转录因子NbMYB1R1通过促进活性氧积累抑制病毒侵染
2024-04-24姜兴林于连伟付涵艾妞崔荧钧李好海夏子豪袁虹霞李洪连杨雪施艳
姜兴林,于连伟,付涵,艾妞,崔荧钧,李好海,夏子豪,袁虹霞,李洪连,3,4,杨雪,2,施艳
转录因子NbMYB1R1通过促进活性氧积累抑制病毒侵染
姜兴林1,于连伟1,付涵1,艾妞1,崔荧钧5,李好海6,夏子豪7,袁虹霞1,李洪连1,3,4,杨雪1,2,施艳
1河南农业大学植物保护学院,郑州 450002;2河南农业大学作物学博士后流动站,郑州 450002;3河南省粮食作物协同创新中心,郑州 450002;4小麦玉米作物学国家重点实验室,郑州 450002;5河南省植物保护新技术推广协会,郑州 450002;6河南省植物保护检疫站,郑州 450002;7沈阳农业大学植物保护学院,沈阳 110866
【背景】黄瓜绿斑驳花叶病毒(cucumber green mottle mosaic virus,CGMMV)是我国重要的检疫性植物病毒,严重降低了世界范围内蔬菜以及瓜类的产量。MYB蛋白家族庞大,功能多样,存在于所有真核生物中。大多数MYB作为转录因子控制植物的发育和代谢,并对植物应对生物和非生物胁迫反应起重要的调控作用。前期研究显示CGMMV侵染后可以显著上调寄主转录因子基因的表达。【目的】明确NbMYB1R1参与CGMMV侵染的机制,为CGMMV病害防控提供理论依据。【方法】运用MEGA 7.0构建系统进化树,对NbMYB1R1的氨基酸序列进行系统进化分析;通过构建的荧光表达载体,转化GV3101农杆菌后浸润本氏烟叶片,激光共聚焦显微镜观察其亚细胞定位;利用qRT-PCR技术分析在CGMMV侵染不同时期的转录水平及沉默烟草植株中活性氧(reactive oxygen species,ROS)相关基因的转录变化;通过沉默及下游调控基因和,分析及下游调控基因和在CGMMV侵染过程中的作用;瞬时过表达NbMYB1R1和NbMYB1R1关键氨基酸突变体,分析NbMYB1R1对CGMMV侵染的影响;使用台盼蓝染色以及DAB染色观察瞬时过表达NbMYB1R1造成的细胞死亡是否与程序性细胞死亡(programmed cell death,PCD)以及ROS的积累有关;运用酵母双杂交技术验证NbMYB1R1是否与CGMMV相关蛋白互作。【结果】系统进化树分析表明,NbMYB1R1属于1R MYB大类并与多种烟草的MYB转录因子同源性极高;亚细胞定位结果显示NbMYB1R1定位于细胞核;在CGMMV侵染的烟草植株中,的转录水平对比健康植株有明显变化,在CGMMV侵染8、12 d时的转录水平显著上调;在沉默内源基因的植株上接种CGMMV,3 d后沉默植株系统叶出现斑驳、卷曲症状,而对照植株在3.5 d时才出现症状;同时CGMMV CP mRNA水平和蛋白水平检测结果也表明沉默可以有效促进CGMMV的积累;在本氏烟叶片瞬时过表达NbMYB1R1及其突变体3 d时检测CGMMV蛋白水平表达量,结果显示过表达NbMYB1R1可以有效抑制CGMMV的侵染,DNA结合结构域缺失后会减轻NbMYB1R1对CGMMV的抑制;台盼蓝和DAB染色结果表明,在瞬时过表达NbMYB1R1蛋白后导致ROS积累并引起细胞死亡;在沉默内源基因的植株叶片上检测ROS相关基因的转录水平,发现交替氧化酶基因、转录水平显著上调;在沉默内源基因和的植株上接种CGMMV,4 d后和沉默植株系统叶出现斑驳、卷曲症状,而对照植株在3.5 d时就出现症状;同时CGMMV CP mRNA水平和蛋白水平检测结果也表明沉默和可以有效抑制CGMMV的积累;酵母双杂交结果显示NbMYB1R1不与CGMMV编码蛋白直接相互作用。【结论】随着CGMMV侵染,防御相关基因表达上调,从而抑制下游基因、的表达并激活细胞内产生ROS抑制病毒侵染,但NbMYB1R1不是通过与CGMMV病毒蛋白直接互作而产生此作用。由此可见,NbMYB1R1在CGMMV侵染过程中发挥了重要作用。
黄瓜绿斑驳花叶病毒;;转录因子;致病机制;活性氧
0 引言
【研究意义】我国是世界上最大的蔬菜生产国之一,黄瓜等葫芦科作物是我国重要的经济作物[1]。黄瓜绿斑驳花叶病毒(cucumber green mottle mosaic virus,CGMMV)引起的病害是我国葫芦科作物上重要的检疫性病毒病。葫芦科作物感染CGMMV后会导致减产,从而影响经济价值[2]。挖掘植物中参与病毒侵染的寄主因子对抗病机制解析及抗病品种选育具有重要意义。【前人研究进展】MYB(v-myb avian myeloblastosis viral oncogene homolog)家族是真核生物中一个庞大的、普遍存在的转录因子(TF)家族。MYB转录因子最初是在禽髓母细胞中发现的[3]。1987年,在玉米中首次发现植物MYB转录因子()[4]。之后,许多植物中的MYB转录因子被鉴定出来,例如拟南芥、棉花、小麦、水稻和番茄等[5-8]。MYB转录因子具有一个高度保守的DNA结合域(DNA binding domain,DBD)[9]。DBD是区分MYB家族的重要区域,MYB转录因子就是通过该区域的空间结构与DNA结合[10]。MYB家族蛋白也是通过该区域的3个不完全重复序列,即R1、R2和R3进行分类。根据MYB结构域的数量和位置,MYB转录因子被分为4组:1R-MYB(R1/R2, R3-MYB)、2R-MYB(R2R3-MYB)、3R-MYB(R1R2R3-MYB)和4R-MYB(R1/R2-MYB)[11]。1R-MYB是一种重要的端粒结合因子,负责维持染色体结构的完整性和调节基因转录[12]。2R-MYB又称R2R3-MYB,在植物中2R-MYB含量最多,功能也最丰富。R2R3-MYB参与细胞分化、器官形成、叶片形态改变、调节植物激素、次生代谢以及对各种应激源的响应[11]。