肠道菌群对抗性淀粉的降解机制
2024-02-23王扶香莫剑慧胡新中
王扶香,莫剑慧,胡新中,马 蓁
(陕西师范大学食品工程与营养科学学院 西安 710119)
1 抗性淀粉的分类及其生理功能
根据淀粉消化速率和程度不同,可将淀粉划分为快速消化淀粉(RDS,体外消化条件下前20 min 被水解)、缓慢消化淀粉(SDS,在20~120 min内被水解)和抗性淀粉(RS,120 min 后不被小肠消化水解的部分淀粉,继续进入大肠[1-3],被定植在大肠内的微生物群完全或部分降解为短链脂肪酸等代谢物[4])。根据植物来源和加工方式不同,可将抗性淀粉细分为以下5 类:1)物理包埋淀粉(RS1):指因物理屏障作用而不能被小肠酶接近消化的淀粉,主要存在于部分碾磨的谷物和种子中,豆类是RS1 的主要来源[5]。2)天然抗性淀粉颗粒(RS2):指一些高直链淀粉含量的作物中淀粉形成紧密堆积的颗粒结构[6],限制了消化酶的可及性。RS2 主要存在于青/生香蕉、马铃薯和未加工的豌豆中[7]。3)老化回生的淀粉(RS3):主要是由氢键驱动淀粉链重结晶形成的产物[8],存在于重结晶的非颗粒淀粉或加工食品中。4)化学改性淀粉(RS4):通过不同化学反应,如交联、酯化、醚化等及引入某些新的化学官能团而获得的改性抗性淀粉,如淀粉醚、淀粉脂和交联淀粉[9]。5)直链淀粉-脂肪复合物(RS5):主要指具有链状结构的直链淀粉与脂类形成的单螺旋复合物[2,10-11],也有学者认为可将淀粉-甘油、淀粉-氨基酸、淀粉-多肽、淀粉-蛋白质、淀粉-脂蛋白等自组装的V 型配合物归类为RS5[12]。
抗性淀粉多通过与宿主肠道微生物的相互作用来实现其对宿主的健康益处。抗性淀粉为肠道菌群提供生长基质,肠道菌群通过降解抗性淀粉获取其生长所需的能量[13]。肠道微生物降解抗性淀粉产生的含乙酸、丙酸、丁酸等短链脂肪酸代谢物(Short-chain fatty acids,SCFAs)[14]对肠道微生物有一定的调节作用,这些短链脂肪酸可以促进有益菌群增殖,抑制有害菌群生长[15-16],并维持肠道菌群动态平衡[17]。短链脂肪酸有助于改善机体的炎症和氧化应激状态,强化内脏敏感性,同时促进其它功能[18]。短链脂肪酸可以降低结肠内环境的pH 值并促进矿物质的吸收[15-16];可在不影响正常上皮细胞增殖和分化的情况下,通过诱导结肠癌细胞成熟和凋亡抑制人体结肠癌细胞的增殖,保护结肠健康[19]。此外,短链脂肪酸在降低血浆胆固醇浓度和减少心血管疾病发生的风险方面也发挥着一定的作用[20-21]。抗性淀粉发酵过程中,可促进肠道蠕动,增加饱腹感,减少能量摄入[22],促进脂质代谢[15]。抗性淀粉在人体内缓慢代谢有助于降低宿主血糖水平,降低糖尿病和肥胖症等疾病发生的风险[23]。据Li 等[24]报道,在紫山药抗性淀粉对高脂血症仓鼠的干预实验中,抗性淀粉的摄入可降低脂肪酸的合成,增加脂肪酸氧化和甘油-磷脂的合成,从而使仓鼠体质量下降;摄入抗性淀粉刺激肠道微生物产生更高水平的短链脂肪酸,影响体内脂质代谢作用,抑制脂肪组织扩张,降低肝脏、肾脏质量,修复高脂饮食对盲肠和结肠组织形态学造成的损伤。同时,高剂量抗性淀粉饮食的仓鼠体内高密度脂蛋白胆固醇浓度增加,甘油三酯、总胆固醇和低密度脂蛋白胆固醇浓度减少。抗性淀粉能增加肠道胰岛素及胰高血糖素样肽含量,减轻炎症反应,改善运动协调性[25]。抗性淀粉可有效改善Ⅱ型糖尿病模型大鼠紊乱的代谢环境,缓解Ⅱ型糖尿病大鼠的病理损伤[26]。