complete Medium,CM),然后放置在培养箱中,温度调至26 ℃,光12 h/暗12 h。待生长12~14 d后,用灭菌水将孢子从培养皿上洗下来,过滤后放于光学显微镜下镜检,用血球计数板计数,将孢子浓度调至1×105/mL。之后根据试验要求,分别制备终浓度为0.00 %、0.05 %、0.10 %、0.20 %和0.50 %的吐温-20和吐温-80的孢悬液。将这些不同浓度的孢悬液放置在26 ℃培养箱中。放置的时间分别为0 min、10 min、30 min、1 h、2 h和5 h。之后,用移液枪吸取5 μl不同浓度和放置不同时间的孢悬液,滴在基本培养基上(Minimal Medium,MM)。将其放置4 h后,利用Zeiss光学显微镜观察孢子的萌发情况。试验重复4次。
1.3 培养基的配方
CM:D-Glucose(10 g/L)、Peptone(2 g/L)、Yeast Extract(1 g/L)、Casamino Acids(1 g/L)、NaNO3(6 g/L)、KCl(0.5 g/L)、MgSO4(0.5 g/L)、KH2PO4(1.5 g/L)、Vitamin(0.1 %)、Agar(15 g/L)和微量元素(0.1 %)。将pH值调至 6.5。
MM:Dextrose(10 g/L)、NaNO3(6 g/L)、KCl(0.5 g/L)、MgSO4(0.5 g/L)、KH2PO4(1.5 g/L)、Biotin(0.00025 %)、Thiamine(0.1 %)、Agar(15 g/L)和微量元素(0.1 %)。将pH值调至 6.5。
1.4 数据分析
利用Excel 2010和SPSS 13.0分析孢子的萌发率。萌发率( %)=视野下萌发的孢子数量/视野下总的孢子数量×100 %,试验重复4次。
2 结果与分析
2.1 稻瘟病菌孢子在水、吐温-20和吐温-80溶液中的萌发
Guy11通常作为研究稻瘟病菌致病性的一个生理小种[5,11-12]。因此,用Guy11作为研究对象,研究稻瘟病菌孢子是否能够在水、吐温-20和吐温-80溶液中萌发。将Guy11的孢子配置的孢悬液放置30 h后,置于显微镜下观察,发现孢子在水、吐温-20和吐温-80的孢悬液(0.2 %)中均能够萌发,但是并没有形成附着胞(图1A)。这说明稻瘟病菌孢子附着胞的形成是需要一定的接触载体。此外,我们发现,稻瘟病菌孢子在水中萌发后产生一条芽管,而在吐温-20孢悬液中能够产生两条芽管,在吐温-80孢悬液中甚至能够产生多余两条的芽管(图1A)。由此表明,吐温-20和吐温-80能够刺激稻瘟病菌孢子产生更多的芽管。
2.2 筛选大小不同的稻瘟病菌孢子
不同稻瘟病菌生理小种的孢子的大小也不尽相同,吐温试剂对它们孢子萌发的影响也可能不同。随机选用了10个稻瘟病菌生理小种,它们孢子的长度从26.3~46.3 μm不等(图1B和 1C)。这些大小不同的孢子对吐温-20和吐温-80的反应是否也不同呢?鉴于此,笔者筛选孢子长度较长的Zhong10-8-14和孢子长度较短的Guy11用作后续试验的生理小种。
2.3 吐温-20和吐温-80对孢子萌发率的影响
孢子生长出芽管为萌发(图1D)。以孢子较小的Guy11为研究对象,在配置的不同浓度的吐温-20孢悬液中,不同浓度的孢悬液放置时间为0 min、10 min、30 min、1 h和2 h时,Guy11孢子萌发率均约为95 %。放置时间为5 h时,萌发率下降约至80 %(图1E)。在配置的不同浓度的吐温-80孢悬液中,当吐温-80的浓度低于0.20 %时,放置时间为0 min、10 min、30 min和1 h时,Guy11的萌发率均约为 90 % ,当吐温-80的浓度为0.20 %和0.50 %时,放置时间为2和5 h时,萌发率下降至约70%(图1F)。结果表明,吐温-20和吐温-80均能够抑制Guy11的孢子萌发,吐温-80强于吐温-20的抑制作用。
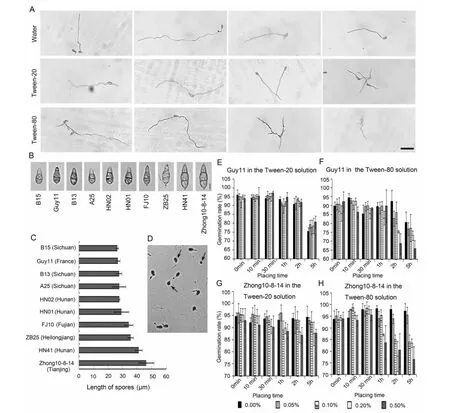
