鳗鲡目鱼类线粒体蛋白质编码基因易位及系统演化关系分析
2014-10-26申欣田美孟学平程汉良阎斌伦
申欣,田美,孟学平,程汉良,阎斌伦
(1.淮海工学院 海洋学院,江苏省海洋生物技术重点实验室,江苏 连云港222005;2.中国科学院 北京生命科学研究院,北京100101)
1 引言
鳗鲡eels在分类学上隶属于辐鳍鱼纲Actinopterygii、鳗鲡目Anguillifor mes,现存约有800种,分属于4个亚目、20个科。绝大多数鳗鲡目鱼类终生生活在海洋中,仅鳗鲡属Anguilla十几种的成体生活在淡水中,但它们繁殖时仍需洄游数千千米,将卵产至深海中[1]。大多数鳗鲡目鱼类是肉食性动物,广泛分布于全球热带、亚热带和温带水域。同时,鳗鲡味道鲜美、营养丰富、肉多刺少,不少种类具有很高的肉用经济价值。鳗鲡目鱼类养殖是世界水产养殖的重要组成部分,但目前种质资源的问题十分突出,亟待关注[2]。后生动物基因组包括核基因组和线粒体基因组,线粒体基因组具有结构稳定和基因排列紧凑等优点,被广泛应用到后生动物种群遗传、生物地理及系统演化的研究中[3—6]。本文系统分析比较了现有的58个鳗鲡目鱼类线粒体基因组,为这一重要生物类群资源保护和利用提供重要的基础资料。本工作不仅有利于鳗鲡目鱼类遗传资源与进化历史的认识,而且有利于其种质资源库的建立以及优良品种的筛选。
2 材料和方法
2.1 鳗鲡目鱼类线粒体基因组
Gen Bank基因组数据库现有58种鳗鲡目鱼类线粒体基因组,包括欧洲鳗鲡Anguillaanguilla、澳洲鳗鲡A.australisaustralis、新澳鳗鲡A.austr alis schmidti、东印度洋鳗鲡A.bengalensislabiata、双色鳗鲡A.bicolorbicolor、太平洋双色鳗鲡A.bicolor pacif ica、西里伯斯鳗鲡A.celebesensis、大鳗鲡A.dief f enbachii、高原长鳍鳗A.interioris、日本鳗鲡A.j aponica、吕宋鳗鲡A.luzonensis、吕宋鳗鲡A.luzonensissp.HYT-2008、印尼鳗鲡A.mal gumor a、花鳗鲡A.mar morata、大口鳗鲡A.megastoma、非洲鳗A.mossambica、云纹鳗鲡A.nebulosanebulosa、灰鳗鲡A.obscura、澳洲长鳍鳗A.reinhar dtii、美洲鳗A.rostrata、小眼蟒鳗Pythonichthysmicrophthalmus、爱氏蚓鳗Moringuaed war dsi、小鳍蚓鳗M.microchir(鳗鲡亚目 