在凡纳滨对虾的日粮中添加抗性淀粉可改善其肠黏膜形态,同时提高其消化酶(包括淀粉酶、脂肪酶、胰蛋白酶和胃蛋白酶)和免疫酶(包括酚氧化酶、总抗氧化能力、总一氧化氮合酶和一氧化氮)的活性,以及部分与免疫相关的基因的表达水平(包括酚氧化酶原、抗脂多糖因子、溶菌酶、热休克蛋白70、硫氧还蛋白、黏蛋白-1、黏蛋白-2、黏蛋白-5AC、黏蛋白-5B 和黏蛋白-19)[27]。也有研究报道抗性淀粉可以直接与免疫系统相互作用,激活免疫反应[28]。此外,有研究发现饲喂抗性淀粉可以提高C57BL/6 雄性小鼠的还原型谷胱甘肽循环水平[29]。谷胱甘肽是一种主要的抗氧化剂,谷胱甘肽的减少与心血管疾病、关节炎、糖尿病和几种恶性肿瘤有关,患有各种恶性疾病的患者的血浆谷胱甘肽水平显著降低[29]。此外,基于年轻人群和动物实验的研究报道,膳食抗性淀粉可有效改善胃肠道健康、葡萄糖耐受量、骨密度以及胰岛素敏感性等与衰老相关的指标[30]。
也有部分研究报道了摄入高剂量的抗性淀粉对机体的负面影响,如抗性淀粉发酵过程产生的大量气体会导致某些人的胃肠道不适和胃肠胀气[31]。也有报道称随着抗性淀粉剂量的增加,与炎症条件相关的有毒气体H2S 的浓度也会随之升高[32]。
2 抗性淀粉的多尺度结构特征
淀粉的多尺度结构模型阐释了淀粉颗粒内部由分子内/间氢键作用集合而成的从微米到纳米尺度的多种结构特征[33],具体包括其颗粒结构(2~100 μm),生长环结构(120~500 nm),止水塞(Blocklets)粒子结构(20~50 nm),层状结构(9~10 nm),结晶结构(nm)和分子结构(支链淀粉和直链淀粉链,0.1~1 nm)[34]。目前国内外学者针对抗性淀粉的多尺度结构进行了研究,包括颗粒结构[28]、长程/短程有序度、超分子结构(包含簇、螺旋、微晶[35])、分子质量、聚集态结构[36]、分子链结构[36]等。抗性淀粉颗粒与原淀粉相比,表面较为粗糙[8,37-38],形状和大小也发生了显著性变化。荞麦抗性淀粉颗粒比原淀粉颗粒更大,且无固定颗粒形状[39],鹰嘴豆抗性淀粉颗粒呈多角形或立方形[40]。不同类型的抗性淀粉在形貌结构上也存在一定的差异,如RS2 多数为不规则球形,少数为圆柱形,而RS3多呈类似于海绵结构的较大的膨胀颗粒[8,37-38]。不同来源的RS3 颗粒形状和大小也有一定的差异[8]。如大米RS3 样品呈碎片状和不规则结构,表面褶皱,部分样品表面出现孔洞状[41]。欧阳梦云[42]通过超声波间歇辅助双酶法和晶种诱导-双酶法制备的灿米RS3,表面粗糙,凹凸不平,出现“蜂窝”状结构。而利用压热-酶解法制备的芸豆抗性淀粉结构致密紧实,呈不规则而棱角分明的无定形团块状结构,横断面呈片层状结构[43]。油莎豆辛烯基琥珀酸酐淀粉脂(RS4)颗粒与原淀粉颗粒相比无明显变化,大颗粒呈卵型,小颗粒呈球形,表面光滑清晰,然而也有部分颗粒表面出现了轻微腐蚀和细纹[44]。辛烯基琥珀酰化改性温和的反应条件没有显著改变木薯淀粉和马铃薯淀粉的粒度,木薯RS4 形态与原淀粉相比也没用发生明显变化,而马铃薯RS4 颗粒表面形成了一些裂纹和气孔[45]。参薯(Dioscorea alata L.)原淀粉呈椭圆形,表面光滑紧密,而参薯RS3 颗粒呈现出不规则的黏连形(作者推断这与支链淀粉和直链淀粉的重结晶有关);参薯RS4 颗粒因有较多淀粉颗粒发生分解和直链淀粉浸出,其表面粗糙且有碎片黏附;参薯RS5 呈块状结构,表面也出现碎片状结构[46]。生姜抗性淀粉呈不规则致密的非晶聚集块形貌,表面粗糙无规则,呈层状、条状,有裂纹,具有B 型晶型[47]。据Zou 等[48]的报道,处于膨大期(山药的4 个生长阶段中的第3 个阶段,膨大期在块茎快速生长的秋季)的天然山药淀粉(ES-NS)和休眠期(山药生长的第4 个阶段,在冬季)的天然山药淀粉(DS-NS)与相对应时期的山药抗性淀粉颗粒在形状上没有明显差异(即颗粒呈多边形或椭圆形),而膨大期的山药抗性淀粉表面与休眠期的山药抗性淀粉相比,颗粒表面更粗糙。