A:Guy11孢子在水和表面活性剂中30 h时的萌发,标尺是50 μm。B:稻瘟病菌孢子的显微照片,标尺是 10 μm 。C:稻瘟病菌孢子长度的柱形图,n=6。D:Guy11孢子萌发的显微照片。箭头显示未萌发的孢子。E:Guy11的孢子在吐温-20孢悬液中的萌发率。F:Guy11的孢子在吐温-80孢悬液中的萌发率。G:Zhong10-8-14的孢子在吐温-20孢悬液中的萌发率。H:Zhong10-8-14的孢子在吐温-80孢悬液中的萌发率,n=4A, the germination of Guy11 in water and surfactants when the placing time is 30h. Bar is 50 μm. B, the micrographs of M. oryzae spore. Bar is 10 μm. C, the chart of the lengths of M. oryzae spores. n=6. D, the germination of Guy11 spores observed using microcopy. Arrows show that the spores do not germinate. The germination rates of Guy11 in the Tween-20 (E) and Tween-80 (F) at the different concentrations and placing times, n=4. The germination rates of Zhong10-8-14 in the Tween-20 (G) and Tween-80 (H) at the different concentrations and placing times, n=4图1 吐温-20和吐温-80对稻瘟病菌孢子萌发和萌发率的影响Fig.1 Germination and germination rate of M. oryzae spores in the Tween-20 and Tween-80 solutions
以孢子较大的Zhong10-8-14为研究对象,配置的吐温-20的孢悬液无论放置哪个时间段,在0.00 %、0.05 %和0.10 %的浓度下,孢子萌发率均约为 95 %,当浓度提升到0.20 %和0.50 %时,孢子的萌发率降低至约90 %(图1G)。当配置成不同浓度吐温-80的孢悬液后,放置时间在1 h内,浓度为0.00 %、0.05 %和0.10 %时,孢子萌发率均约为95 %,而当放置时间为2和5 h时,浓度为0.20 %和0.50 %的萌发率降至约80 %(图1H)。结果表明吐温-20和吐温-80均能够抑制Zhong10-8-14孢子的萌发,而吐温-80强于吐温-20的抑制作用。
通过比较吐温-20和吐温-80对稻瘟病菌孢子萌发率的影响,发现体型较小的Guy11对吐温-20和吐温-80更敏感。与吐温-80相比,吐温-20更适合制作孢悬液。新鲜配置的吐温-20的孢悬液,当吐温-20的浓度小于0.50 %时,较大或较小的稻瘟病菌孢子均能够在26 ℃存放2 h而萌发率不会显著降低。
3 讨论与结论
本研究较为系统的研究了表面活性剂吐温-20和吐温-80对稻瘟病菌孢子萌发的影响。稻瘟病菌孢子能够在水或者吐温溶液中萌发,这与 黑曲霉(Aspergillusniger)不能在水中萌发[13-14]完全不同。尽管稻瘟病菌与黑曲霉均为病原真菌,且都属于半知菌亚门,但是其孢子的萌发机制存在一定的差异。这可能与其各自的不同侵染机制有关。此外,试验还发现吐温-20和吐温-80能够刺激稻瘟病菌孢子产生多条芽管,这增加了稻瘟病菌孢子选择更多的侵染位点,对稻瘟病菌孢子成功入侵宿主提供了更好的保障,因此,在制作稻瘟病菌孢悬液的过程中,加入吐温试剂,不仅有利于孢子均匀分布和更好的附着在叶片表面,而且还有可能增加稻瘟病菌孢子的成功侵染水稻的机率。
前期研究发现稻瘟病菌孢子能够在玻璃纸上萌发且形成附着胞[12]。本研究发现稻瘟病菌孢子在没有固体介质时,并没有发现附着胞的形成。由两方面结果推断,稻瘟病菌附着胞的形成需要固体介质。
伯格曼法则中哺乳动物和鸟类的体型大小明显受到环境的影响[15-16]。本试验中,体型较小的孢子对吐温-20和吐温-80抑制孢子萌发的作用更敏感,这与动物中的伯格曼法则相似。由此表明稻瘟病菌和其它真菌也可能存在伯格曼法则。
稻瘟病菌孢子能够在水、吐温-20和吐温-80的溶液中萌发,但不形成附着胞。吐温-20和吐温-80能够诱导稻瘟病菌孢子产生更多的芽管。体型较小的孢子对吐温-20和吐温-80抑制孢子萌发的作用更敏感。与吐温-80相比,吐温-20更适合用来制作稻瘟病菌的孢悬液。新鲜制作的孢悬液,当吐温-20的浓度不超过0.50 %时,能够存放2 h,而孢子萌发率不会显著下降。这为稻瘟病菌的喷菌试验提供了明确的指导。
[1]Dean R, Vankan J L, Pretorius ZA, et al. The top 10 fungal pathogens in molecular plant pathology[J]. Molecular Plant Pathology, 2012, 13:414-430.