3R-MYB主要调控细胞周期、细胞分化和植物对胁迫的耐受性[13]。4R-MYB仅在拟南芥、葡萄和毛杨中发现,其功能还未被发掘[14-16]。在植物中许多MYB转录因子的转录被诱导以应对病原体的攻击。例如番茄MYB基因在转基因烟草中的表达提高了对尖镰孢()和灰霉病菌()的抗性[17],GhMYB36在拟南芥和棉花中均可增强其对黄萎病菌()的抗性[18];GhMYB4也可增强棉花对黄萎病菌的抗性[19];PnMYB2能够增强三七对根腐病病原体茄病镰孢()的抗性[20];番茄MYB49增强植物对致病疫霉()的抗性[21];SmMYB44能够提高茄类植物对青枯病菌()的耐受性[22]。同时一些MYB转录因子在植物对抗病原物的过程中起着负调控的作用。例如AtMYB44负调控植物对软腐病的耐受性[23]。MYB转录因子在植物防御病毒入侵反应中也起到了重要作用。例如在烟草花叶病毒(tobacco mosaic virus,TMV)感染期间,观察到和转录上调,和的表达可以增强烟草对于TMV的抗性[24-25];SlMYB28作为负调节因子参与了番茄黄化曲叶病毒(tomato yellow leaf curl virus,TYLCV)感染的调控[26];在感染番茄曲叶病毒(tomato leaf curl virus,ToLCV)的番茄植株中,观察到MYB33转录水平较低[27];在拟南芥中表达花椰菜花叶病毒(cauliflower mosaic virus,CaMV)基因VI(P6)后,诱导了基因的上调[28];在黄瓜中,CGMMV感染后已经发现6种差异表达的MYB转录因子,在侵染早期这些MYB转录因子在表达上出现上调的趋势,暗示它们对在植物响应CGMMV过程中起重要的调节作用[29]。植物为了避免受到病原体的侵害,进化出了多重防御系统来保护自身。这些防御机制中最重要的一步是快速识别入侵者和调配资源[30]。过敏性细胞坏死(hypersensitive response,HR)是植物免疫反应中最有效、最快速的防御反应之一[31]。HR反应是直接与病原体接触或靠近病原体的植物细胞迅速死亡从而将病原体限制在坏死细胞内阻止病原菌的扩散[32]。有研究表明MYB转录因子可以通过诱导HR反应来增强植物的抗病性。如在拟南芥中,AtMYB30作为HR反应的正调节因子来增强植物抗病性[32-33];辣椒中的CaPHL8正向调节辣椒防御反应,并伴随HR反应[34];以及油菜中BnaMYB111L诱导HR反应,并诱导一系列防御相关基因上调[35]。HR反应会诱导活性氧(reactive oxygen species,ROS)爆发从而引起细胞死亡[36]。BnaMYB111L表达诱导油菜籽原生质体中ROS积累以及细胞死亡[35]。因此,笔者推测MYB转录因子可以通过诱导ROS积累以及HR来增强自身抗病。【本研究切入点】通过RNA-Seq数据分析发现,CGMMV侵染后会特异性上调本氏烟()内源基因表达,该基因属于MYB转录因子家族。目前暂无研究报道NbMYB1R1蛋白参与病毒侵染。【拟解决的关键问题】探索转录因子NbMYB1R1参与CGMMV侵染的机制,为CGMMV防治提供理论依据。
1 材料与方法
试验于2022年5月至2023年10月在河南农业大学植物保护学院完成。
1.1 材料
本氏烟及核定位H2B转基因烟草种植于光周期为16 h光照/8 h黑暗、相对湿度为60%、温度为25 ℃的温室。含CGMMV侵染性克隆菌株、GV3101菌株保存于本实验室-80 ℃超低温冰箱。大肠杆菌DH5感受态购自擎科生物,DAB和台盼蓝染料购自索莱宝生物科技有限公司。载体TRV-RNA1、TRV-RNA2、pEG103、pGADT7和pGBKT7均由本实验室保存。供试引物见表1。
1.2 NbMYB1R1同源进化树的构建
在NCBI网站查找不同植物中MYB的氨基酸序列,利用DNAMAN软件对NbMYB1R1的氨基酸序列进行Blast比对,对同源性高的氨基酸序列采用极大似然法(maximum likelihood)进行系统进化分析。利用MEGA 7.0软件建立系统发育树。用于建立系统发育树的基因登录号见表2。
1.3 亚细胞定位
使用引物-GFP-F/-GFP-R,以本氏烟cDNA为模板进行RT-PCR扩增,将扩增出的片段利用Gateway方法克隆至pEG103载体,构建好的质粒电击转化GV3101农杆菌感受态,将含有NbMYB1R1-GFP质粒的农杆菌悬浮于瞬时转染缓冲液(10 mmol·L-1MgCl2,10 mmol·L-1MES,200 μmol·L-1乙酰丁香酮,pH 5.6),调节菌液浓度OD600为0.5,挑选28 d H2B转基因本氏烟植株浸润,进行瞬时表达。农杆菌浸润48 h后,在共聚焦显微镜下进行观察,绿色荧光蛋白GFP的激发光为488 nm,红色荧光蛋白RFP的激发光为560 nm。
1.4 VIGS系统沉默
通过TRV侵染性克隆介导的VIGS技术对、和进行系统沉默。以和与的基因同源区域为模板,利用网站SGN VIGS Tool(http://vigs.solgenomics. net)预测该基因上一个300 bp左右的片段,根据该片段设计引物VIGS-F/VIGS-R和VIGS-F/VIGS-R,利用Gateway方法将该片段克隆至TRV-RNA2载体,构建好的质粒电击转化农杆菌GV3101,得到含有TRV-RNA2和TRV-RNA2载体的农杆菌,摇菌TRV-RNA1、TRV-RNA2、TRV-RNA2-和TRV-RNA2-农杆菌,用瞬时转染缓冲液将其悬浮OD600为1.0。1﹕1混合浸润,TRV-RNA1和TRV-RNA2-农杆菌以及TRV-RNA1和TRV-RNA2-/农杆菌共浸为处理组,TRV-RNA1和TRV-RNA2农杆菌共浸为对照组。每组处理至少8个生物学重复。
1.5 病毒接种