抗性淀粉有A、B、C 以及V 型(复合晶型)4种结晶类型[16,49]。A 型结构主要存在谷物中,B 型结晶大多存在于块茎和富含直链淀粉的淀粉中,C型结晶是A 型和B 型的混合物,主要存在于豆类[48]。据文献报道,晶型呈A 型的有山药抗性淀粉[48]、经过发酵的马铃薯抗性淀粉、绿豆抗性淀粉、板栗抗性淀粉、锥栗抗性淀粉[50],以及RS4 型交联玉米抗性淀粉[51];呈B 型晶体的抗性淀粉包括绿豆抗性淀粉[50]、豌豆抗性淀粉[8]、马铃薯抗性淀粉[50]、玉米抗性淀粉[8],及美人蕉抗性淀粉[37]。据报道,II 型抗性淀粉包含了B 型或C 型的半结晶淀粉[52];III 型玉米抗性淀粉常呈现出B 型结晶结构[51];鹰嘴豆抗性淀粉颗粒在2θ 为17°、20°和22°处呈现出强衍射峰,为典型的C 型结晶结构[50],压热-酶解法制备的芸豆抗性淀粉晶形结构从原淀粉的A 型转变为抗酶解性更强的C 型[43];不同直链含量的大米III 型抗性淀粉呈现出B 型或C 型结晶[41]。灿米III 型抗性淀粉颗粒的晶型为B 型或B+V 型[38]。锥栗、板栗抗性淀粉晶型呈V 型结晶结构[50]。而荞麦抗性淀粉的峰形平缓,半峰宽>3°,为非晶峰,主要为玻璃体结构[39]。
3 抗性淀粉与肠道微生物
3.1 肠道微生物
人体肠道中寄居着种类繁多的微生物,称为肠道菌群。肠道细菌组成的庞大而复杂的微生态系统,在维持人体健康方面起重要作用[53]。肠道微生物分属1 000 多个物种[54],成人肠道中的微生物数量可达1012~1014个[55],总质量超过1.5 kg[56]。肠道中有超过300 多万个细菌基因[57],是整个人类基因组的至少150 倍[58]。尽管细菌数量(3.8×1013个)和人体细胞数(3.0×1013个)的比值随着研究的进展从100∶1 到10∶1 更新到接近1∶1,而这丝毫没有消减微生物区系的生物学重要性[59]。成人胃肠道的微生物可分为细菌(Bacteria)、古菌(Archaea)和真核生物(Euarya)[55]。居住在大肠中的绝大多数微生物是细菌[60],目前已鉴定出的人体肠道细菌有厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria)、梭杆菌门(Fusobacteria)、蓝藻菌门(Cyanobacteria)、疣微菌门(Verrucomicrobia)、螺旋藻门(Spirochaeates)、放线菌门(Actinobacteria)、VadinBE97 门、史氏甲烷短杆菌(Methanobrevibacter smithii)等10 大类[17],其中优势菌群主要有:1)厚壁菌门,该菌门占菌群总数的60%~65%[58],包含10 个核心属:布劳特氏菌属(Blautia)、丁酸弧菌属(Butyrivibrio)、粪球菌属(Coprococcus)、梭菌属(Clostridium)、优(真)杆菌属(Eubacterium)、粪杆菌属(Faecalibacterium)、颤杆菌克属(Oscillibacter)、玫瑰杆菌属(Roseburia)、瘤胃球菌属(Ruminococcus)和链球菌属(Streptococcus)[61]。