牛肉源热杀索丝菌的致腐差异和比较基因组学研究
2024-02-23方金玉吴诗媛施永清石双妮朱军莉
方金玉,张 俊,吴诗媛,施永清,石双妮,朱军莉
(浙江工商大学食品与生物工程学院 浙江省食品安全重点实验室 杭州 310018)
牛肉作为我国主要消费的肉类,营养价值高,是肉类消费市场的重要组成部分。国家统计局数据显示,2021 年全国牛肉产量698 万t,同比增长3.7%[1]。随着冷藏保鲜技术的发展,牛肉的消费市场进一步扩大。然而在冷链运输过程中,由于嗜冷微生物的新陈代谢活动,产生大量的醛、酮、有机酸和挥发性物质,导致牛肉品质下降[2]。在冷鲜牛肉基质中,主导腐败活动的微生物只有1 种或几种[3],包括假单胞菌和热杀索丝菌。
热杀索丝菌(Brochothrix thermosphacta)为环丝菌属,革兰氏阳性,需氧兼性厌氧型嗜冷菌,短链状或长丝状链状。热杀索丝菌在0~30 ℃环境下生存,最佳生长温度范围20~25 ℃,能够在pH 值5~9 的范围内生长[4]。该菌自1951 年首次从猪肉香肠中分离得到,广泛存在于猪肉、牛肉、羊肉、鸡肉等多种生鲜肉及肉制品中[5]。从牛肉胴体、动物皮肤和瘤胃到屠宰场设备及最终销售产品,热杀索丝菌在牛肉生产链中分布广泛[6]。因其普遍存在且具有耐低温的特性,故热杀索丝菌是低温贮藏肉类以及海产品中的优势腐败菌。刘爱芳等[7]研究表明,4 ℃冷藏金枪鱼中热杀索丝菌能够产生大量挥发性盐基氮。除有氧包装外,热杀索丝菌还是气调包装和真空包装产品中的优势微生物[8-9]。
随着高通量测序技术的发展,越来越多的菌株完成了全基因组测序。比较基因组学可以基于比较对象进行基因组元件的预测,以及功能蛋白注释,能够更加全面地了解不同菌株的发育特点,解释表型差异。目前,比较基因组学技术逐渐被应用于食品科学和安全领域研究。例如:彭明芳等[10]对3 株乳杆菌的比较基因组学研究发现,其具有丰富的碳水化合物酶谱。Li 等[11]从冷藏虾中分离得到波罗的海希瓦氏菌,发现其基因组中8.26%的编码基因被注释到氨基酸转运和代谢功能,表明波罗的海希瓦氏菌具有良好的降解蛋白质的能力。Stanborough 等[12]分析比较不同来源的13 株莓实假单胞菌和7 株隆德假单胞菌,发现其具有高度的遗传多样性,且不同菌株的挥发性成分产生能力存在差异。目前,从比较基因组学角度对热杀索丝菌的致腐能力的研究鲜有报道。本研究以冷鲜牛肉热杀索丝菌分离株为研究对象,以挥发性盐基氮(TVB-N)和乙偶姻为指标,筛选致腐差异的菌株,借助全基因组测序技术,分析预测致腐基因的分布,对深入探究该菌的致腐机制具有重要意义。
1 材料与方法
1.1 材料与试剂
从杭州市某菜市场购买的冷鲜牛肉冰运至实验室,无菌条件下分割成片状(长10 cm×宽8 cm×厚1.5 cm),放置于聚乙烯透氧托盘包装,在4 ℃贮藏。
热杀索丝菌分离培养基(Steptomycin thallous acetate agar,STAA),青岛海博生物技术有限公司;琼脂糖、缓冲液、引物、DNA Marker,上海生工有限公司;氧化镁、硼酸、甲醇、乙腈,国药集团化学试剂有限公司;盐酸、乙偶姻、萘酚、肌酸等,阿拉丁公司;Dneasy Blood &Tissue Kit 试剂盒,美国Kapa Biosciences 公司。