将含有CGMMV侵染性克隆的农杆菌菌液在含Kan+Rif(50 μg·mL-1)的LB平板上划线活化,28 ℃培养箱倒置培养1.5—2 d。挑取多克隆于5 mL含Kan+Rif(50 μg·mL-1)抗性的LB液体培养基中,28 ℃,220 r/min摇床振荡培养过夜。用瞬时转染缓冲液将其悬浮OD600调为1.0,浸润健康本氏烟植株,等待6—8 d后发病。将发病的系统叶取下用石英砂磨碎加水混匀,并摩擦接种到本氏烟植株上。
1.6 突变体的构建
以为模板,利用网站预测该基因上的关键功能的氨基酸位点为第121—127位氨基酸。在该位点的前端以及后端设计引物121-127VIGS-F/121-127VIGS-R,以为模板扩增片段,利用同源重组的方法将片段克隆到GFP载体上,得到缺失121—127位氨基酸的载体121-127-GFP。
1.7 qRT-PCR分析
植物总RNA的提取使用Trizol法,利用HiScript&III RT SuperMix for qPCR(+gDNA wiper)试剂盒将提取的总RNA反转录为cDNA。在无酶的八连排中配制如下混合液:2×ChamQ Universal SYBR qPCR Master Mix 5 μL,正向与反向引物各0.2 μL,cDNA 1 μL,ddH2O 3.6 μL。在实时荧光定量PCR仪中进行反应,反应程序如下:预变性95 ℃ 30 s;95 ℃ 10 s、56 ℃ 30 s、72 ℃ 30 s,进行40个循环;添加熔解曲线。得到数据后,采用2-ΔΔCT方法进行分析。
1.8 Western blot分析
将样品装入2 mL离心管中,液氮降温并置于研磨仪上破碎。植物组织变成粉末后加入适量的蛋白裂解液,冰上充分裂解30 min,低温离心后取上清液并加入5×Loading buffer,放入沸水中变性。将变性后的蛋白样品进行聚丙烯酰胺凝胶电泳。采用半干转的方法进行转膜,25 V电压转膜20—30 min。5 %脱脂牛奶,NC膜正面朝上室温封闭1 h。根据目的蛋白所带的标签不同,选择不同的抗体,将封闭后的NC膜加入一抗,4 ℃过夜。二抗室温孵育1 h。使用ECL发光液显色。

表1 本研究所用引物
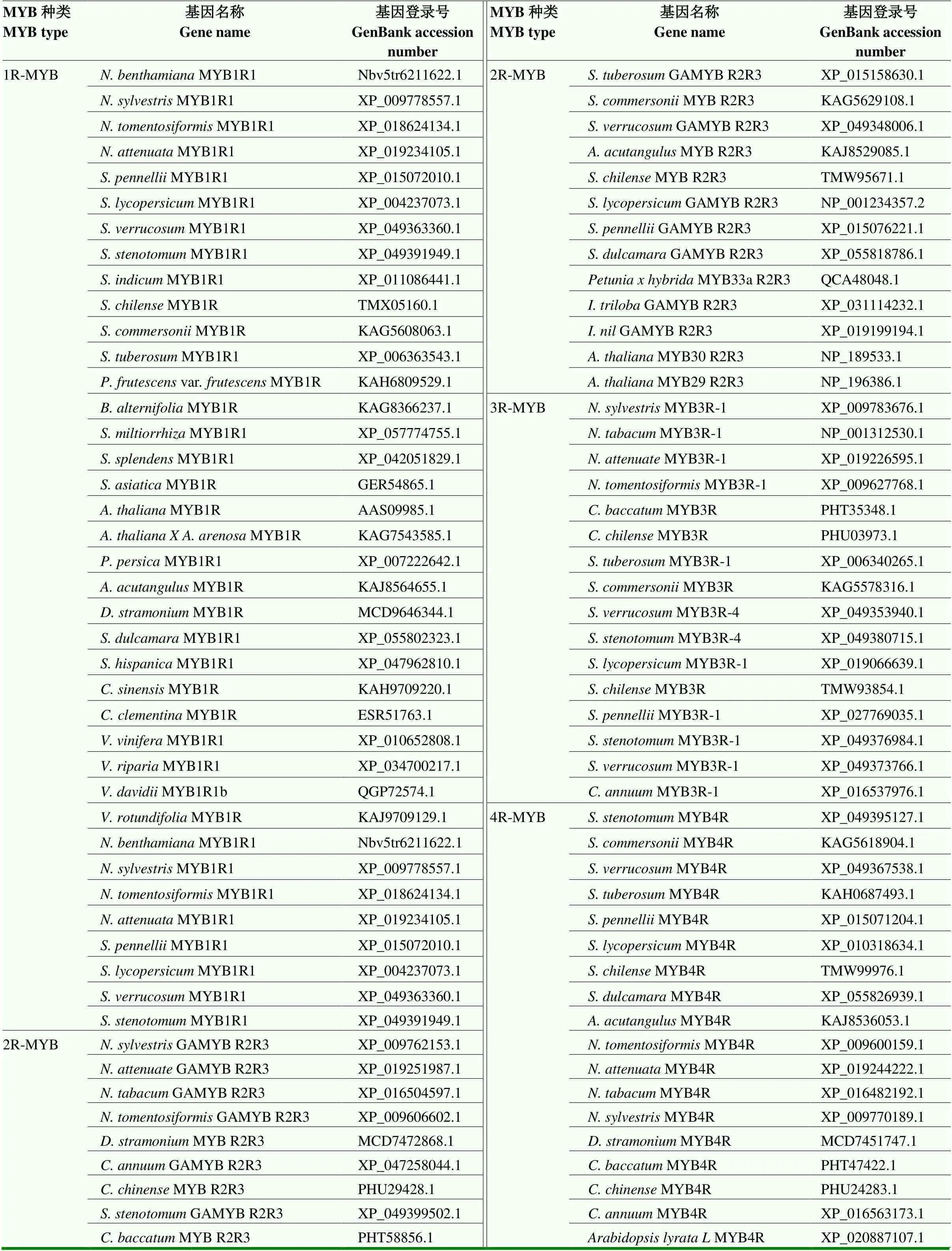
表2 用于建立系统发育树的基因登录号
1.9 台盼蓝染色
台盼蓝染色时,将叶片置于台盼蓝溶液(1 mg·mL-1)中25 ℃黑暗浸泡12 h。反应结束后,将叶片置于沸水3 min,静置2 min,重复两次以去除多余的台盼蓝染料。冷却后,将叶片置于水合氯醛(1 mg·mL-1)2 d去除叶绿素。
1.10 DAB染色
DAB染色时,将叶片置于DAB(2 mg·mL-1)溶于Tris-HCl(pH 3.8)中,25 ℃黑暗浸泡12 h。反应结束后,将叶片置于沸腾的95%乙醇中去除叶绿素。冷却后,将叶片置于75%乙醇中2—3 h。观察其产生的棕黄色物质即为产生的H2O2量。
1.11 酵母双杂交试验
利用同源重组的方法将载体pGBKT7与CP、MP、129K、MET、HEL和P2 PCR产物,载体pGADT7与PCR产物连接。将pGADT7-NbMYB1R1+ pGBKT7-CP、pGADT7-NbMYB1R1+pGBKT7-MP、pGADT7-NbMYB1R1+pGBKT7-129K、pGADT7- NbMYB1R1+pGBKT7-MET、pGADT7-NbMYB1R1+ pGBKT7-HEL和pGADT7-NbMYB1R1+pGBKT7-P2载体共转化酵母感受态Y2H,以pGADT7-T+ pGBKT7- 53为阳性对照,pGADT7-T+pGBKT7-Lam为阴性对照。将上述组合在二缺固体培养基SD-L-T和SD-L-T- H-Ade平板上梯度稀释培养。观察SD-L-T-H-Ade平板上菌落是否变蓝证明是否互作。
1.12 数据处理与分析
每项试验设置至少3个生物学重复,采用T测验进行数据分析。数据均用平均值±标准差表示。采用GraphPad Prism 8.0.2软件制作柱形图和折线图。
2 结果
2.1 本氏烟NbMYB1R1同源进化树分析
同源进化树分析是鉴定转录因子蛋白功能的一种方法,聚在同一支上的同源蛋白结构相似,可能具有相似的功能。对NbMYB1R1进行同源进化树分析,由图1可知,NbMYB1R1属于MYB分类中的1R MYB大类并与多种烟草的MYB转录因子相聚成簇,与拟南芥和辣椒中的MYB1R1转录因子亲缘关系较远。
2.2 NbMYB1R1转录因子亚细胞定位
构建了NbMYB1R1-GFP,通过共聚焦显微镜观察NbMYB1R1亚细胞定位。结果显示,对照GFP分布在整个细胞中,而MYB1R1-GFP与H2B-RFP共定位于细胞核中(图2)。推测NbMYB1R1在细胞核中作为转录因子调控下游基因的转录,进而影响病毒的侵染。
2.3 CGMMV侵染对NbMYB1R1转录水平的影响
CGMMV侵染植株在8 d时出现症状,因此取8、10和12 d的发病植株的系统叶为处理组,健康植株的系统叶为对照组,提取RNA并反转录。qRT-PCR结果显示,在CGMMV侵染8、12 d时,的转录水平显著上调;10 d转录水平与健康植株无显著差异。表明CGMMV侵染初期及中后期会诱导的转录表达(图3)。
2.4 NbMYB1R1沉默促进CGMMV的侵染
为了明确在CGMMV侵染中的作用,通过VIGS技术研究沉默后对CGMMV侵染的影响。将TRV-RNA2-与TRV-RNA1农杆菌菌液等体积混合后,浸润本氏烟进行基因沉默,标记为TRV:;TRV-RNA1菌液与TRV-RNA2菌液混合后浸润本氏烟作为对照组,标记为TRV:00。基因沉默10 d后,观察发现与对照植株相比,沉默后植株无表型变化(图4-A)。检测沉默效率为81%(图4-B)。