Anguilloidei),凯氏短尾康吉鳗Colocongercadenati、细美体鳗Ariosomashiroanago、星康吉鳗Congermyriaster、哈氏异康吉鳗Heterocongerhassi、几内亚副康吉鳗Paracongernotialis、短吻颈鳗Derichthysser pentinus、鸭颈鳗Nessor hamphusingol f ianus、高眉鳝Anarchiassp.、蠕纹裸胸鳝Gy mnothor axkidako、黑身管鼻鯙Rhinomuraenaquaesita、喙吻鳗Avocettinainf ans、唇线鳗Labichthyscarinatus、线鳗Nemichthysscolopaceus、尖吻小鸭嘴鳗Facciolellaoxyrhyncha、斑点长犁齿鳗Hoplunnispunctata、弱头鳗Leptocephalussp.、小头鸭嘴鳗Nettastoma parviceps、巨斑花蛇鳗Myrichthysmaculosus、大吻沙蛇鳗Ophisur usmacror hynchos、锯犁鳗Serrivomer beanii、长齿锯齿鳗S.sector、前鳍锯犁鳗Stemonidiumhypomelas、褐泥蛇鳗Il yophisbrunneus、寄生鳗Si menchel yspar asitica、高氏合鳃鳗Synaphobr anchus kaupii(糯鳗亚目Congroidei),高体眶鼻鳗Kaupichthyshyoproroides、罗宾斯草鳗Robinsiacatherinae、龟草鳗Thalassenchelyssp.、粗犁齿海鳗Cynoponticus f erox、百吉海鳗Mur aenesoxbagio、扁身油康吉鳗Myrocongercompressus(鯙亚目 Muraenoidei),以及月尾鳗Cyemaatrum、吞鳗Eurypharynxpelecanoides、杰氏单颌鳗Monognathusjesperseni和拉文囊鳃鳗Saccopharynxlavenber gi(囊喉鱼亚目Saccopharyngoidei)[1,7—14]。
2.2 变异位点与分子标记基因分析
基于58种鳗鲡目鱼类线粒体基因组数据,分别计算4个亚目(鳗鲡亚目、糯鳗亚目、鯙亚目和囊喉鱼亚目)内部的单基因变异位点,全面揭示鳗鲡线粒体基因组的变异位点特征,以此确定合适的分子标记基因。借助于Clustal X 2.1软件[15]对所有蛋白质编码基因和核糖体RNA基因(at p6、at p8、cob、cox1-3、nad1-6、nad4L、sr RNA和lr RNA)的 核 苷酸序列,分别做多重序列比对。借助于DnaSP 5.10.1软件[16]分析鳗鲡4个亚目内部单基因的变异位点。