[2]Li W T, He M, Wang J, et al. Zinc finger protein (ZFP) in plants-a review[J]. Plant Omics Journal, 2013(6):474-480.
[3]Fernandez J, Wilson R A. Cells in cells:morphogenetic and metabolic strategies conditioning rice infection by the blast fungusMagnaportheoryzae[J]. Protoplasma, 2014, 251:37-47.
[4]Jia Y, Valent B, Lee F N. Determination of host responses toMagnaporthegriseaon detached rice leaves using a spot inoculation method[J]. Plant Disease, 2003, 87:129-133.
[5]Dagdas Y F, Yoshino K, Dagdas G, et al. Septin-mediated plant cell invasion by the rice blast fungus,Magnaportheoryzae[J]. Science, 2012, 336:1590-1595.
[6]Schena M, Shalon D, Davis R W, et al. Quantitative monitoring of gene expression patterns with a complementary DNA microarray[J]. Science, 1995, 270:467-470.
[7]史 婵, 陈薇兰, 王 静, 等. 三份重要水稻资源的稻瘟病抗性鉴定及不同评价指标间的相关性分析[J]. 植物保护学报, 2014, 41(4):390-395.
[8]李增智, 宣 言, 刘玉珍. 几种表明活性剂及营养物对虫生真菌孢子萌发的影响[J]. 安徽农业大学学报, 1996, 23(3):355-359.
[9]Li W T, Chen W L, Yang C, et al. Identification and network construction of zinc finger protein (ZFP) genes involved in the rice-Magnaportheoryzaeinteraction[J]. Plant Omics Journal, 2014(7):540-548.
[10]任巧云, 殷 宏, 关贵全, 等. 不同浓度吐温-80与白僵菌分生孢子的相容性测定[J]. 安徽农业科学, 2009, 37(18):8555-8687.
[11]Kankanala P, Czymmek K, Valent B. Roles for rice membrane dynamics and plasmodesmata during biotrophic invasion by the blast fungus[J]. The Plant Cell, 2007, 19:706-724.
[12]Mentlak T A, Kombrink A, Shinya T, et al. Effector-mediated suppression of chitin-triggered immunity byMagnaportheoryzaeis necessary for rice blast disease[J]. The Plant Cell, 2012, 24:322-335.
[13]Ynagita T. Biochemical aspects of the germination of conidia spores ofAspergillusniger[J]. Archives of Microbiology, 1957, 26:329-344.
[14]Trinci A P J. Exponential growth of the germ tubes of fungal spores[J]. Journal of General Microbiology, 1971, 67:345-348.
[15]Meiri S, Dayan T. On the validity of Bergmann's rule[J]. Journal of Biogeography, 2003(30):331-351.
[16]Millien V, Kathleen Lyons S, Olson L, et al. Ecotypic variation in the context of global climate change:revisiting the rules[J]. Ecology Letters, 2006, 9:853-869.
(责任编辑 李 洁)
Effects of Tween-20 and Tween-80 on Germination ofMagnaportheoryzaeSpores
RAN Li, ZHU Zi-wei*, YANG Chao, XU You-pin, LI Pei-hua, HE Min, WANG Jing, CHEN Xue-wei, LI Wei-tao**
(Rice Research Institute, Sichuan Agricultural University, Sichuan Chengdu 611130, China)
To determine whether Tween-20 and Tween-80 affect the spore germination ofMagnaportheoryzae(M.oryzae) or not, which can guide how to make up of spore suspension and how long we may store the spore suspension, we configured different concentrations of solution using Tween-20 or Tween-80 and placed them in different times. Then detect the influence of Tween-20 and Tween-80 on spores ofM.oryzaeusing microscope. Spores ofM.oryzaecan germinate in water, Tween-20 and Tween-80 solution, and one spore may produce two or more germ tubes in the surfactant solutions, Tween-20 and Tween-80. The germination ofM.oryzaespores was more sensitive in Tween-80 than in Tween-20. The race Zhong10-8-14 with bigger size was more sensitive than the race Guy11 with smaller size for Tween-20 and Tween-80. Therefore, Tween-20 is more suitable for using as the surfactant of spore suspension. It was no problem to spray spore suspension ofM.oryzaewith no more than 0.50 % Tween-20 in 2 h because the germination rate ofM.oryzaespores did not obviously decrease.
Magnaportheoryzae; Germination; Tween-20; Tween-80
1001-4829(2016)10-2379-04
10.16213/j.cnki.scjas.2016.10.023
2015-10-27
国家自然科学基金项目(31501627, 31301626);四川省教育厅重点项目(15ZA0020);四川省教育厅国际合作计划(2014HH0066)
冉 莉(1992-),女,重庆巫溪人,硕士,主要从事水稻真菌病害的研究,E-mail:ranli8023@163.com,*为共同第一作者,**为通讯作者,E-mail:weitao-li@live.cn。
S435.11
A