对沉默10 d的TRV:和TRV:00植株进行摩擦接种CGMMV。第3.5天TRV:植株开始发病,第4天TRV:00植株开始发病,因此每0.5 d观察并统计两组处理的发病植株,发现从第3—6天,TRV:组的发病植株数量始终多于TRV:00(图4-C)。第6天,沉默植株系统叶上病毒症状明显较重(图4-A)。利用qRT-PCR和Western blot检测病毒积累量,沉默植株的CGMMV CP无论在转录水平(图4-D)还是蛋白水平(图4-E),其积累量均显著高于对照植株。证明沉默促进了CGMMV CP积累,即促进CGMMV的侵染。
2.5 NbMYB1R1瞬时过表达抑制CGMMV侵染
为进一步明确NbMYB1R1对CGMMV侵染的影响,在CGMMV侵染的叶片上浸润NbMYB1R1-GFP,并以GFP为对照(OD600=1.0),浸润在同一叶片的左右两侧,72 h后,在激光共聚焦显微镜下观察绿色荧光均正确表达后进行取样。western blot结果显示,NbMYB1R1-GFP和GFP均正常表达,与对照相比,NbMYB1R1-GFP过表达后的CGMMV CP积累量较少(图5-A),表明NbMYB1R1瞬时过表达抑制CGMMV的侵染。对NbMYB1R1关键结构域进行预测,其中121—127位氨基酸为MYB DNA结合结构域,在CGMMV侵染的叶片上浸润NbMYB1R1-GFP和NbMYB1R1121-127-GFP,并以GFP为对照(OD600= 1.0),72 h后western blot结果表明,NbMYB1R1-GFP关键氨基酸缺失后会缓解NbMYB1R1对CGMMV的抑制作用(图5-B)。

图1 基于氨基酸序列的NbMYB1R1进化树分析
2.6 NbMYB1R1促进ROS积累及细胞坏死
NbMYB1R1瞬时过表达叶片出现细胞坏死症状,推测由于NbMYB1R1表达调控ROS的积累导致叶片坏死。对NbMYB1R1瞬时过表达叶片进行了台盼蓝和DAB染色。结果显示,NbMYB1R1-GFP的浸润区域出现坏死,台盼蓝染色显示蓝色;DAB染色后会有棕黄色物质积累(图6),表明NbMYB1R1过表达会诱导ROS积累并出现细胞坏死的症状。
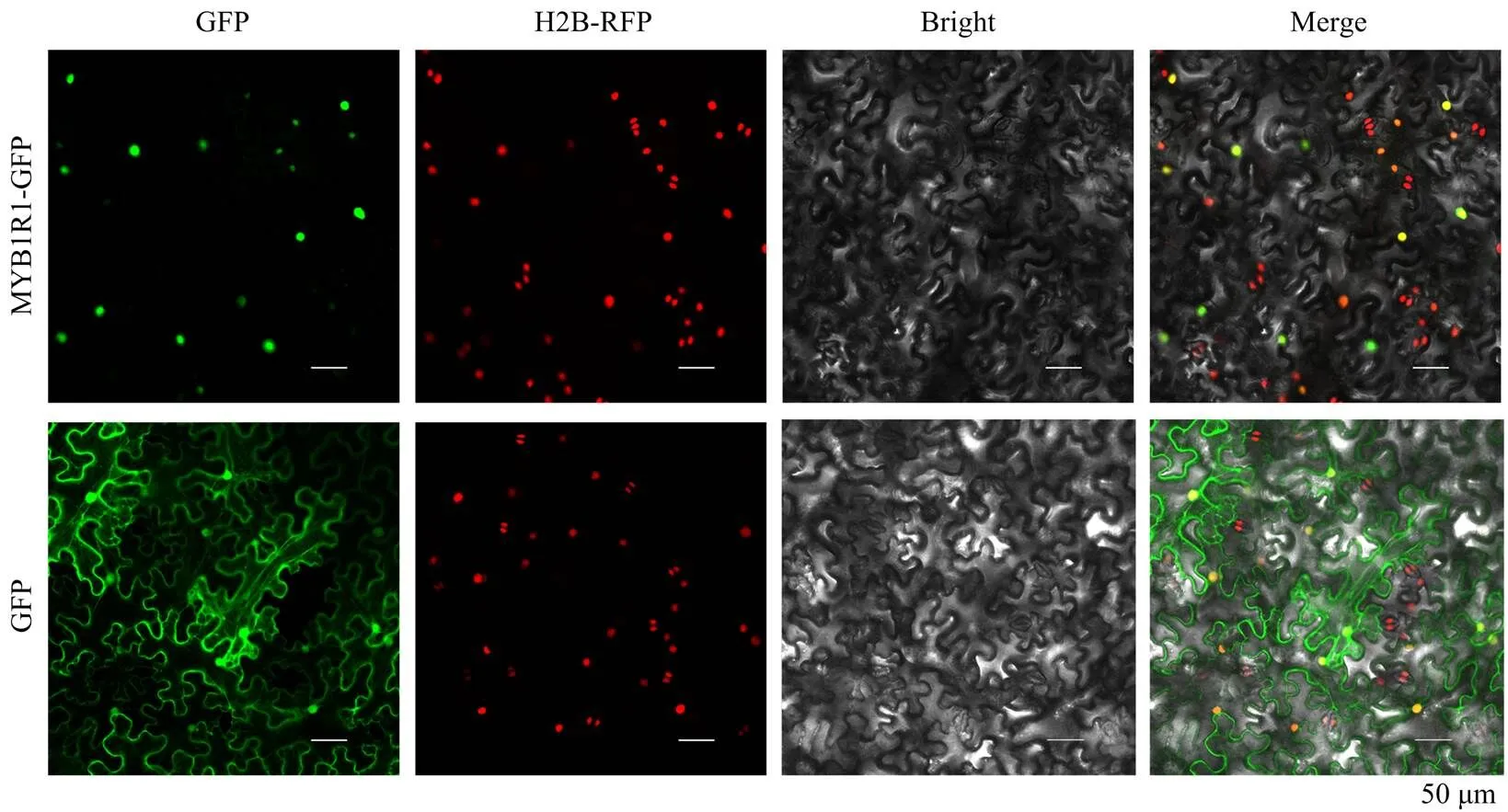
图2 NbMYBIR1的亚细胞定位

*:差异显著significant difference (P<0.05)。下同The same as below
2.7 沉默NbMYB1R1激活了交替氧化酶基因NbAOX1a和NbAOX1b的转录
为探究如何调控ROS的积累,检测了沉默植株中ROS负调控相关基因的转录水平,结果显示交替氧化酶基因和转录水平显著上调,而其余ROS清除相关基因、、、和[37]的转录水平无显著变化(图7)。上述结果表明可特异性调控交替氧化酶基因和,推测可能通过抑制和的转录从而促进ROS积累。
2.8 NbAOX1a和NbAOX1b沉默抑制CGMMV的侵染
为了明确和在CGMMV侵染中的作用,通过VIGS技术研究沉默和对CGMMV侵染的影响。将TRV-RNA2-/与TRV-RNA1农杆菌菌液等体积混合后,浸润本氏烟进行基因沉默,标记为TRV:/;TRV-RNA1菌液与TRV-RNA2菌液混合后浸润本氏烟作为对照组,标记为TRV:00。基因沉默10 d后,发现与对照植株相比,/沉默后植株有很明显的矮化(图8-A)。和的沉默效率分别为71%和64%(图8-B)。
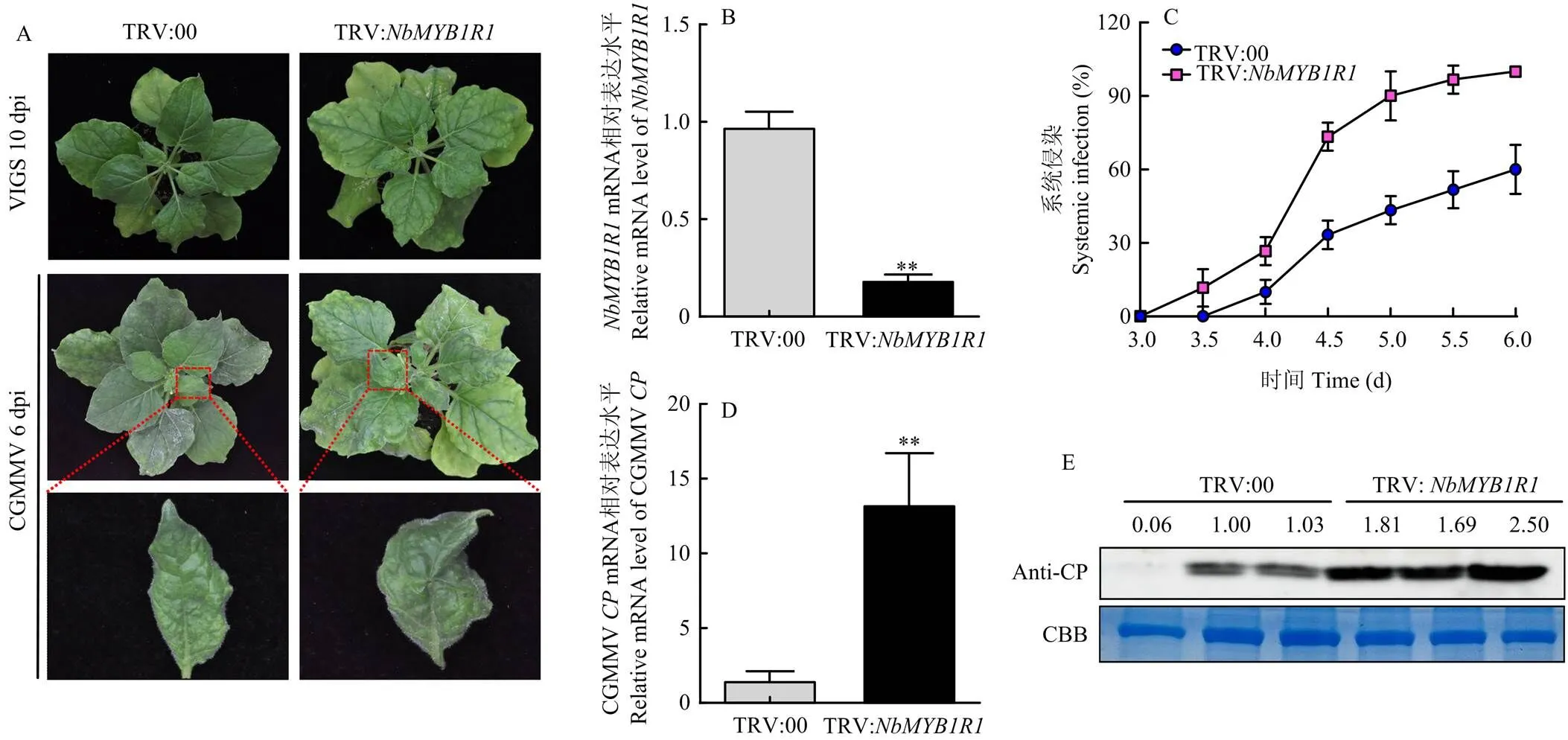
A:TRV:NbMYB1R1和TRV:00在CGMMV侵染前后的生物学表型Biological phenotypes of TRV:NbMYBIR1 and TRV:00 before and after CGMMV infection;B:VIGS NbMYBIR1的沉默效率Silencing efficiency of VIGS NbMYBIR1;C:TRV:NbMYBIR1和TRV:00的植株被CGMMV侵染后系统侵染植株占比Proportion of systemic infected plants after infection with CGMMV in plants of TRV:NbMYBIR1 and TRV:00;D:TRV:NbMYBIR1对CGMMV CP转录水平的影响Effect of TRV:NbMYBIR1 on the transcription level of CGMMV CP;E:TRV:NbMYBIR1对CGMMV CP蛋白水平的影响 Effect of TRV:NbMYBIR1 on the protein level of CGMMV CP

A:NbMYB1R1瞬时过表达对CGMMV CP蛋白水平的影响Effect of transient overexpression of NbMYB1R1 on the protein level of CGMMV CP;B:NbMYB1R1 DNA结合结构域缺失突变对CGMMV CP蛋白水平的影响Effect of NbMYB1R1 DNA binding domain deletion mutation on the protein level of CGMMV CP
对沉默10 d的TRV:/和TRV:00植株进行摩擦接种CGMMV。第4天TRV:00植株开始发病,第4.5天TRV:/植株开始发病,因此每0.5 d观察并统计两组处理的发病植株,发现从第3.5—7天,TRV:00组的发病植株数量始终多于TRV:/(图8-C)。第7天,/沉默植株系统叶上病毒症状明显较轻(图8-A)。利用qRT-PCR和Western blot检测病毒积累量,/沉默植株的CGMMVCP无论在转录水平(图8-D)还是蛋白水平(图8-E),其积累量都比对照植株显著降低。证明/沉默减少了CGMMV CP积累,即抑制CGMMV的侵染。