2.3 系统演化分析
所有13个蛋白质编码基因(cox1-3、cob、nad1-4、nad4L、nad5、nad6、atp6和atp8)的核苷酸序列首尾相连后,借助于Clustal X 2.1软件[15]做多重序列比对。对于多重序列比对结果,借助于j Model Test 2.1.3软件[17]进行嵌套似然比检验,以获得最适的替换模型。采取贝叶斯法进行系统演化关系重建,使用的软件为 Mr Bayes3.2[18]。4个马尔可夫链运行1 000 000个世代,从而保证达到收敛,将开始25%的树去除,余下75%的树进行合议,以用来估算各个分支的支持率(BPP)。
3 结果和讨论
3.1 鳗鲡目鱼类线粒体基因组主要特征
鳗鲡线粒体基因组的基因数量较为保守,在58个鳗鲡线粒体基因组中,仅有3个物种存在基因数量的差异(见表1):月尾鳗缺少一个t RNA(t RNA-Asn)基因,弱头鳗和杰氏单颌鳗均多了一个t RNA基因(分别为t RNA-Leu和t RNA-Met)。在鳗鲡亚目中,蚓鳗科的小鳍蚓鳗最短,仅有15 858 bp,异鳗科的小眼蟒鳗最长,为17 042 bp,相差1 184 bp;同时,异鳗科的小眼蟒鳗A+T含量最低(55.82%),鳗鲡科的美洲鳗最高(59.07%)。对于糯鳗亚目来讲,线粒体基因组主编码链的A+T含量差异最大,鯙科的高眉鳝A+T含量最低(53.52%),糯鳗科的星康吉鳗最高(63.18%),差距达到9.66%;锯齿鳗科的长齿锯齿鳗最短,只有16 099 bp,糯鳗科的星康吉鳗最长,为18 705 bp,相差2 606 bp。在鯙亚目中,拟鯙科的罗宾斯草鳗最短,只有16 627 bp,海鳗科的百吉海鳗最长(18 247 bp),相差1 620 bp;拟鯙科的龟草鳗A+T含量最低(56.33%),海鳗科的百吉海鳗最高(60.49%)。对于囊喉鱼亚目来讲,线粒体基因组的长度差距最大,月尾鳗科的月尾鳗最短,只有16 868 bp;单颌鳗科的杰氏单颌鳗最长,达到22 217 bp,差距高达5 349 bp;囊鳃鳗科的拉文囊鳃鳗A+T含量最低(58.52%),宽咽鱼科的吞鳗最高(60.44%)。总体比较来看,鳗鲡线粒体基因组长度介于15 858 bp(小鳍蚓鳗)与22 217 bp(杰氏单颌鳗)之间;线粒体基因组主编码链的A+T含量最低的是高眉鳝(53.52%),最高的为星康吉鳗(63.18%)(见表1)。
3.2 基因重排分析