2)拟杆菌门占菌群总数的20%~25%,主要属有另枝菌属(Alistipes)、拟杆菌属(Bacteroides)、副拟杆菌属(Parabacteroides)、叶琳单胞菌属(Porphyromonas)、普雷沃氏菌属(Prevotella)[58]和臭气杆菌属(Odoribacter)[61]。3)变形菌门占菌群总数的5%~10%[58],该菌门的主要代表有脱硫弧菌属(Desulfovibrio)、埃希氏菌属(Escherichia)、克雷伯氏菌属(Klebsiella)和志贺氏菌属(Shigella)[61]。4)放线菌门占菌群总数的3%,如双歧杆菌属(Bifidobacterium)[61]。
肠道微生物在人体的消化、营养吸收、能量供应、脂肪代谢、免疫调节等方面发挥着重要作用[53]。同时肠道微生物也可以抵御肠道病原体,并调节宿主体内的代谢途径。肠道微生物失调与多种代谢疾病密切相关,包括肥胖、代谢综合征糖尿病、结肠癌和炎症性肠道疾病等胃肠疾病[54]。此外,还有报道称肠道微生物群紊乱与神经系统性疾病有关[62]。因其规模巨大及维护健康的重要作用,肠道微生物被视为人体内的“新器官”[63]。最近的研究发现肠道菌群的组成受遗传、饮食习惯和生活方式等诸多因素的影响[41],而大肠中的绝大多数细菌以糖代谢为主,因此碳水化合物的供应被认为是控制微生物区系组成和代谢活动最重要的营养因素,而以抗性淀粉为代表的膳食纤维可被用于快速改变肠道微生物组成和活性[41]。
3.2 抗性淀粉与肠道微生物的相互作用研究
表1 列举了国内外研究学者在动物(大鼠[14,64]、小鼠[17,25,29,62,65]、仓鼠[24]、猪[66-67]、肉鸡[68])和人群[20,69](婴儿[70]、代谢综合症患者[71]、II 型糖尿病患者、慢性肾病患者[72])受试者中所进行的关于抗性淀粉体内和体外[11,13,70]的实验概述。在各种动物实验中,摄入抗性淀粉均引起了肠道微生物群组成的改变[73]。总体表现为有益菌群增加和有害菌群减少。研究发现抗性淀粉对肠道菌群的影响主要取决于抗性淀粉来源和类型,以及人类肠道微生物群的个体差异[11]。Li 等[11]在高直链玉米淀粉RS2和RS3 的体外发酵试验中发现,优杆菌科(Eubacteriaceae)和毛螺菌科(Lachnospiraceae)具有类似的促进碳水化合物代谢和转化功能。不同类型的RS 在体内可选择性地促进某些结肠菌群增殖。Martínez 等[74]对食用含RS2 和RS4 饼干受试者的粪便微生物进行16S rRNA 多重测序的结果显示,2 种RS 均引起了粪便微生物种群组成的显著变化,其中,RS4 增加了放线菌门和拟杆菌门的数量,同时减少了厚壁菌门的数量。在种水平上,RS4增加了青春双歧杆菌(Bifidobacterium adolescentis)和狄氏副拟杆菌(Parabacteroides distasonis)的丰度,与RS4 相比,RS2 则显著提高了布氏瘤胃球菌(Ruminococcus bromii)和直肠真杆菌(Eubacterium rectale,E.rectale)的比例。Kleessen 等[64]发现同一菌群对不同类型抗性淀粉的响应程度不同,具体表现为:饲喂改性马铃薯抗性淀粉(RS2)的大鼠盲肠中乳酸杆菌(Lactobacilli)、链球菌(Streptococci)和肠杆菌(Enterobacteria)的数量显著高于饲喂天然颗粒马铃薯抗性淀粉RS1 的大鼠,作者推测这可能与饲料中淀粉种类有关,饲喂含RS2 的饲料对肠道菌群的刺激作用更强,表明淀粉化学结构的微小修饰有可能改变肠道微生物区系的组成。