图6 NbMYB1R1过表达引起活性氧积累与细胞死亡
2.9 NbMYB1R1不与CGMMV编码蛋白互作
为了解NbMYB1R1是否与CGMMV编码蛋白之间存在互作关系,进而直接影响CGMMV的侵染,利用酵母双杂交系统验证NbMYB1R1与CGMMV相关蛋白之间的互作。构建重组质粒pGADT7-NbMYB1R1、pGBKT7-CP、pGBKT7-MP、pGBKT7-129K、pGBKT7- MET、pGBKT7-HEL和pGBKT7-P2。将pGADT7- NbMYB1R1+pGBKT7-CP、pGADT7-NbMYB1R1+ pGBKT7-MP、pGADT7-NbMYB1R1+pGBKT7-129K、pGADT7-NbMYB1R1+pGBKT7-MET、pGADT7- NbMYB1R1+pGBKT7-HEL和pGADT7-NbMYB1R1+ pGBKT7-P2载体共转化酵母感受态Y2H,以pGADT7- T+pGBKT7-53为阳性对照,pGADT7-T+pGBKT7-Lam为阴性对照。将上述组合涂布于二缺培养基。3 d后,所有组合均能在二缺培养基上正常生长,说明质粒成功转入酵母菌株。将上述组合在二缺固体培养基SD-L-T和SD-L-T-H-Ade平板上梯度稀释培养。结果显示,仅pGADT7-T+pGBKT7-53阳性对照生长且变蓝色,其余包含NbMYB1R1组合在SD-L-T-H-Ade平板上生长的菌落均未显现蓝色(图9),表明酵母双杂交试验中NbMYB1R1不与CGMMV编码蛋白直接互作。
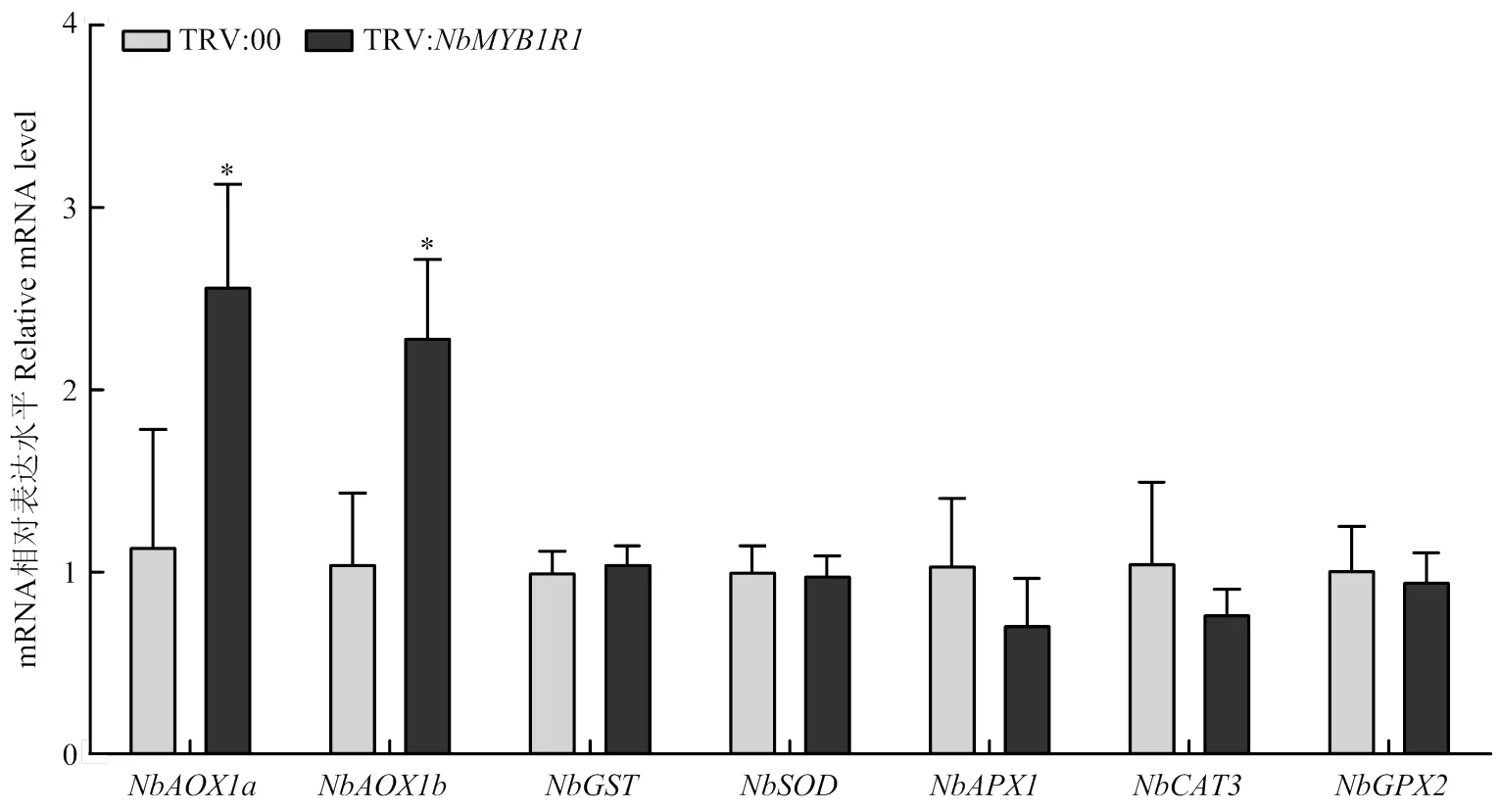
图7 NbMYBIR1沉默对ROS相关基因转录水平的影响
3 讨论
3.1 NbMYB1R1作为转录因子调控病毒侵染
MYB转录因子在调控植物生长发育过程和其对生物及非生物胁迫的反应等方面发挥着重要作用[38]。自1987年在玉米中首次发现植物MYB转录因子(ZmMYBC1)后[4],越来越多的研究表明MYB作为转录因子在植物生物胁迫和非生物胁迫中发挥重要作用[39]。CiMYB4作为转录因子可以显著降低镉胁迫对烟草和野菊生长发育的影响[40];过表达转录因子VvMYB30可增强转基因拟南芥的耐盐性[41];转录因子CitMYB20可以增强柑橘株系对柑橘溃疡病的抗性[42];NtMYB1可能参与病程相关(PR)基因的转录激活进而增强烟草对于TMV的抗性[24];番茄黄化曲叶病毒侵染植株后,SlMYB28作为转录因子激活多个防御相关基因的表达进而增强植株自身抗病能力[26]。前期研究表明,CGMMV侵染上调本氏烟表达[43],进一步探索发现在CGMMV侵染初期及中后期,其转录水平明显上调(图3)。过表达和沉默均能对CGMMV的侵染产生影响(图4、图5),且NbMYB1R1中DNA结合结构域是影响CGMMV侵染的关键结构域(图5-B)。利用其氨基酸序列进行系统发育分析,表明NbMYB1R1与多种烟草中的MYB1R1具有高度相似性(图1),且NbMYB1R1主要定位于烟草细胞核(图2)。因此,笔者推测NbMYB1R1可能作为转录因子参与病毒的侵染过程。

A:TRV:NbAOX1a/b和TRV:00在CGMMV侵染前后的生物学表型Biological phenotypes of TRV:NbAOX1a/b and TRV:00 before and after CGMMV infection;B:VIGS NbAOX1a和NbAOX1b的沉默效率Silencing efficiency of VIGS NbAOX1a and NbAOX1b;C:TRV:NbAOX1a/b和TRV:00的植株被CGMMV侵染后系统侵染植株占比Proportion of systemic infected plants after infection with CGMMV in plants of TRV:NbAOX1a/b and TRV:00;D:TRV:NbAOX1a/b对CGMMV CP的转录水平影响Effect of TRV: NbAOX1a/b on the transcription level of CGMMV CP;E:TRV:NbAOX1a/b对CGMMV CP蛋白水平的影响Effect of TRV:NbAOX1a/b on the protein level of CGMMV CP

图9 酵母双杂交分析NbMYB1R1与CGMMV编码蛋白的互作
3.2 NbMYB1R1通过抑制NbAOX1a和NbAOX1b相关基因的转录激活细胞内ROS积累从而抑制病毒侵染
细胞内ROS信号转导或ROS激活的防御机制可抑制病原物侵染。例如,在植株中沉默内源基因后可以增强寄主植物活性氧积累从而提高植株抗病能力[44];可能通过调控植物体内活性氧产生诱导病程相关蛋白基因的表达,从而提高植株对赤霉菌的抗性[45];H2O2参与StRab5b介导的对马铃薯疫霉的抗病过程[46];柑橘衰退病毒(citrustristeza virus,CTV)侵染会引发本氏烟和天然宿主中ROS爆发,进而增强植物对CTV的抗性。本研究结果也显示NbMYB1R1增强细胞内ROS积累导致细胞坏死从而抑制病毒侵染。同时检测了ROS清除酶相关的基因交替氧化酶(和)、超氧化物歧化酶()、抗坏血酸过氧化物酶()、过氧化氢酶()、谷胱甘肽过氧化物酶()和谷胱甘肽转移酶()[37]。其中和活性可以降低和NO的产生,从而减少细胞内ROS的产生[47]。本研究结果显示沉默可以特异性地上调和的转录(图7)。已经有研究证明MYB转录因子可以与AOX基因的启动子相结合进而影响植株的抗性。转录因子MYB29是的负调节因子,MYB29通过抑制的表达增强植株的胁迫防御反应[48];MYB30结合的启动子以调节植物耐盐性[49]。本研究结果显示沉默和能抑制CGMMV的侵染(图8)。推测NbMYB1R1作为转录因子与和的启动子相结合,通过抑制和相关基因的转录促进细胞内ROS积累从而抑制病毒侵染。