1.2 仪器与设备
3-18K 型台式高速冷冻离心机,德国Sigma公司;LRH-250A 型生化培养箱,上海一恒科技有限公司;BHC-1300 型生物安全柜,苏州市金净净化设备科技有限公司;FE20 型数显pH 计,梅特勒-托利多仪器(上海)有限公司;UV-1800 型分光光度计,日本Kyoto 公司;KDN-103F 型凯氏定氮仪,上海华烨公司;Agilent 2100 型生物分析仪,美国Agilent 公司;NovaSeq 6000 型测序仪,美国Ilumina 公司;Vanquish 型电泳仪、Qubit@2.0 型荧光计,美国Thermo Fisher Scientific 公司。
1.3 方法
1.3.1 热杀索丝菌的分离纯化及鉴定 牛肉在4℃贮藏初期和贮藏末期5 d 取样25 g,于225 mL生理盐水中,稀释涂布于STAA 培养基培养,挑取典型形态的菌落经纯化培养后,革兰氏染色。参考Illikoud 等[13]方法对分离得到的革兰氏阳性菌,通过rpoB 特异性引物(Fw1:5’-GCGTGCATTAGG TTTCAGTACA -3’,Rev1:5’ -TCCAAGACCAGA CTCTAATTGCT-3’)扩增,目的片段大小为396 bp,以李斯特菌及金黄色葡萄球菌为阴性对照。琼脂糖凝胶电泳观察分离菌株扩增出特定目的片段,以此鉴定。
1.3.2 热杀索丝菌的腐败能力测定
1)无菌牛肉汁的接种 根据顾春涛等[14]的方法稍作修改,将牛肉经无菌绞肉机绞碎后取25 g 在100 mL 无菌水中均质1 min,在4 ℃离心(6 000×g,20 min)。然后将上清液过0.45 μm 和0.22 μm 无菌滤膜得到无菌牛肉汁。将分离株以1‰接种于牛肉汁中,4 ℃培养5 d 和9 d 取样分析。
2)挥发性盐基氮TVB-N 测定 参考GB 5009.228-2016 中半微量定氮法检测TVB-N 值。取10 mL 牛肉汁,加入饱和氧化镁溶液和2%硼酸溶液,以甲基红-溴甲酚绿指示剂,放置于半自动凯氏定氮仪中反应,用0.01 mol/L HCl 滴定。结果表示为mg N/100 g 牛肉。
3)产乙偶姻能力测定 根据Illikoud 等[15]的方法,取样品离心的上清液,分别按顺序添加0.05 g/L 肌酸、0.5 g/L 萘酚,95%乙醇,0.5 g/L KOH,在波长560 nm 处测吸光值。同时以不同浓度的乙偶姻标准品绘制标准曲线,由标准曲线计算样品乙偶姻浓度。
1.3.3 热杀索丝菌强弱腐败株全基因组测序
1)基因组DNA 的提取 将分离菌株BT25和BT27 划线至STAA 培养基,经30 ℃培养24 h,挑取单个菌落接种于5 mL TSB,过夜培养。采用Dneasy Blood&Tissue Kit 试剂盒提取2 株分离菌株基因组DNA。琼脂糖凝胶电泳检测提取的DNA,并通过荧光计(Thermo Scientific)进行定量。
