长爪沙鼠ATPase8,ATPase6,COX3基因的克隆及序列分析
2011-02-01李长龙柯贤福卢领群郭红刚萨晓婴
李长龙,柯贤福,卢领群,郭红刚,萨晓婴
(浙江省医学科学院 浙江省实验动物中心,杭州 310013)
长 爪 沙 鼠 (Mongolian Gerbil,Meriones unguiculatus)俗称蒙古沙鼠,属于啮齿目,仓鼠科,沙鼠亚科,沙鼠属,又称长爪沙土鼠、蒙古沙鼠和黄耗子等,野生长爪沙鼠主要分布于我国内蒙古及其毗邻的干旱和半干旱地区[1]。1935年大连卫生所的春日送给日本北里研究所20对长爪沙鼠并开始驯化,后引种到美、英、法等国。我国有两个主要的长爪沙鼠群体,分别保存于浙江省实验动物中心和首都医科大学[1]。长爪沙鼠具有独特的解剖学、生理学和行为学性状,对于一些疾病(如:脑缺血、癫痫、高血脂、寄生虫、细菌、病毒和老年性疾病等)的研究具有极为重要的价值,已在越来越多的研究所使用[2]。
线粒体是细胞的能源中心,在细胞能量代谢、细胞凋亡和程序化死亡中发挥重要作用。线粒体DNA (mitochondrial DNA,mtDNA)是高等动物唯一的核外遗传物质,呈共价闭合的环状双链结构。m tDNA结构简单、进化速度快(是单拷贝核基因的5-10倍),呈严格的母性遗传,遗传行为相对独立,变异发生的几率相对稳定、无组织特异性、提取方便[3]。由于mtDNA的遗传特性,已被广泛地用于物种起源与进化、生物分类及群体遗传结构等方面的研究[4]。
本研究首次对长爪沙鼠线粒体 ATPase8,ATPase6,COX3基因全序列进行测定和鉴定,并结合已公布啮齿类动物 ATPase8,ATPase6,COX3基因序列进行系统进化分析,旨在为长爪沙鼠系统进化关系和长爪沙鼠能量代谢研究提供遗传学资料,为全长线粒体测定分析奠定基础。
1 材料和方法
1.1 材料
长爪沙鼠来自浙江省实验动物中心[生产许可证号:SCXK(浙)2008-033;使用许可证号:SYXK (浙)2008-0014],长爪沙鼠解剖采集肝脏样品,-20℃保存。
1.2 总DNA提取
采用酚氯仿抽提法提取基因组DNA,0.7%琼脂糖凝胶电泳检测,-20℃保存。
1.3 引物设计和PCR扩增
分别参照长爪沙鼠 m tDNA序列 AB381914和MUU83801设计上下游引物[5-6],引物由上海美吉生物公司合成。上游(COX2 3'端)5'-TCCAACCACAGCTTTATACCG-3',下游(ND3 5'端) 5'-GGCATGGACTTTTTCTGCAT-3'。扩增片段内应包括ATPase8,ATPase6,COX3三个编码基因全序列及tRNA-Gly和tRNA-Lys等两个tRNA基因全序列。
扩增反应体系总体积为25μL:模板DNA 100 ng、10×buffer 2.5μL、MgC12(2.5 mol/L)2μL、dNTP 2μL、上下游引物(10 pmol/L)各1μL、1 U pfuDNA聚合酶,加灭菌纯水补足。扩增条件:94℃预变性5 min,94℃变性30 s,56.5℃复性30 s,72℃延伸1 min,30个循环,72℃延伸7 min。