4 结论
NbMYB1R1作为1R MYB家族转录因子在细胞核中行使功能,其在CGMMV侵染初期及中后期表达量显著上调。NbMYB1R1在CGMMV侵染过程中发挥重要作用,其可能原因是NbMYB1R1作为转录因子抑制和的表达,从而促进细胞内ROS积累,抑制了病毒的积累。
[1] 赵文. 黄瓜栽培的现状及其发展趋势. 智慧农业导刊, 2022, 2(3): 32-34.
ZHAO W. The current situation and development trends of cucumber cultivation. Journal of Smart Agriculture, 2022, 2(3): 32-34. (in Chinese)
[2] LING K S, LI R, ZHANG W. First report of cucumber green mottle mosaic virus infecting greenhouse cucumber in Canada. Plant Disease, 2014, 98(5): 701.
[3] KLEMPNAUER K H, GONDA T J, BISHOP J M. Nucleotide sequence of the retroviral leukemia gene v-myb and its cellular progenitor c-myb: the architecture of a transduced oncogene. Cell, 1982, 31(1): 453-463.
[4] PAZ-ARES J, GHOSAL D, WIENAND U, PETERSON P A, SAEDLER H. The regulatory c1 locus ofencodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators. The Embo Journal, 1987, 6(12): 3553-3558.
[5] KATIYAR A, SMITA S, LENKA S K, RAJWANSHI R, CHINNUSAMY V, BANSAL K C. Genome-wide classification and expression analysis of MYB transcription factor families in rice and. BMC Genomics, 2012, 13: 544.
[6] SALIH H, GONG W, HE S, SUN G, SUN J, DU X. Genome- wide characterization and expression analysis of MYB transcription factors in. BMC Genetics, 2016, 17(1): 129.
[7] WEI Q, CHEN R, WEI X, LIU Y, ZHAO S, YIN X, XIE T. Genome- wide identification of R2R3-MYB family in wheat and functional characteristics of the abiotic stress responsive gene. BMC Genomics, 2020, 21(1): 792.
[8] ZHAO P, LI Q, LI J, WANG L, REN Z. Genome-wide identification and characterization of R2R3MYB family in. Molecular Genetics and Genomics, 2014, 289(6): 1183-1207.
[9] CHEN G, HE W, GUO X, PAN J. Genome-wide identification, classification and expression analysis of the myb transcription factor family in. International Journal of Molecular Sciences, 2021, 22(9): 4838.
[10] OGATA K, MORIKAWA S, NAKAMURA H, SEKIKAWA A, INOUE T, KANAI H, SARAI A, ISHII S, NISHIMURA Y. Solution structure of a specific DNA complex of the Myb DNA-binding domain with cooperative recognition helices. Cell, 1994, 79(4): 639-648.
[11] DUBOS C, STRACKE R, GROTEWOLD E, WEISSHAAR B, MARTIN C, LEPINIEC L. MYB transcription factors in. Trends in Plant Science, 2010, 15(10): 573-581.
[12] YU E Y, KIM S E, KIM J H, KO J H, CHO M H, CHUNG I K. Sequence-specific DNA recognition by the Myb-like domain of plant telomeric protein RTBP1. The Journal of Biological Chemistry, 2000, 275(31): 24208-24214.