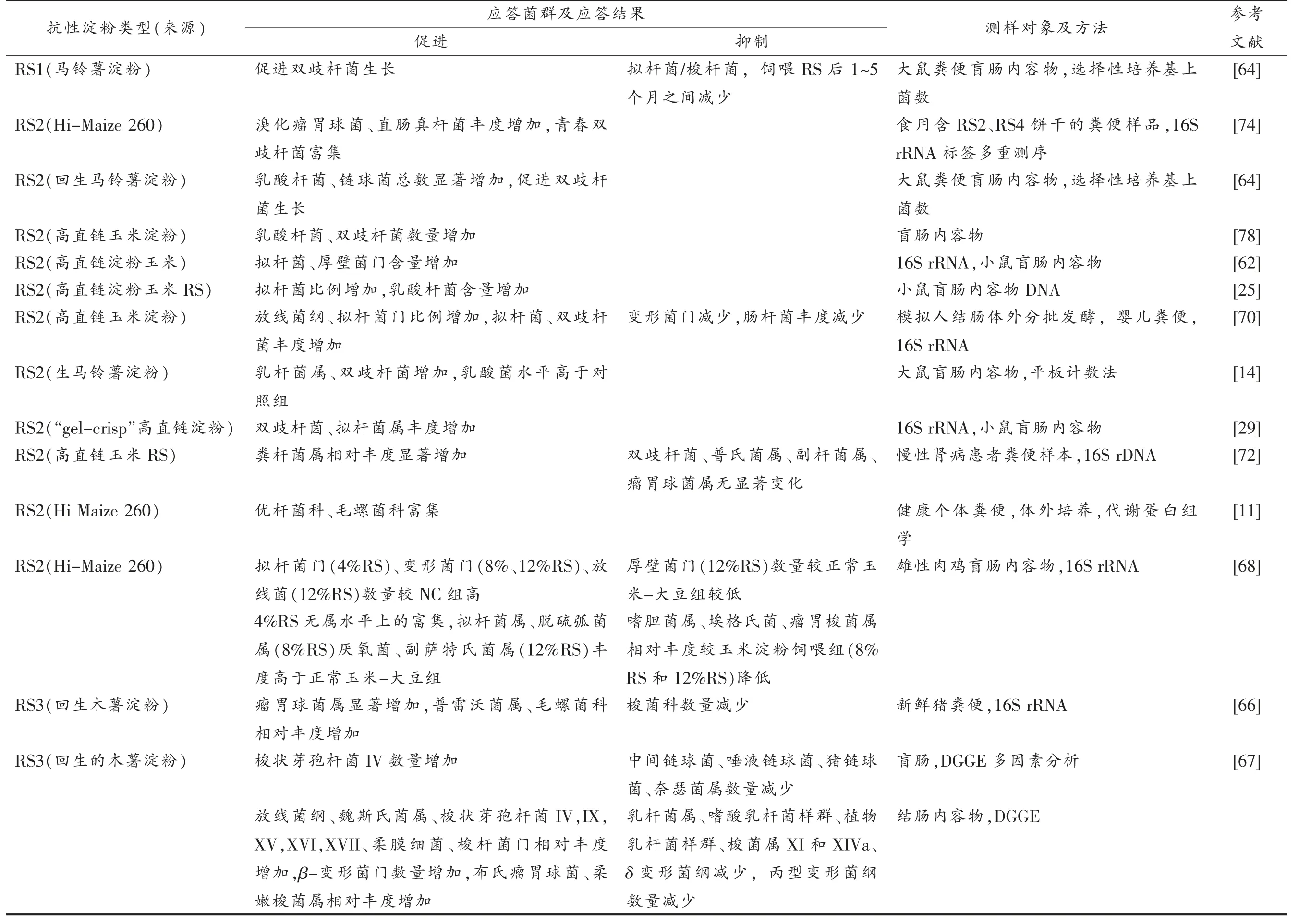
表1 肠道菌群对不同类型抗性淀粉的响应情况Table 1 Response of the intestinal flora to different types of resistantstarch
哺乳动物肠道中定居的细菌拥有比宿主更多的降解酶和更强的代谢能力。Herrmann 等[73]利用稳定同位素探针鉴定参与抗性淀粉同化的细菌菌种,发现拟杆菌门、普雷沃氏菌科(Prevotellaceae)和瘤胃菌科(Ruminococcacea)是抗性淀粉的主要同化菌。抗性淀粉首先在初级降解菌青春双歧杆菌和布氏瘤胃球菌的作用下降解为一些可溶性底物;接着梭状芽孢杆菌(Clostridia)如直肠真杆菌或拟杆菌(Bacteroidetes)如多形拟 杆菌(Bacteroides thetaiotaomicron)将这些可溶性淀粉和链长较长的低聚糖进一步降解为小分子寡糖[75]。最后,大肠杆菌(Escherichia coli)和罗伊氏乳杆菌(Lactobacillus reuteri)进一步利用释放出来的葡萄糖等分子质量较小的寡糖。Li 等[11]将粪便样本培养在添加了抗性淀粉的培养基上培养后发现,优杆菌科、毛螺菌科和瘤胃菌科的碳水化合物代谢和转运功能显著性增强。双歧杆菌科(Bifidobacteriaceae)淀粉代谢能力提高,同时,还观察到戊糖磷酸途径相关的类群也得到特异性富集。然而目前关于各菌群对抗性淀粉的作用机制尚不明确。
淀粉是一种分子质量较大的聚合物,在酶的作用下淀粉从细胞表面开始分解,并在酶的持续作用下逐渐裂解成葡萄糖、麦芽糖或者麦芽寡糖。据文献报道,在糖苷水解酶家族(Glycoside hydrolase,GH)中,GH3、GH13、GH14、GH15、GH57、GH119及GH126 与淀粉的降解有关[76]。其中,GH13 酶是肠道细菌中含量最丰富的酶之一,与淀粉的初始降解菌有关[77]。肠道细菌降解淀粉的机制主要取决于细菌细胞的生理特性和细菌所降解的淀粉类型。根据目前的研究报道,同时结合相关肠道菌群中淀粉活性酶的结构和功能,下文将对几种重要的具有抗性淀粉降解能力的肠道菌群进行综述。
4 布氏瘤胃球菌对抗性淀粉的降解作用
布氏瘤胃球菌在肠道菌群中占有较大的比例,布氏瘤胃球菌对抗性淀粉的响应明显[81]。有研究表明,布氏瘤胃球菌可以利用抗性淀粉[82],是抗性淀粉酵解过程中的关键初始微生物[83]。布氏瘤胃球菌对抗性淀粉具有高度专一性,可形成一种专用于降解抗性淀粉的保守胞外淀粉样复合体[84],这种淀粉样复合体是一种多酶复合物,可通过将多肽锚定到支架蛋白上的Dockerin-Coherin 蛋白结合域上,从而将碳水化合物的结合功能和酶功能绑定在一起[85]。布氏瘤胃球菌可通过这种细胞壁的锚定作用和“淀粉样复合体”的酶解作用联合对抗性淀粉进行利用与降解[85]。布氏瘤胃球菌在培养基中可将RS2 和RS3 分解为葡萄糖和麦芽糖[10]。布氏瘤胃球菌L2-63 基因编码的21 个糖苷水解酶(GHs)中有15 个属于淀粉水解酶GH13[86]。其中,GH13 基因在人类分离菌株之间具有显著保守性(在氨基酸水平上有90%~100%的同源性)[84],在高RS2 浓度下淀粉降解酶家族GH13、GH77 相关的糖苷水解酶以及GH13 的特定多糖结合模块均得到不同程度的升高[32]。据Mukhopadhya 等[84]的研究报道,4 株来自人体粪便的布氏瘤胃球菌菌株(L2-63,L2-36,5AMG,及ATCC 27255)都表现出较强的RS 降解能力。