对于线粒体基因组的主编码基因,39种鳗鲡共享脊椎动物线粒体基因组的典型排列方式[3—4,19]。在另外19个线粒体基因组中,存在主编码基因的重排(图1)。来自于糯鳗亚目的凯氏短尾康吉鳗(倭糯鳗科),细美体鳗、星康吉鳗、哈氏异康吉鳗、几内亚副康吉鳗(糯鳗科),短吻颈鳗、鸭颈鳗(长颈鳗科),尖吻小鸭嘴鳗、斑点长犁齿鳗、弱头鳗、小头鸭嘴鳗(鸭嘴鳗科),巨斑花蛇鳗、大吻沙蛇鳗(蛇鳗科),以及来自于鯙亚目的龟草鳗(拟鯙科),粗犁齿海鳗、百吉海鳗(海鳗科),这16个物种的线粒体基因组共享nad6基因的易位。在囊喉鱼亚目中,吞鳗(宽咽鱼科)和拉文囊鳃鳗(囊鳃鳗科)共享6个基因的重排:at p8、atp6、cox3、nad3、nad5和nad6;同时,杰氏单颌鳗(单颌鳗科)存在6个基因的易位:cob、nad3、nad4L、nad4、nad5和nad6(图1)。当前的数据资料显示,在鳗鲡目内部,基因排列顺序并不保守;结合系统演化分析的结果可以看出,基因重排可能会成为探讨鳗鲡内部系统演化关系的重要信息来源(详述见后文)。

图1 鳗鲡目鱼类线粒体主编码基因的基因排列(不包含转运RNA基因)
3.3 鳗鲡目鱼类线粒体基因组的变异位点及分子标记分析
基于58种鳗鲡线粒体基因组数据,分别计算4个亚目(鳗鲡亚目、糯鳗亚目、鯙亚目和囊喉鱼亚目)内部的单基因变异位点,全面揭示鳗鲡线粒体基因组的变异位点特征。在鳗鲡亚目线粒体基因组中,变异位点数最多的基因为nad5(758个),其次为nad4、cox1、lr RNA和nad2基因,变异位点的数目分别为587、558、501和474个。糯鳗亚目、鯙亚目和囊喉鱼亚目变异位点数的分析结果与鳗鲡亚目类似,5个基因(nad5、nad4、cox1、lr RNA和nad2)的 变 异位 点 数均排在前列。综合比较4个类群单基因的变异位点及变异位点率,均支持nad5、nad4和nad2基因作为cox1和lr RNA基因辅助的分子标记,为鳗鲡目鱼类分子生态和群体遗传的研究提供支持,进而为其种质资源库的建立以及优良品种的选育提供有价值的遗传信息。
3.4 系统演化历史
以线粒体所有蛋白质编码基因的核苷酸序列为数据群,采用贝叶斯法构建的系统演化树,绝大多数节点的支持率达到100%,表明利用线粒体基因组来分析和探讨鳗鲡目的系统演化关系,具有拓扑结构稳定、演化关系可靠的优点(见图2)。鳗鲡科的20个种(亚种)聚在一起,强烈支持鳗鲡科为单系群(BPP=100)。印尼鳗鲡位于鳗鲡科的基部,另外19个种(亚种)分为2支。在其中一支中,澳洲鳗鲡与新澳鳗鲡首先聚类,然后与大鳗鲡聚为一支,3种鳗鲡均属于大洋洲类群(Oceanian group,BPP=100);欧洲鳗鲡和美洲鳗鲡亲缘关系最近(BPP=100),二者组成大西洋类群(Atlantic group),[(澳洲鳗鲡+新澳鳗鲡)+大鳗鲡]与(欧洲鳗鲡+美洲鳗鲡)聚类(BPP=100),非洲鳗关系最远(见图2)。另外一支均属于印度洋-太平洋类群(Indo-Pacific group),线粒体基因组的系统演化分析结果将它们很好的聚为一支(BPP=100),这与先前的研究结果一致[7]。