[13] HAGA N, KATO K, MURASE M, ARAKI S, KUBO M, DEMURA T, SUZUKI K, MÜLLER I, VOß U, JÜRGENS G, ITO M. R1R2R3-Myb proteins positively regulate cytokinesis through activation of KNOLLE transcription in. Development, 2007, 134(6): 1101-1110.
[14] HOU X J, LI S B, LIU S R, HU C G, ZHANG J Z. Genome-wide classification and evolutionary and expression analyses of citrus MYB transcription factor families in sweet orange. PLoS One, 2014, 9(11): e112375.
[15] MATUS J T, AQUEA F, ARCE-JOHNSON P. Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization acrossandgenomes. BMC Plant Biology, 2008, 8(1): 83.
[16] WILKINS O, NAHAL H, FOONG J, PROVART N J, CAMPBELL M M. Expansion and diversification of theR2R3-MYB family of transcription factors. Plant Physiology, 2009, 149(2): 981-993.
[17] LIU Z, LUAN Y, LI J, YIN Y. Expression of a tomato MYB gene in transgenic tobacco increases resistance toand. European Journal of Plant Pathology, 2016, 144(3): 607-617.
[18] LIU T, CHEN T, KAN J, YAO Y, GUO D, YANG Y, LING X, WANG J, ZHANG B. The GhMYB36 transcription factor confers resistance to biotic and abiotic stress by enhancing PR1 gene expression in plants. Plant Biotechnology Journal, 2022, 20(4): 722-735.
[19] XIAO S, HU Q, SHEN J, LIU S, YANG Z, CHEN K, KLOSTERMAN S J, JAVORNIK B, ZHANG X, ZHU L. GhMYB4 downregulates lignin biosynthesis and enhances cotton resistance to. Plant Cell Reports, 2021, 40(4): 735-751.
[20] QIU B, CHEN H, ZHENG L, SU L, CUI X, GE F, LIU D. An MYB transcription factor modulatesresistance against the root rot pathogenby regulating the jasmonate acid signaling pathway and photosynthesis. Phytopathology, 2022, 112(6): 1323-1334.
[21] CUI J, JIANG N, ZHOU X, HOU X, YANG G, MENG J, LUAN Y. Tomato MYB49 enhances resistance toand tolerance to water deficit and salt stress. Planta, 2018, 248(6): 1487-1503.
[22] QIU Z, YAN S, XIA B, JIANG J, YU B, LEI J, CHEN C, CHEN L, YANG Y, WANG Y, TIAN S, CAO B. The eggplant transcription factor MYB44 enhances resistance to bacterial wilt by activating the expression of spermidine synthase. Journal of Experimental Botany, 2019, 70(19): 5343-5354.
[23] LIU R, CHEN L, JIA Z, Lü B, SHI H, SHAO W, DONG H. Transcription factor AtMYB44 regulates induced expression of the ETHYLENE INSENSITIVE2 gene inresponding to a harpin protein. Molecular Plant-Microbe Interactions, 2011, 24(3): 377-389.
[24] YANG Y, KLESSIG D F. Isolation and characterization of a tobacco mosaic virus-inducible myb oncogene homolog from tobacco. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(25): 14972-14977.
[25] ZHU T, ZHOU X, ZHANG J L, ZHANG W H, ZHANG L P, YOU C X, JAMESON P E, MA P T, GUO S L. Ethylene-induced NbMYB4L is involved in resistance against tobacco mosaic virus in. Molecular Plant Pathology, 2022, 23(1): 16-31.
[26] ZHOU H, PENG Q, ZHAO J, OWITI A, REN F, LIAO L, WANG L, DENG X, JIANG Q, HAN Y. Multiple R2R3-MYB transcription factors involved in the regulation of anthocyanin accumulation in peach flower. Frontiers in Plant Science, 2016, 7: 1557.
[27] KOUNDAL V, VINUTHA T, HAQ Q M R, PRAVEEN S. Modulation of plant development and MYB down regulation: both duringexpression ofand in natural ToLCV infection. Journal of Plant Biochemistry and Biotechnology, 2010, 19: 171-175.
[28] SEO P J, XIANG F, QIAO M, PARK J Y, LEE Y N, KIM S G, LEE Y H, PARK W J, PARK C M. The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in. Plant Physiology, 2009, 151(1): 275-289.
[29] SLAVOKHOTOVA A, KOROSTYLEVA T, SHELENKOV A, PUKHALSKIY V, KOROTTSEVA I, SLEZINA M, ISTOMINA E, ODINTSOVA T. Transcriptomic analysis of genes involved in plant defense response to the cucumber green mottle mosaic virus infection. Life, 2021, 11(10): 1064.
[30] EULGEM T. Regulation of thedefense transcriptome. Trends in Plant Science, 2005, 10(2): 71-78.
[31] YU Y, ZHANG S, YU Y, CUI N, YU G, ZHAO H, MENG X, FAN H. The pivotal role of MYB transcription factors in plant disease resistance. Planta, 2023, 258(1): 16.
[32] VAILLEAU F, DANIEL X, TRONCHET M, MONTILLET J L, TRIANTAPHYLIDèS C, ROBY D. A R2R3-MYB gene,, acts as a positive regulator of the hypersensitive cell death program in plants in response to pathogen attack. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(15): 10179-10184.
[33] RAFFAELE S, VAILLEAU F, LéGER A, JOUBèS J, MIERSCH O, HUARD C, BLéE E, MONGRAND S, DOMERGUE F, ROBY D. A MYB transcription factor regulates very-long-chain fatty acid biosynthesis for activation of the hypersensitive cell death response in. The Plant Cell, 2008, 20(3): 752-767.
[34] NOMAN A, HUSSAIN A, ADNAN M, KHAN M I, ASHRAF M F, ZAINAB M, KHAN K A, GHRAMH H A, HE S. A novel MYB transcription factor CaPHL8 provide clues about evolution of pepper immunity againstsoil borne pathogen. Microbial Pathogenesis, 2019, 137: 103758.
[35] YAO L, YANG B, XIAN B, CHEN B, YAN J, CHEN Q, GAO S, ZHAO P, HAN F, XU J, JIANG Y Q. The R2R3-MYB transcription factor BnaMYB111L from rapeseed modulates reactive oxygen species accumulation and hypersensitive-like cell death. Plant Physiology and Biochemistry, 2020, 147: 280-288.
[36] DANEVA A, GAO Z, VAN DURME M, NOWACK M K. Functions and regulation of programmed cell death in plant development. Annual Review of Cell and Developmental Biology, 2016, 32: 441-468.
[37] CHEN B, NIU F, LIU W Z, YANG B, ZHANG J, MA J, CHENG H, HAN F, JIANG Y Q. Identification, cloning and characterization of R2R3-MYB gene family in canola (L.) identify a novel member modulating ROS accumulation and hypersensitive-like cell death. DNA Research, 2016, 23(2): 101-114.
[38] NG D W, ABEYSINGHE J K, KAMALI M. Regulating the regulators: The control of transcription factors in plant defense signaling. International Journal of Molecular Sciences, 2018, 19(12): 3737.
[39] VISWANATH K K, KUO S Y, TU C W, HSU Y H, HUANG Y W, HU C C. The role of plant transcription factors in the fight against plant viruses. International Journal of Molecular Sciences, 2023, 24(9): 8433.
[40] 李强, 康璠, 薛晴, 陈斌, 孙颖. 神农香菊R2R3-MYB转录因子CiMYB4在镉胁迫中的功能分析. 草业学报, 2024, 33(5): 128-142.
LI Q, KANG F, XUE Q, CHEN B, SUN Y. Functional analysis of the R2R3-MYB transcription factor CiMYB4 ofvar.in response to cadmium stress. Acta Prataculturae Sinica, 2024, 33(5): 128-142. (in Chinese)
[41] 解振强, 许桓瑜, 黄金霞, 蔡善亚, 李刚, 赵鹏程. 葡萄MYB转录因子基因的克隆及其耐盐性分析. 中外葡萄与葡萄酒, 2024(1): 20-27.