Walker 等[87]对14 名超重男性的饮食和24 h粪便收集物进行分析,发现在接受高剂量RS3 饮食时,有2 名志愿者有超过60%的RS 未被发酵(分别为69%和65%),而其它12 名志愿者只有少于4%的RS 未被发酵,通过qPCR 检测,这2 位志愿者粪便菌群中的布氏瘤胃球菌数量较低。Ze 等[10]发现给上述1 名低RS 发酵志愿者的混合粪便细菌中添加布氏瘤胃球菌后,则可进一步提高RS3的体外发酵程度。Ze 等[10]的研究还发现,将布氏瘤胃球菌分别与多形拟杆菌、青春双歧杆菌、直肠真杆菌共同培养后,RS2 与RS3 的利用率显著提高(与每种菌株单一培养相比),证明布氏瘤胃球菌在刺激其它3 种菌对RS2 和RS3 利用方面的重要作用,也进一步表明布氏瘤胃球菌强大有效的酶系统,具有可使底物分解为可供其它菌群利用的功能[10]。Mukhopadhya 等[88]发现,将布氏瘤胃球菌L2-63、布氏瘤胃球菌L2-36、布氏瘤胃球菌ATCC 27255、布氏瘤胃球菌5AMG 和布氏瘤胃球菌YE282 在含RS 的M2 培养基中培养48 h 后,RS2 的总糖消失率约为77%~89%,而RS3 的总糖消失率为71%~87%。
5 双歧杆菌对抗性淀粉的降解作用
双歧杆菌(Bifidobacterium)是人类肠道中最初的定居者之一[89],是放线菌门革兰氏阳性菌的代表[90],双歧杆菌中,青春型双歧杆菌是成人大肠中最大的居民群之一[91]。双歧杆菌作为抗性淀粉的主要降解者[10,75,92],主要参与淀粉和蔗糖代谢途径及磷酸戊糖代谢途径[11]。
根据动物模型和人类受试者试验报道,肠道双歧杆菌的数量与宿主所摄入的抗性淀粉密切相关[93]。摄入抗性淀粉可显著促进双歧杆菌的增殖,饲喂玉米抗性淀粉4 周后,大鼠粪便中的双歧杆菌活菌数【(10.42±0.60)lg CFU/g 粪便】较对照组【(9.21±0.65)lg CFU/g 粪便】增加了1.2 lg CFU/g[94]。Zhang 等[79]研究发现,经纯化的莲子抗性淀粉可促进青春双歧杆菌的生长,提高丁酸的产量。据Bouhnik 等[95]的报道,每天摄入10 g 抗性淀粉的志愿者粪便中双歧杆菌数量从开始接受RS 时的(7.41±0.37)lg CFU/g 增加到(9.43±0.49)lg CFU/g(接受治疗7 d)。摄入马铃薯抗性淀粉使受试者粪便中双歧杆菌/青春双歧杆菌/斯特氏双歧杆菌的相对丰度增加了6.5 倍(P<0.001)[96]。一项分别以2%马铃薯抗性淀粉和绿豆抗性淀粉为碳源的体外发酵试验的发酵液中,双歧杆菌数量达8.07×107个/mL(马铃薯抗性淀粉)和2.1×108个/mL(绿豆抗性淀粉),以1%的锥栗抗性淀粉和板栗抗性淀粉为碳源的发酵液中,双歧杆菌数量分别为1.39×108个/mL 和1.87×108个/mL[97]。有研究发现不同含量的荞麦抗性淀粉对粪便中双歧杆菌的增殖有一定的促进作用,相对而言,较高剂量组的促进效果更加显著,最高数量可达到7.6×108个/mL[98]。