1.4 序列纯化、测序、拼接
用北京天根胶回收试剂盒对PCR扩增产物进行纯化,纯化后的产物由上海美季公司完成测序工作,Chromas 2.22校对测序图,DNAMAN5.5拼接序列。
1.5 序列的鉴定和分析
从GenBank下载其他具有完整m tDNA基因组序列的18种啮齿类动物类的 ATPase8,ATPase6,COX3序列,GenBank登录号分别为:NC_005089家鼠(Musmusculus)、NC_012387小家鼠(Musmusculus castaneus)、NC_006914西欧家鼠(Mus musculus domesticus)、NC_006915日本小鼠(Mus musculus molossinus)、NC_010339东欧家鼠(Mus musculus musculus)、NC_012389缅鼠(Rattus exulans)、NC_ 012461帅家鼠(Rattus praetor)、NC_012374黑家鼠(Rattus rattus)、NC_011638达氏家鼠 (Rattus tanezum i)、AC_000022褐家鼠(Rattus norvegicus)、NC_013068大仓鼠(Tscherskia triton)、NC_007936中国仓鼠(Cricetulus griseus)、NC_003041台湾田鼠(Microtus kikuchii)、NC_001892胖睡鼠(Glis)、NC_ 005314非洲跳鼠(Jaculus)、NC_009056鳞尾松树(Anomalurus)、NC_000884豚鼠(Cavia porcellus)和NC_002658蔗鼠(Thryonomys swinderianus)等。用DNAMAN5.5进行同源性分析;用 EMBOSS中compseq程序统计碱基含量和计算 A和 G偏离[7];用MEGA 4.0统计物种间遗传距离,并基于Kimura-Parameter双参数模型,用UPGMA和最小进化法构建分子系统进化树[8]。
2 结果与分析
2.1 长爪沙鼠线粒体基因克隆和测序
从长爪沙鼠基因组 DNA中特异性克隆获得1.9 kb的片段(图 1)。经过测序获得长爪沙鼠m tDNA序列1952 bp,其中包括ATPase8,ATPase6,COX3编码基因全序列、tRNA-Gly和tRNA-Lys转运RNA基因序列。
2.2 长爪沙鼠 ATPase8基因的鉴定及遗传进化分析
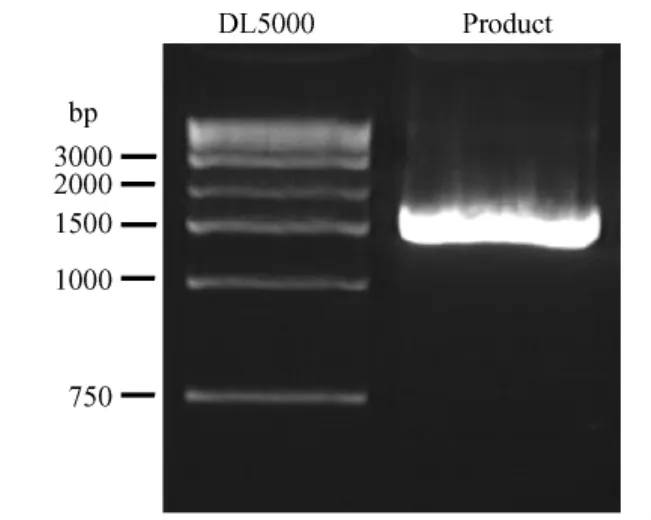