线鳗科Nemichthyidae内部的系统演化关系为:(喙吻鳗+唇线鳗)+线鳗(BPP=100),在锯齿鳗科Serrivomeridae中,锯犁鳗与长齿锯齿鳗近缘,然后与前鳍锯犁鳗聚类(BPP=100)。线鳗科和锯齿鳗科亲缘关系最近,二者聚类后,与鳗鲡科构成姊妹群(BPP=100)。在囊喉鱼亚目Saccopharyngoidei中,宽咽鱼科Eurypharyngidae与囊鳃鳗科Saccopharyngidae聚类(BPP=100),同时,单颌鳗科 Monognathidae与月尾鳗科Cyematidae聚类(BPP=100),4个科聚在一支,支持囊喉鱼亚目为单系群(BPP=100)。在囊喉鱼亚目线粒体基因组中,存在主编码基因的重排(见图1)。
鳗鲡亚目蚓鳗科Moringuidae的2个物种(爱氏蚓鳗和小鳍蚓鳗)与拟鯙科Chlopsidae 2个物种(罗宾斯草鳗和高体眶鼻鳗)聚类,而且支持率很高(BPP=100),而拟鯙科的另外一个物种龟草鳗与它们关系较远,从而说明,基于线粒体基因组的系统演化分析,不支持拟鯙科为单系群。同时,龟草鳗线粒体基因组存在2个蛋白质编码基因(cob和nad6基因)的易位,因此拟鯙科的有效性值得进一步研究。海鳗科Myrocongridae的扁身油康吉鳗与鯙科Muraenidae的高眉鳝、蠕纹裸胸鳝和黑身管鼻鯙近缘(BPP=100),随后与异鳗科Heterenchelyidae的小眼蟒鳗聚为一支(BPP=100)。海鳗科的另外2个物种(粗犁齿海鳗和百吉海鳗)与蛇鳗科聚为一支(BPP=100),而且均共享cob和nad6基因的易位。因此,从线粒体基因组的数据来看,海鳗科是否是一个单系类群,值得深究。
通鳃鳗科Synaphobranchidae分类学上属于糯鳗亚目Congroidei,为海洋底栖鱼类,常被发现于1 000余米的海底[20]。线粒体基因组的系统演化结果,强烈支持通鳃鳗科的寄生鳗、高氏合鳃鳗和褐泥蛇鳗聚为一支(BPP=100),位于进化树的基部。线粒体基因组的证据表明,淡水生活的鳗鲡起源于海洋鳗鲡,同时淡水鳗鲡保留了洄游到起源地(海洋)中产卵的有趣特征[1]。