据孙永康[99]的报道,鹰嘴豆抗性淀粉样品能促进双歧杆菌增殖,并且比低聚果糖能更早的促进双歧杆菌增殖,而发酵后期对双歧杆菌的增殖效果不如低聚果糖。骞宇[100]的研究发现,不同种类(RS2、RS3、RS4)及不同含量(5%,10%,15%)的抗性淀粉试验组均能促进双歧杆菌和乳酸杆菌的生长增殖。其中RS3 各含量组与RS2 和RS4 组相比,对双歧杆菌生长增殖的促进作用均更显著;同一类型RS 组中,10%含量的RS 对双歧杆菌和乳酸杆菌的生长增殖的促进效果优于5%和15%的含量组。据Turroni 等[101]报道,在单独培养时,两歧双歧杆菌 PRL2010(Bifidobacterium bifidum PRL2010)在RS2 基质上未表现出显著的生长增殖,然而将两歧双歧杆菌PRL2010 与短双歧杆菌12L(Bifidobacterium breve 12L),青春双歧杆菌22L 或嗜热双歧杆菌JCM1207(Bifidobacterium thermophilum JCM1207)共培养时,可以显著提高两歧双歧杆菌PRL2010 的数量。同时,两歧双歧杆菌PRL2010 与青春双歧杆菌22L 或短双歧杆菌12L 在含RS 的MRS 上共同培养时,细胞数量分别增加了约3 或4 倍,表明双歧杆菌菌株具有交叉摄食能力。当青春双歧杆菌22L 和JCM1207共同培养时,葡萄糖和麦芽糖的消耗增大1 倍,且共培养时醋酸盐和乳酸的产量也较单一菌落培养时增加了2 倍。
青春双歧杆菌是RS2 的主要降解菌,可以在以RS2 为唯一碳源的培养基上生长[6]。表面粗糙且晶体结构完善的RS 为青春双歧杆菌提供了更好黏附条件和生长底物,有助于促进双歧杆菌的耐受性和增殖能力[102]。Jung 等[103]的研究发现,将青春双歧杆菌P2P3 与抗性淀粉共孵育48 h 后,抗性淀粉颗粒分解消失,细菌接种后,分散的RS 颗粒开始形成凝结团。降解过程中,在RS 颗粒群周围观察到大量的细胞,青春双歧杆菌P2P3 对RS的利用率达63.3%。青春双歧杆菌P2P3 与未糊化的RS2 共培养后,可检测到葡萄糖、麦芽糖和麦芽三糖等还原糖的存在[104]。同时,在该菌株的基因组中发现了19 个编码α-葡聚糖底物的活性淀粉水解酶基因,作者认为这些淀粉酶可能直接或间接参与抗性淀粉颗粒降解以及从抗性淀粉颗粒降解为小分子糖的过程。该学者推断其中有3 种酶包括α-淀粉酶、糖原去分支酶和普鲁兰酶含有碳水化合物结合模块,均参与到了酶和底物的结合的过程中[103]。在青春双歧杆菌的基因组中鉴定出3种酶(RSD1、2、3),比较基因组学结合酶学试验证实RSDs 可能是双歧杆菌在特定的生态位(人类肠道)中降解RS 的关键酶[93]。
降解抗性淀粉的双歧杆菌有多个预测的细胞表面GH13。多模块的GH13 酶包含多个CBM(Carbohydrate-binding modules,碳水化合物结合模块),其中至少有1 个酶包含CBM74[75]。CBM74可以与颗粒状淀粉特异性结合,增强α-淀粉酶降解抗性淀粉的能力,同时,CBM74 也可能通过与淀粉颗粒结合来辅助GH13α-淀粉酶降解抗性淀粉[105]。大多数CBM74 家族来源于人类肠道双歧杆菌的α-淀粉酶[105]。且在哺乳动物肠道双歧杆菌α-淀粉酶中的相对丰度较高,是哺乳动物胃肠道中降解抗性淀粉的关键所在[105]。
6 其它肠道菌群对抗性淀粉的应答