图1 PCR扩增长爪沙鼠线粒体DNA部分序列注:泳道DL5000为DL5000 Marker,泳道Product为线粒体DNA部分序列扩增结果Fig.1 The result of PCR amplified the partical sequence ofm tDNA in Mongolian GerbilNote:The line 1 is DL5000 Marker,the line 2 is product of PCR amplified the partical sequence ofmtDNA

图2 长爪沙鼠ATPase8与其他啮齿类动物同源性分析Fig.2 The homology analysis between Mongolian Gerbil ATPase8 yene and other rodents
长爪沙鼠ATPase8基因序列与其他啮齿动物的基因序列分别进行序列比对、同源性分析和进化分析。结果显示,长爪沙鼠 ATPase8基因序列全长204 bp,编码67个氨基酸的序列。COX3基因与家鼠同源性为90%、与小家鼠同源性为98%,与仓鼠同源性为83%(图2)。
利用MEGE4.1软件将长爪沙鼠和18个啮齿类动物的ATPase8基因进行系统进化分析。结果显示,家鼠、小家鼠分别聚合与A,B两支,之后与长爪沙鼠分支聚合,之后依次为台湾田鼠,非洲跳鼠,仓鼠等分支聚合(图3)。分枝长表示分歧度,枝上的数值为1000次重复抽样检验得到的支持率。分别选择家鼠、黑家鼠、中国仓鼠、豚鼠、蔗鼠和睡鼠与长爪沙鼠计算遗传距离。基于ATPase8基因结果显示(表1),长爪沙鼠与家鼠遗传距离最近为0.633;与黑家鼠遗传距离为0.685;与中国仓鼠遗传距离为0.867(表1)。
2.3 长爪沙鼠 ATPase6基因的鉴定及遗传进化分析
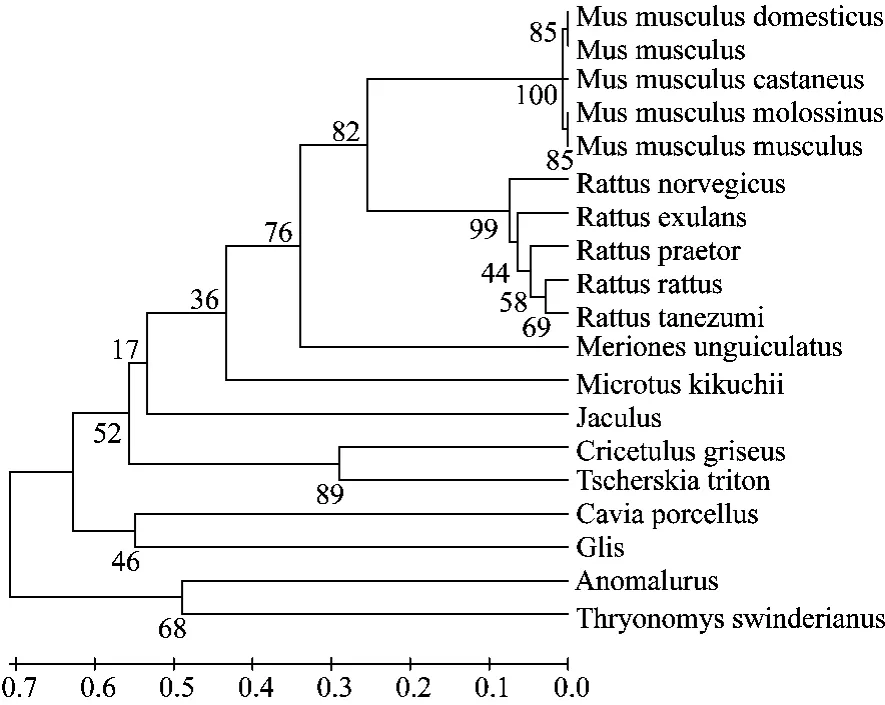
图3 UPGMA法构建的ATPase8基因系统发生树Fig.3 Phylogenetic tree based on ATPase8 sequences by UPGMA methods
长爪沙鼠ATPase6基因序列与其他啮齿动物的基因序列分别进行序列比对、同源性分析和进化分析。结果显示,长爪沙鼠 ATPase6基因序列全长681 bp,编码226个氨基酸的序列。ATPase6基因与家鼠同源性为89%、与小家鼠同源性为98%,与仓鼠同源性为76%(图4)。
利用MEGE4.1软件将长爪沙鼠和18个啮齿类动物的ATPase6基因序列进行系统进化分析。结果显示,家鼠、小家鼠分别聚合与A,B两支,之后与中国仓鼠等仓鼠分支聚合,之后依次为长爪沙鼠等分支聚合(图5)。分别选择家鼠、黑家鼠、中国仓鼠、豚鼠、蔗鼠和睡鼠与长爪沙鼠计算遗传距离。结果显示(表 1),长爪沙鼠与家鼠遗传距离最近为0.273;与黑家鼠遗传距离为0.281;与中国仓鼠遗传距离为0.29(表1)。

图4 长爪沙鼠ATPase6与其他啮齿类动物的同源性分析Fig.4 The homology analysis between Mongolian Gerbil ATPase6 gene and other rodents