线粒体基因组的基因排列可以提供深层次的系统演化信息,鳗鲡目线粒体主编码基因的原始排列为 :sr RNA-lr RNA-nad1-nad2-cox1-cox2-atp8-atp6-cox3-nad3-nad4L-nad4-nad5-nad6-cob。16 种 鳗 鲡(细美体鳗、短吻颈鳗、凯氏短尾康吉鳗、鸭颈鳗、龟草鳗、粗犁齿海鳗、百吉海鳗、巨斑花蛇鳗、大吻沙蛇鳗、几内亚副康吉鳗、哈氏异康吉鳗、小头鸭嘴鳗、弱头鳗、斑点长犁齿鳗、尖吻小鸭嘴鳗和星康吉鳗),与鳗鲡目线粒体主编码基因的原始排列相比,共享nad6基 因 的 易 位,为:sr RNA-lr RNA-nad1-nad2-cox1-cox2-atp8-atp6-cox3-nad3-nad4L-nad4-nad5-cob
nad6(见图1)。同时,基于线粒体基因组13个蛋白质编码基因构建的系统演化树,强烈支持这16个物种聚为一支(BPP=100)。然而,由此而导致海鳗科、拟鯙科和糯鳗科是否为单系群的问题,尚待进一步探讨。

图2 基于线粒体所有蛋白质编码基因构建的系统演化关系树
4 结论
鳗鲡目鱼类线粒体基因组的基因数量较为保守,在58个鳗鲡线粒体基因组中,仅有3个物种存在基因数量的差异。在19个鳗鲡目鱼类线粒体基因组中存在主编码基因的重排。主编码基因的变异位点分析结果支持nad5、nad4和nad2基因作为cox1和lr-RNA基因辅助的分子标记。鳗鲡科的20个种(亚种)聚在一起,强烈支持鳗鲡科为单系群。鳗鲡科下属的3个类群(大洋洲类群、大西洋类群和印度洋-太平洋类群)也分别得到有力的验证。线鳗科和锯齿鳗科亲缘关系最近,二者聚类后,与鳗鲡科构成姊妹群。在囊喉鱼亚目中,宽咽鱼科与囊鳃鳗科聚类,同时,单颌鳗科与月尾鳗科聚类,4个科聚在一支,支持囊喉鱼亚目为单系群。在囊喉鱼亚目线粒体基因组中,存在主编码基因的重排。16种鳗鲡(细美体鳗、短吻颈鳗、凯氏短尾康吉鳗、鸭颈鳗、龟草鳗、粗犁齿海鳗、百吉海鳗、巨斑花蛇鳗、大吻沙蛇鳗、几内亚副康吉鳗、哈氏异康吉鳗、小头鸭嘴鳗、弱头鳗、斑点长犁齿鳗、尖吻小鸭嘴鳗和星康吉鳗),与鳗鲡目线粒体主编码基因的原始排列相比,共享nad6基因的易位。同时,基于线粒体基因组13个蛋白质编码基因构建的系统演化树,强烈支持这16个物种聚为一支。然而,由此而导致海鳗科、拟鯙科和糯鳗科是否为单系群的问题,值得今后深入探讨。
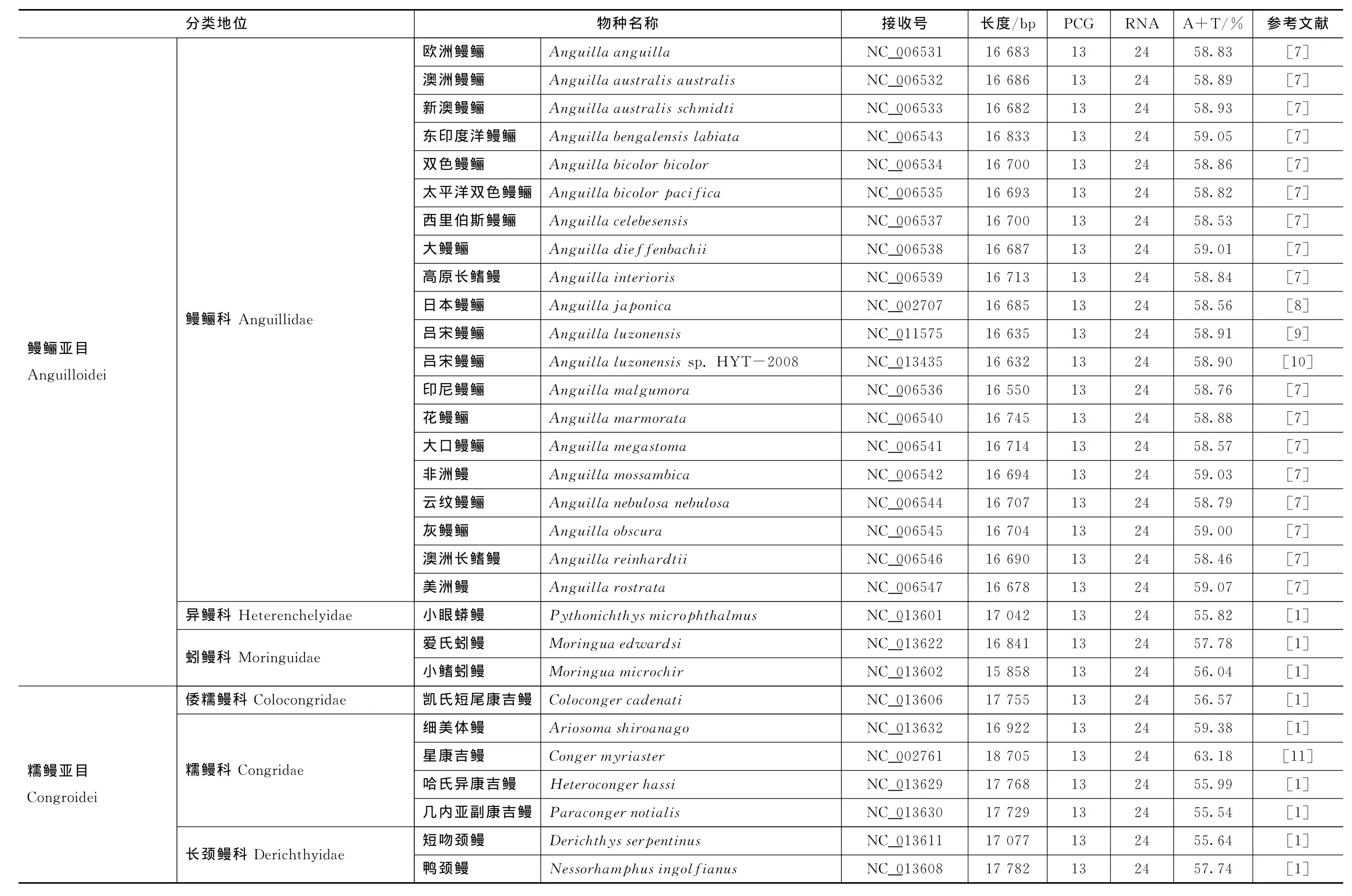
表1 鳗鲡目 鱼类线 粒体 基因组 的基本 特征
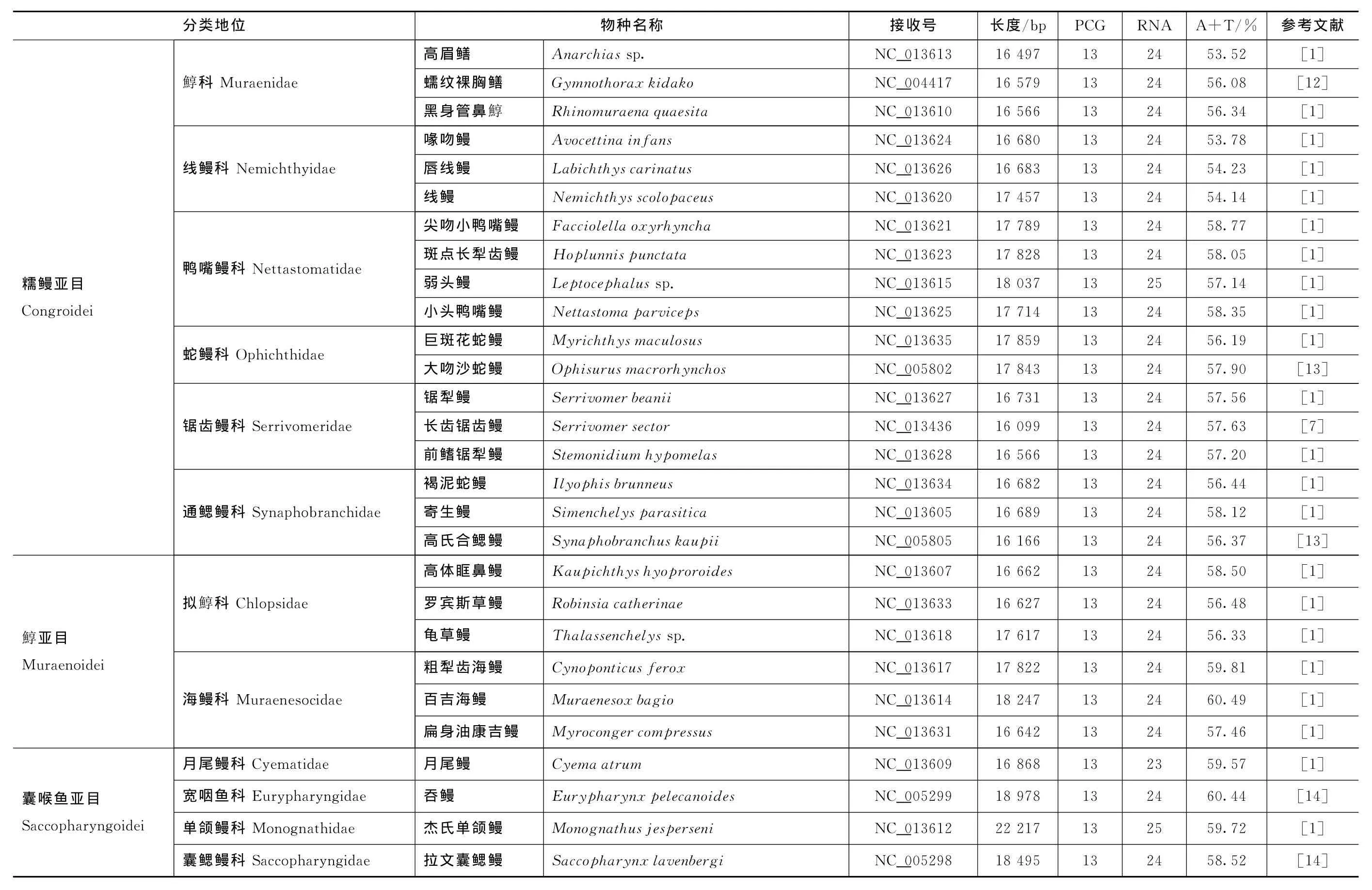
续表1
[1]Inoue J G,Miya M,Miller M J,et al.Deep-ocean origin of the freshwater eels[J].Biology Letters,2010,6(3):363—366.