3)基因测序及组装 全基因组测序由北京诺源生物信息技术有限公司完成。使用Clean Data、SOAP denovo 和SPAdes 软件进行基因组组装,将初步组装结果与CISA 软件进行整合,选出支架最少的组装结果;使用gapclose 软件填补空白,通过过滤低测序深度(小于平均深度的0.35)的reads 去除相同的lane 污染,得到最终组装结果,随后过滤掉500 bp 以下的片段。
4)基因预测与功能注释 采用geneMarkS对测序基因组进行编码基因预测,并通过tRNAscan-SE、Rfam、RNAmmer 分析全基因组特征。预测的蛋白序列通过Clusters of Orthologous Groups of proteins(COG)、Kyoto Encyclopedia of Genes and Genomes(KEGG)、Carbohydrate-Active enzymes(CAZy)数据库进行功能注释。
1.3.4 数据处理与统计分析 热杀索丝菌在牛肉汁中腐败试验重复3 次,每组样品设立3 组平行,结果取平均值。采用GraphPad Prism 8.0 版本进行数据处理及绘图。显著性差异通过GraphPad Prism 8.0 的two-way ANOVA 分析,P<0.05 表示有统计学显著性差异。
2 结果与分析
2.1 热杀索丝菌的分离和鉴定
通过STAA 分离培养得到29 株分离株,经革兰氏染色后形态为蓝色,细杆状。采用rpoB 特异性基因鉴定热杀索丝菌分离菌株。如图1 所示,有29 株分离株成功扩增出396 pb 的目的条带,鉴定为热杀索丝菌,以单增李斯特菌和金黄色葡萄球菌作为阴性对照,发现未扩增出条带。Adékambi等[16]发现编码RNA 聚合酶β 亚基的rpoB 管家基因可以针对一系列密切相关的细菌物种,如乳酸菌和李斯特菌的DNA,进行细菌鉴定和系统发育研究。

图1 牛肉中热杀索丝菌的分离和鉴定Fig.1 Isolation and identification of B.thermosphacta strains isolated from beef
2.2 热杀索丝菌分离株的致腐能力评价
热杀索丝菌29 株分离株接种于灭菌牛肉汁中,以TVB-N 和乙偶姻为指标评价致腐能力。如图2 所示,所有分离株的TVB-N 生成缓慢,经9 d 冷藏下含量为7.0~12.6 mg N/100 g,均未超过国家标准,其中BT23 和BT30 最高,含量增长至11.5,12.6 mg N/100 g。TVB-N 是评价肉质新鲜度的重要指标,国家标准规定新鲜肉中TVB-N 含量应不超过15 mg N/100 g[7]。另外,29 株分离株在牛肉汁中的乙偶姻增加明显,在第5 天BT27、BT01、BT2、BT14、BT17、BT24 从高到低的乙偶姻产量为94.0~81.5 μg/mL,超过60 μg/mL,而BT16、BT25较低。牛肉汁冷藏至第9 天,BT06、BT09、BT20、BT21、BT26、BT27的乙偶姻产量为101.7~139.3 μg/mL,并且BT27 的产量最高,达到139.3 μg/mL,而BT16、BT25 仍较低。乙偶姻为热杀索丝菌在牛肉中的重要腐败代谢产物,29 株分离株在牛肉汁中的腐败能力存在差异,BT25 产乙偶姻能力最低,而BT27 最高,因此选择BT25 和BT27 强弱菌株进行比较基因组学分析。