图5 UPGMA法构建的ATPase6基因系统发生树Fig.5 Phylogenetic tree based on ATPase6 sequences by UPGMA methods
2.4 长爪沙鼠COX3基因的鉴定及遗传进化分析
长爪沙鼠COX3基因序列与其他啮齿动物的基因序列分别进行序列比对、同源性分析和进化分析。结果显示,长爪沙鼠 COX3基因序列全长784 bp,编码261个氨基酸的序列。COX3基因与家鼠同源性为90%;与小家鼠同源性为98%;与仓鼠同源性为80%(图6)。
利用MEGE4.1软件将长爪沙鼠和18个啮齿类动物的COX3基因进行系统进化分析。结果显示,家鼠、小家鼠分别聚合与A,B两支,之后与中国仓鼠等仓鼠分支聚合,之后依次为长爪沙鼠等分支聚合(图7)。分别选择家鼠、黑家鼠、中国仓鼠、豚鼠、蔗鼠和睡鼠与长爪沙鼠计算遗传距离。基于COX3基因结果显示(表1),长爪沙鼠与家鼠遗传距离最近为0.211;与黑家鼠遗传距离为0.231;与中国仓鼠遗传距离为0.244。基于ATPase8基因结果显示(表1)

图6 长爪沙鼠COX3与其他啮齿类动物的同源性分析Fig.6 The homology analysis between Mongolian Gerbil COX3 gene and other rodents

图7 UPGMA法构建的COX3基因系统发生树Fig.7 Phylogenetic tree based on COX3 sequences by UPGMA methods
2.5 基因序列组成分析
用EMBOSS中compseq程序统计长爪沙鼠3个基因的碱基含量和碱基偏离。长爪沙鼠ATPase8,ATPase6,COX3三个基因的中G(%)均为含量最少的碱基;A+T含量分别为59.2、64.1和68.2,均明显高于 G+C的含量。A-skew的范围为 -0.256 (COX3)至 -0.568(ATPase8);G-skew的范围为-0.030(COX3)至0.058(ATPase8);碱基偏倚明显(表2)。
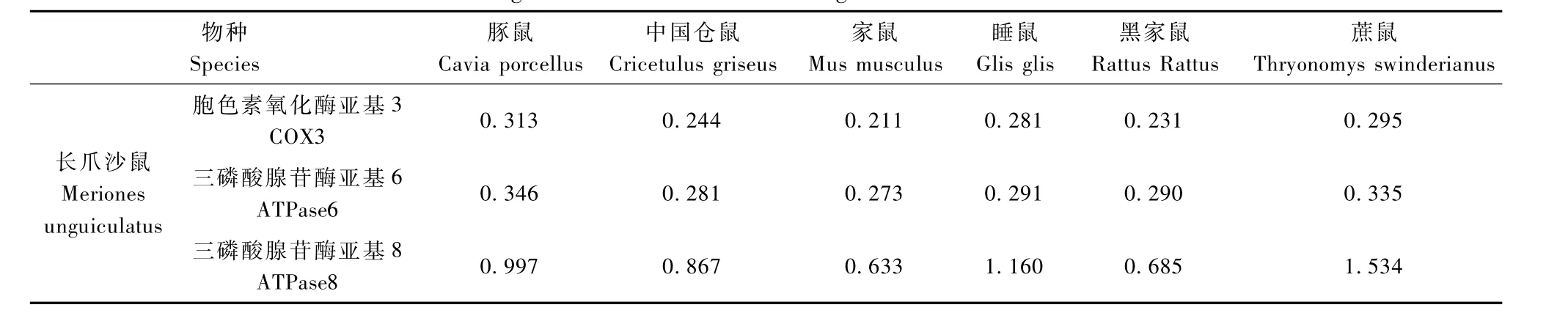
表1 长爪沙鼠与其他鼠类遗传距离Tab.1 The genetic distance between Mongolian Gerbil and other rodents

表2 长爪沙鼠ATPase8,ATPase6,COX3编码序列的碱基组成Tab.2 The nucleotide composition in COX3,ATPase6,ATPase8 gene sequence of Mongolian Gerbil
3 讨论
3.1 长爪沙鼠 ATPase8,ATPase6,COX3基因序列的测定和鉴定