[2]Lee W C,Chen Y H,Lee Y C,et al.The competitiveness of the eel aquaculture in Taiwan,Japan,and China[J].Aquaculture,2003,221(1-4):115—124.
[3]Boore J L,Brown W M.Big trees from little genomes:mitochondrial gene order as a phylogenetic tool[J].Curr Opin Genet Dev,1998,8(6):668—674.
[4]Curole J P,Kocher T D.Mitogenomics:digging deeper with complete mitochondrial genomes[J].Trends Ecol Evol,1999,14(10):394—398.
[5]Boore J L,Macey J R,Medina M.Sequencing and comparing whole mitochondrial genomes of animals[J].Methods Enzy mol,2005,395:311—348.
[6]Shen X,Ma X,Ren J,et al.A close phylogenetic relationship bet ween Sipuncula and Annelida evidenced from the complete mitochondrial genome sequence ofPhascolosomaesculenta[J].BMC Genomics,2009,10:136—146.
[7]Minegishi Y,Aoyama J,Inoue J G,et al.Molecular phylogeny and evolution of the freshwater eels genusAnguillabased on the whole mitochondrial genome sequences[J].Molecular Phylogenetics and Evolution,2005,34(1):134—146.
[8]Inoue J G,Miya M,Aoyama J,et al.Complete mitochondrial DNA sequence of the Japanese eelAnguillaj aponica[J].Fisheries Science,2001,67(1):118—125.
[9]Watanabe S,Aoyama J,Tsukamoto K.A new species of freshwater eelAnguillal uzonensis(Teleostei:Anguillidae)from Luzon Island of the Philippines[J].Fisheries Science,2009,75(2):387—392.
[10]Teng H Y,Tzeng C S,Lin Y S.A new Anguilla species and a reanalysis of t he phylogeny of freshwater eels[J].Zoological Studies,2009,48(6):808—822.
[11]Inoue J G,Miya M,Tsukamoto K,et al.Complete mitochondrial DNA sequence ofCongermyriaster(Teleostei:Anguillifor mes):novel gene order for vertebrate mitochondrial genomes and t he phylogenetic i mplications for anguillifor m families[J].Jour nal of Molecular Evolution,2001,52(4):311—320.
[12]Inoue J G,Miya M,Tsukamoto K,et al.Basal actinopterygian relationships:a mitogenomic perspective on the phylogeny of the“ancient fish”[J].Molecular Phylogenetics and Evolution,2003,26(1):110—120.
[13]Inoue J G,Miya M,Tsukamoto K,et al.Mitogenomic evidence for t he monophyly of elopomorph fishes(Teleostei)and t he evolutionary origin of the leptocephalus larva[J].Molecular Phylogenetics and Evolution,2004,32(1):274—286.
[14]Inoue J G,Miya M,Tsukamoto K,et al.Evolution of the deep-sea gulper eel mitochondrial genomes:large-scale gene rearrangements originated within the eels[J].Molecular Biology and Evolution,2003,20(11):1917—1924.
[15]Larkin M A,Blackshields G,Brown N P,et al.Clustal W and Clustal X version 2.0[J].Bioinfor matics,2007,23(21):2947—2948.
[16]Librado P,Rozas J.DnaSP v5:a soft ware for comprehensive analysis of DNA poly morphism data[J].Bioinfor matics,2009,25(11):1451—1452.
[17]Darriba D,Taboada G L,Doallo R,et al.j Model Test 2:more models,new heuristics and parallel computing[J].Nature Met hods,2012,9(8):772.
[18]Ronquist F,Teslenko M,van der Mark P,et al.Mr Bayes 3.2:efficient Bayesian phylogenetic inference and model choice across a large model space[J].Systematic Biology,2012,61(3):539—542.
[19]San Mauro D,Gower D J,Zardoya R,et al.A hotspot of gene order rearrangement by tandem duplication and random loss in t he vertebrate mitochondrial genome[J].Molecular Biology and Evolution,2006,23(1):227—234.
[20]Kar movskaya E S,Merrett N R.Taxonomy of the deep-sea eel genus,Histiobranchus(Synaphobranchidae,Anguillifor mes),with notes on the ecology ofH.bathybiusin the eastern North Atlantic[J].Journal of Fish Biology,1998,53(5):1015—1037.