除了上述2 种抗性淀粉初始降解菌外,抗性淀粉完全降解和发挥生理功能还需要其它菌群的协同作用。抗性淀粉在初级降解菌布氏瘤胃球菌和青春双歧杆菌的作用下开始降解,初级降解菌改变了淀粉的颗粒结构,并释放出一些可被其它菌群利用的可溶性底物,如葡萄糖、麦芽低聚糖,以及一些可溶性淀粉。葡萄糖、麦芽寡糖可以支持罗伊氏乳杆菌、大肠杆菌和多形拟杆菌的生长。而初级降解菌降解抗性淀粉产生的可溶性淀粉被直肠真杆菌或者多形拟杆菌获取利用[75]。Jung 等[104]的研究证实,多形拟杆菌(ATCC 29148)不能降解RS2 却具有降解可溶性淀粉的能力,可在含0.5%的可溶性淀粉的培养基中生长良好。将其和青春双歧杆菌P2P3 共同培养时,抗性淀粉降解率(24 h:9%;36 h:42%)显著低于较单独培养青春双歧杆菌时RS 的降解率(24 h:22%;36 h:55%),表明多形拟杆菌ATCC 29148 生长需要青春型双歧杆菌P2P3 释放的寡糖作为养料。而随着多形拟杆菌ATCC 29148 的生长,青春型双歧杆菌P2P3 的相对比例下降,导致抗性淀粉降解率随之下降。而直肠真杆菌可以附着在淀粉颗粒上,促使淀粉颗粒接近布氏瘤胃球菌,在布氏瘤胃球菌的作用下,抗性淀粉暴露出可被直肠真杆菌接触到的无定形区域,并释放出能被直肠真杆菌摄取的初级产物[106]。在次级降解菌的作用下,初级降解产物被发酵为短链脂肪酸并发挥其生理功能[75]。
结肠微生物和肠道之间重要相互作用关键在于结肠上皮细胞中短链脂肪酸代谢[107]。乙酸是肠道内含量最丰富的短链脂肪酸[108],由丙酮酸经乙酰辅酶A 或Wood-Ljungdahl 途径产生[109],拟杆菌属、双歧杆菌属、真杆菌属、瘤胃球菌属、消化链球菌属、梭菌属和链球菌属均可产生乙酸[110]。大部分乙酸被吸收后进入血液,在肌肉、脾脏、心脏和脑组织内代谢[111];丙酸是拟杆菌属发酵的主要产物,可以通过琥珀酸途径、丙烯酸途径和丙二醇途径产生[109],其作为糖异生的底物,能降低胆固醇活性,刺激瘦素释放[112];直肠真杆菌和肠道菌群的其它成员产生的丁酸是初级发酵的终产物[113],也是结肠细胞的主要能量来源[114],可以促进上皮细胞发育[108],并在多种炎症性肠道疾病的预防和治疗中发挥着重要作用。一些疾病(如溃疡性结肠炎、克罗恩病、糖尿病等)患者的肠道中,丁酸产生菌的丰度明显低于健康人群[113]。此外,丁酸有提高胰岛素敏感性,抑制结肠癌细胞的生长,诱导分化肠上皮细胞,刺激细胞骨架形成及改变基因表达等作用[108]。乳酸杆菌可以发酵抗性淀粉产生乙酸、丙酸和丁酸,这些短链脂肪酸已被证明具有降低糖尿病风险,降低血清胆固醇,改善通便,预防结肠癌和促进有益菌群生长等益生作用[115]。值得一提的是,一些微生物如厌氧菌属(Anaerostipes spp)、霍氏真杆菌(Eubacterium hallii)可以利用乳酸和乙酸合成丁酸,从而阻止乳酸蓄积并维持肠道内环境稳定[109]。
7 结语
肠道菌群被视为人体内的“新器官”,在维持机体健康方面发挥着重要作用。抗性淀粉作为肠道菌群在营养物质、初级降解菌和次级降解菌的协同作用下,被降解为短链脂肪酸等产物,短链脂肪酸是肠道微生物群和宿主健康之间的关键信号分子。抗性淀粉及其发酵产物对人体有降血糖,降低胆固醇,预防肥胖,促进矿物质吸收,防止腹泻,预防胆结石,改善代谢紊乱,减缓慢性疾病发展等诸多健康益处。肠道微生物对抗性淀粉的降解作用一直备受学者关注。然而目前研究多停留在揭示分类学和功能反应阶段,关于肠道菌群对抗性淀粉的降解机制,尤其是已知初级降解物及特定的次级降解物之间的相互作用,还有待深入研究。