哺乳动物m tDNA内包含13个编码基因,均参与呼吸的电子传递[9]。三磷酸腺苷酶亚基 6、8 (adenosine triphosphatase 6、8,ATPase6、ATPase8)和胞色素氧化酶亚基3(cytochrome oxidase,COX3)在m tDNA上呈相邻分布,均可用于分子进化研究[10-12]。ATPase是生物体内广泛存在细胞膜上的一种极为重要的酶,功能主要是维持细胞内外离子及渗透压和跨膜电化学平衡以及细胞能量代谢。ATPase 8和 ATPase 6基因是编码 ATPase(又称F0F1-ATPase酶复合物)F0部分的基因,是呼吸链电子传递中不可或缺的部分[3]。COX3是线粒体呼吸链电子传递的终末复合物,属亚铁血红素,铜氧化酶的超家族,是线粒体氧化能力的关键调节部位[13]。
m tDNA是闭合的双链环状 DNA,ATPase8,ATPase6,COX3依次排列于 m tDNA中段,在 COX2和ND3基因之间。本研究根据已知的长爪沙鼠COX2和ND3基因部分序列分别设计上下游引物,并在基因组DNA中扩增出约1.9 kb的DNA片段(图1)。共测得长爪沙鼠m tDNA序列1952 bp,其中包括ATPase8,ATPase6,COX3编码基因全序列、tRNA-Gly和 tRNA-Lys转运 RNA基因序列。序列比对和同源分析结果显示,长爪沙鼠线粒体COX3序列全长为 784bp,ATPase6序列全长为 681 bp,ATPase8序列全长为 204 bp(图 8);ATPase8和 ATPase6基因间存在43 bp的碱基重叠,ATPase6和COX3基因间存在1 bp的碱基重叠。本研究选择18种不同的啮齿类动物线粒体 ATPase8,ATPase6,COX3基因序列进行同源性分析,结果显示,长爪沙鼠3个基因与家鼠、小家鼠和仓鼠序列同源性在76%~98%之间。线粒体基因由于没有组氨酸保护,而且处于高氧化条件之下,其突变频率较核基因高[3],因此认定上述序列为长爪沙鼠 ATPase8,ATPase6,COX3基因全序列。
3.2 长爪沙鼠线粒体DNA序列及进化分析
动物的起源进化长期以来一直困扰着遗传学家。分子遗传学方法为研究群体的起源和进化开辟了新的途径[7]。m tDNA比核DNA的突变率高5~10倍[8]。利用m tDNA序列构建的系统发育树能很好的反映动物母系的迁徙和进化。许多科学家利用m tDNA序列差异来研究动物的遗传多样性与起源演化,取得了一系列令人瞩目的成果[14]。
不同物种的mtDNA碱基组成和特性存在明显的差异[15]。DNA的鸟嘌呤和胞嘧啶的碱基含量(G +C含量)在不同物种的基因组中波动范围广阔[16],甚至在同一物种的基因组不同区域也不相同。这种基因组局部碱基不均衡的现象表现为不同区域的核苷酸碱基组成存在差异[17]。长爪沙鼠ATPase8,ATPase6,COX3基因编码序列中碱基 G的含量分别为6.9%,10.7%,15.2%,明显低于其他碱基的含量m tDNA的一个显著特征[18];A+T含量分别为68.2%,64.1%,59.2%,符合哺乳动物A、T含量高,G、C含量低的特点相似[19]。