XIE Z Q, XU H Y, HUANG J X, CAI S Y, LI G, ZHAO P C. Cloning and salt tolerance analysis offrom grape (). Sino-Overseas Grapevine & Wine, 2024(1): 20-27. (in Chinese)
[42] 姚利晓, 范海芳, 张庆雯, 何永睿, 许兰珍, 雷天刚, 彭爱红, 李强, 邹修平, 陈善春. 柑橘溃疡病抗性相关转录因子CitMYB20的功能. 中国农业科学, 2020, 53(10): 1997-2008. doi: 10.3864/j.issn.0578- 1752.2020.10.007.
YAO L X, FAN H F, ZHANG Q W, HE Y R, XU L Z, LEI T G, PENG A H, LI Q, ZOU X P, CHEN S C. Function of citrus bacterial canker resistance-related transcription factor CitMYB20. Scientia Agricultura Sinica, 2020, 53(10): 1997-2008. doi: 10.3864/j.issn.0578-1752.2020. 10.007. (in Chinese)
[43] YANG L L, LI Q L, HAN X Y, JIANG X L, WANG H, SHI Y J, CHEN L L, LI H L, LIU Y Q, YANG X, SHI Y. A cysteine-rich secretory protein involves in phytohormone melatonin mediated plant resistance to CGMMV. BMC Plant Biology, 2023, 23(1): 215.
[44] LI Y, ZHANG R, WU Y, WU Q, JIANG Q, MA J, ZHANG Y, QI P, CHEN G, JIANG Y, ZHENG Y, WEI Y, XU Q. TaRBP1 stabilizes TaGLTP and negatively regulates stripe rust resistance in wheat. Molecular Plant Pathology, 2023, 24(10): 1205-1219.
[45] 陈同睿, 罗艳君, 赵潘婷, 贾海燕, 马正强. 过表达基因提高了小麦赤霉病抗性. 作物学报, 2021, 47(1): 19-29.
CHEN T R, LUO Y J, ZHAO P T, JIA H Y, MA Z Q. Overexpression ofenhances the Fusarium head blight resistance in wheat. Acta Agronomica Sinica, 2021, 47(1): 19-29. (in Chinese)
[46] TIAN Z, ZHANG Z, KANG L, LI M, ZHANG J, FENG Y, YIN J, GONG X, ZHAO J. Small G protein StRab5b positively regulates potato resistance to. Frontiers in Plant Science, 2022, 13: 1065627.
[47] CVETKOVSKA M, VANLERBERGHE G C. Alternative oxidase impacts the plant response to biotic stress by influencing the mitochondrial generation of reactive oxygen species. Plant, Cell and Environment, 2013, 36(3): 721-732.
[48] ZHANG X, IVANOVA A, VANDEPOELE K, RADOMILJAC J, VAN DE VELDE J, BERKOWITZ O, WILLEMS P, XU Y, NG S, VAN AKEN O,. The transcription factor myb29 is a regulator of. Plant Physiology, 2017, 173(3): 1824-1843.
[49] GONG Q, LI S, ZHENG Y, DUAN H, XIAO F, ZHUANG Y, HE J, WU G, ZHAO S, ZHOU H, LIN H. SUMOylation of MYB30 enhances salt tolerance by elevating alternative respiration via transcriptionally upregulating AOX1a in. The Plant Journal, 2020, 102(6): 1157-1171.
The Transcription Factor NbMYB1R1 Inhibits Viral Infection by Promoting ROS Accumulation
JIANG XingLin1, YU LianWei1, Fu Han1, Ai Niu1, Cui YingJun5, Li HaoHai6, XIA ZiHao7, Yuan HongXia1, LI HongLian1,3,4, YANG Xue1,2, SHI Yan1
1College of Plant Protection, Henan Agricultural University, Zhengzhou 450002;2Crop science postdoctoral programme of Henan Agricultural University, Zhengzhou 450002;3Collaborative Innovation Centre of Henan Grain crops, Zhengzhou 450002;4State Key Laboratory of Wheat and Maize Crop Science, Zhengzhou 450002;5Henan Association for the promotion of new plant protection techniques, Zhengzhou 450002;6Henan plant protection and quarantine station, Zhengzhou 450002;7College of Plant Protection, Shenyang Agricultural University, Shenyang 110866
【Background】Cucumber green mottle mosaic virus (CGMMV) is an important quarantine plant virus in China, which has seriously reduced the production of vegetables and melons worldwide. The MYB protein family is large, multifunctional and present in all eukaryotes. Most MYB proteins act as transcription factors that control development, and metabolism in plants, and regulate biotic and abiotic stress responses. Previous studies have shown that during CGMMV infection, the expression of transcription factor genewas significantly up-regulated.【Objective】The objective of this study is to clarify the mechanism of NbMYB1R1 involved in CGMMV infection, and to provide a theoretical basis for controlling CGMMV infection.【Method】MEGA 7.0 was used to construct a phylogenetic tree to analyze the amino acid sequence of NbMYB1R1 protein. The expression vector NbMYB1R1-GFP was constructed, transformed intoGV3101 and infiltrated the tobacco leaves to observe the subcellular localization of NbMYB1R1 by confocal microscopy. The transcriptional levels ofat different stages of CGMMV infection and ROS related genes in silencedplants were analyzed using qRT-PCR technology. TRV-mediated gene silencing (VIGS) was utilized to analyze the function of,, andduring CGMMV infection. transient overexpression of NbMYB1R1 and NbMYB1R1 mutant was conducted to analyze the effect of NbMYB1R1 on CGMMV infection. trypan blue staining and DAB stainingwereused toobserve whether the cell death caused by the transient overexpression of NbMYB1R1 was related to the programmed cell death (PCD) and the accumulation of ROS. Yeast two-hybrid system was used to verify whether NbMYB1R1 interacted with CGMMV related proteins.【Result】Phylogenetic tree analysis showed that NbMYB1R1 belonged to the 1R MYB category and had high homology with MYB transcription factors in a variety of tobaccos. The results of subcellular localization showed that NbMYB1R1 was localized in the nucleus. In CGMMV-infected tobacco plants, the transcript level ofwas significantly changed compared with healthy plants, which was significantly up-regulated at 8 and 12 d of CGMMV infection. CGMMV was inoculated into TRV:and TRV:00 plants. The-silenced plants showed the mottled and curled symptoms at 3 dpi, while the control plant appeared the symptoms at 3.5 dpi. Silencingcould effectively promote the accumulation of CGMMV at mRNA and protein levels. the CGMMV accumulation in NbMYB1R1 and NbMYB1R1 mutant transiently overexpression leaves was detected, and the results showed that NbMYB1R1 overexpression could effectively inhibit CGMMV infection and DNA binding domain deletion mutant reduced the inhibition of CGMMV by NbMYB1R1. The results of trypan blue and DAB staining showed that NbMYB1R1 overexpression could induce the ROS accumulation and cell death. the transcription level of ROS-related genes in TRV:and TRV:00 plant was also detected, and the results showed that silencingcould specifically up-regulate the transcription ofand. After CGMMV inoculation on silencing endogenous genesand, the systemic leaves of silencedandplants showed mottled and curled symptoms at 4 dpi, while the control plants appeared symptoms at 3.5 d. Meanwhile, the results of CGMMV CP mRNA and protein levels also indicated that silencingandcould effectively inhibit the accumulation of CGMMV. The yeast two-hybridresults showed that NbMYB1R1 did not directly interact with the CGMMV proteins.【Conclusion】During the CGMMV infection, the defense-related geneis up-regulated. It speculates that the upregulation ofinhibits the transcription of the downstream genesandand activates the production of ROS in cells, thereby inhibiting viral infection. However, NbMYB1R1 does not produce this effect through a direct interaction with the CGMMV viral protein. NbMYB1R1 plays an important role in CGMMV infection.
cucumber green mottle mosaic virus (CGMMV);; transcription factor; pathogenic mechanism; reactive oxygen species (ROS)

10.3864/j.issn.0578-1752.2024.08.006
2023-12-06;
2024-02-04
国家自然科学基金(3210170372)、河南农业大学科技创新项目(KJCX2021A13)
姜兴林,E-mail:1442295312@qq.com。于连伟,E-mail:ylwnet972@163.com。姜兴林和于连伟为同等贡献作者。通信作者杨雪,E-mail:yangxuepphappy@126.com。通信作者施艳,E-mail:shiyan00925@126.com
(责任编辑 岳梅)