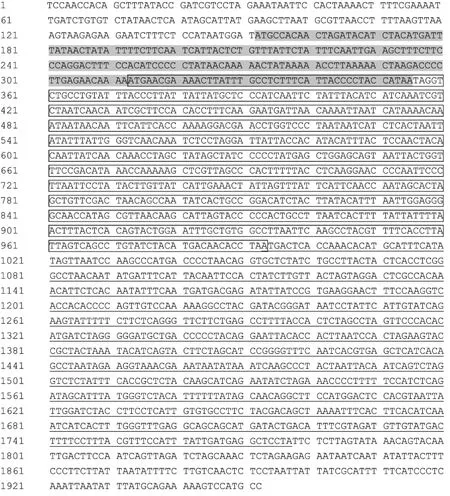
图8 长爪沙鼠ATPase8,ATPase6,COX3基因序列Fig.8 The complete sequence of ATPase8,ATPase6,COX3 gene in Mongolian Gerbil序列阴影部分为
Chevret等[20]曾利用 DNA在溶解温度的方法研究长爪沙鼠与家鼠和小家鼠的进化关系,结果显示长爪沙鼠与家鼠和小家鼠存在较相近的进化关系。Colangelo等[21]利用cyt B和16S rRNA具有序列研究在非洲的沙鼠的分子进化关系;Chevret等[22]利用cyt B和12S rRNA基因序列研究沙鼠亚科内分子进化关系。李长龙等[2]利用 D-Loop区序列分析长爪沙鼠,结果显示长爪沙鼠与家鼠、小家鼠和仓鼠存在较相近的进化关系。
本研究应用 MEGA 4.0软件,分别基于ATPase8,ATPase6,COX3基因全序列,用 UPGMA法构建系统进化树得到相似的拓扑结构,结果显示,家鼠和小家鼠均分别聚合为一支。在基于ATPase8和 COX3基因构建的进化树中,长爪沙鼠最先与家鼠和小家鼠的分支聚合,之后与仓鼠分支结合;在基于ATPase8基因构建的进化树中,仓鼠属的3个动物先与家鼠和小家鼠的分支聚合,之后在与长爪沙鼠分支聚合。说明家鼠、小家鼠、仓鼠和长爪沙鼠遗传关系较近。分别基于 ATPase8,ATPase6,COX3基因的遗传距离分析结果基本验证了进化分析结果,长爪沙鼠与家鼠的遗传距离最近,与黑家鼠和仓鼠的遗传距离较近,与蔗鼠、豚鼠、睡鼠等其他动物遗传距离较远。
从3个基因编码序列分析显示,基于相同动物不同基因构建的系统发生树存在一定的差异。于燕等发现,基于黑鲈不同脂代谢相关基因构建的系统发生数结果也存在不同,研究者认为其原因是基因进化速率的不同所导致。研究者认为组织分布单一、功能简单、直接作用于靶分子的蛋白质、其编码基因序列变化大,适于反映近缘物种的亲缘关系;组织分布广、功能多样、作用机理复杂的基因适于分析远缘物种间的亲缘关系[23]。利用长爪沙鼠D-Loop区序列构建的系统发生树与本研究也存在不同[2],其原因可能是 D-loop区的进化速率是m tDNA其他区域的3~5倍多导致。但是,以上数据根据m tDNA部分序列计算得出,进一步研究应采用m tDNA全序列信息并增加动物数量,在充分参考考古学数据的基础上进行综合分析。
[1]方喜业,邢瑞昌,贺争鸣.实验动物质量控制[M].北京:中国标准出版社,2008:913-916.
[2]李长龙,卢领群,郭红刚,等.长爪沙鼠线粒体DNA控制区全序列测定及分析[J].中国比较医学杂志,2010,20(4): 40-45.
[3]徐其放,陈嘉昌,朱世杰,等.动物线粒体DNA的特异结构及应用分子系统学分析的方法[J].中国比较医学杂志,2005,15(5):315-319.
[4]廖顺尧,鲁成.动物线粒体基因组研究进展[J].生物化学与生物物理进展,2000,27(5):508-512.
[5]Martin Y,Gerlach G,Schlotterer C,et al.Molecular phylogeny of European muroid rodents based on complete cytochrome b sequences[J].Mol Phylogenet Evol,2000,16(1):37-47.
[6]Chevret P,Dobigny G.Systematics and evolution of the subfamily Gerbillinae(Mammalia,Rodentia,Muridae)[J].Mol Phylogenet Evol,2005,35(3):674-688.
[7]Salvato P,Simonato M,Battisti A,et al.The complete mitochondrial genome of the bag-shelter moth Ochrogaster lunifer (Lepidoptera,Notodontidae)[J].BMC Genomics,2008,15 (9):331-346.
[8]Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular Evolutionary Genetics Analysis(MEGA)software version 4.0[J].Mol Biol Evol,2007,24(8):1596-1599.
[9]Goios A,Pereira L,Bogue M,Macaulay V,Amorim A.m tDNA phylogeny and evolution of laboratory mouse strains[J].Genome Res,2007;17(3):293-8.
[10]管敏强,曹琼洁,陈忠义,等.线粒体DNA序列分析封闭群小鼠遗传稳定性[J].实验动物与比较医学,2009,29(2): 113-116.
[11]宋宏图,李兵,沈卫德.老挝家蚕线粒体cox3基因序列分析及分子进化研究初探[J].江苏蚕业,2006,28(4):14-17.
[12]邵爱华,杜建,陈葵,等.暗纹东方纯线粒体 ATPase8和ATPase6基因的克隆与序列分析[J].苏州科技学院学报:自然科学版,2010,27(1):41-46.
[13]江海洪,谢燕,刘情予.细胞色素c氧化酶的分子生物学研究进展[J].国外医学、生理、病理科学与临床分册,2001,21 (1):20-22.
[14]Moreira MA,Bonvicino CR,Soares MA,Seuánez HN.Genetic diversity of neotropical primates:phylogeny,population genetics,and animalmodels for infectious diseases[J].Cytogenet Genome Res,2010,128(1-3):88-98.
[15]高英凯,苗永旺,苏小茜,等.74种鸟类线粒体基因组碱基组成及特征分析[J].云南农业大学学报:自然科学版,2009,24(1):51-58.
[16]Daubin V,Perrière G.G+C3 structuring along the genome:a common feature in prokaryotes[J].Mol Biol Evol,2003,20 (4):471-483.
[17]Sueoka N.Two aspects of DNA base composition:G+C content and translation-coupled deviation from intra-strand rule of A=T and G=C[J].JMo Evo,1999,49(1):49-62.
[18]樊乃昌,施银柱.中国鼢鼠(Eospalax)亚属分类研究[J].兽类学报,1982,2(2):183-197.
[19]杨路存,陈桂琛,刘荣堂,等.3种鼢鼠mtDNA控制区结构及其系统发育分析[J].草业科学,2009,26(2):100-106.
[20]Chevret P,Denys C,Jaeger JJ,et al.Molecular evidence that the spiny mouse(Acomys)is more closely related to gerbils (Gerbillinae)than to truemice(Murinae)[J].Proc Natl Acad Sci USA,1993,90:3433-3436.
[21]Colangelo P, Granjon L, Taylor PJ, et al. Evolutionary systematics in African gerbilline rodents of the genus Gerbilliscus:Inference from mitochondrial genes[J].Mol Phylogenet Evol,2007,42(3):797-806.
[22]Chevret P,Dobigny G.Systematics and evolution of the subfamily Gerbillinae(Mammalia,Rodentia,Muridae)[J].Mol Phylogenet Evol,2005,35(3):674-688.
[23]于燕,梁旭方,李诗盈,等.大口黑鲈脂代谢相关基因NPY、UCP2、LPL、HL克隆与分子进化分析[J].水生生物学学报,2008,32(6):900-907.