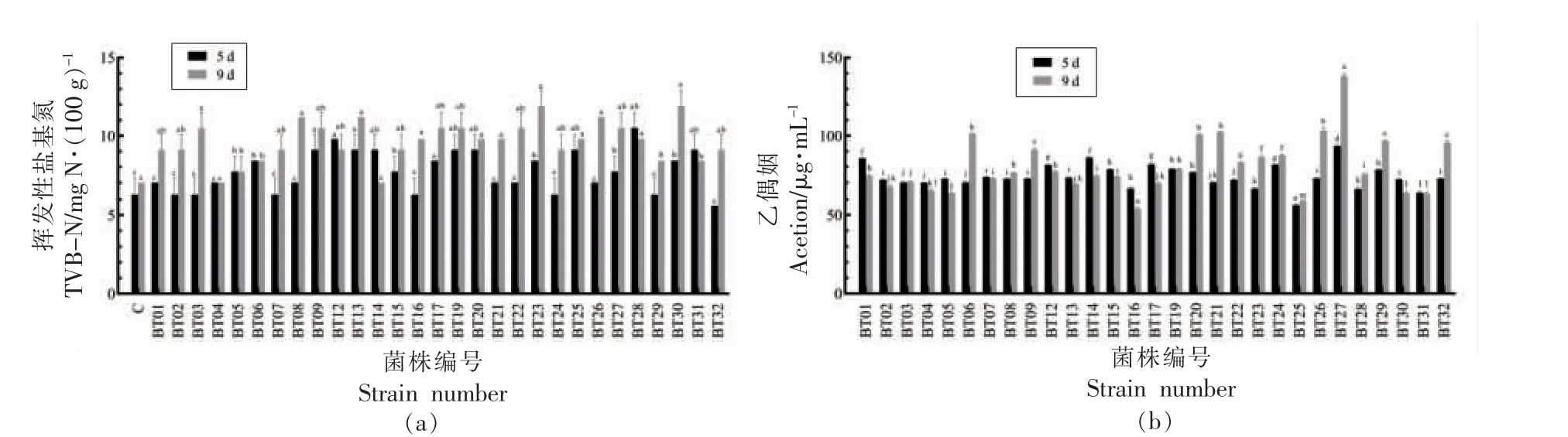
图2 热杀索丝菌分离株在4 ℃牛肉汁中TVB-N(a)和乙偶姻(b)的变化Fig.2 Changes of TVB-N(a)and acetion(b)of B.thermosphacta inoculated in sterile beef juice stored at 4 ℃
热杀索丝菌是导致冷鲜肉类腐败的优势嗜冷菌之一,使冷却肉产生具有异味的挥发性有机化合物,如乙偶姻、二乙酰,使腐败肉产生令人不适的奶酪或奶制品风味特征,多数情况下还会导致冷鲜肉表面发生绿变并伴随绿色液体产生[17]。本研究表明,热杀索丝菌对蛋白质和非蛋白质含氮化合物的降解能力较弱,所有分离株的TVB-N 积累量低,与假单胞菌、希瓦氏菌和气单胞菌等腐败菌致腐表型不同[18]。课题组前期研究已发现[19],热杀索丝菌能快速降解猪肉和牛肉中的葡萄糖,导致pH 值下降,积累大量的乙偶姻。分离株在牛肉汁中产乙偶姻能力较强,且不同菌株间具有差异,其中菌株BT27 最高,BT25 最弱,这与其分解代谢葡萄糖产生乙偶姻有关[20]。
2.3 热杀索丝菌强弱腐败株比较基因组学分析
2.3.1 两株菌基因组特点 利用Illumina NovaSeq PE150 测序平台测定两株强、弱致腐性的热杀索丝菌的全基因序列。经组装拼接后,其基因组特征如表1 所示。BT27 基因全长为3 028 989 bp,86 个基因组重叠群,GC 含量为36.10%,其中包含了2 980 个编码基因,预测出rRNA 和tRNA数量分别为6 个和51 个;BT25 基因全长为2617130 bp,9 个基因组重叠群,GC 含量为36.18%,其中包含了2 526 个编码基因,预测出rRNA 和tRNA 数量分别为4 个和20 个。

表1 两株热杀索丝菌基因组特点Table 1 Genome features of two B.thermosphacta isolates
2.3.2 泛基因组分析 泛基因组(Pan genome)是某一物种全部基因的总称,包括核心基因(Core genome)、附属基因(Accessory gene)和特异性基因(Unique gene)[21]。核心基因组是所有菌株中都存在的一组基因,主要参与基础生物学过程[22];而附属基因则是指存在于生物体子集或单个生物体中编码次级代谢途径相关的基因;特异性基因指仅在某一菌株中独特存在的基因。如图3 显示,热杀索丝菌BT27 和BT25 菌株一共预测出3 887 个基因,其中核心基因有1 491 个,占预测总基因数的38.4%,而独特基因分别为1 419 个和977 个,占预测总基因数的36.5%和25.1%。

图3 两株热杀索丝菌泛基因组分析Fig.3 Pan genome of two B.thermosphacta isolates
2.3.3 基因功能注释
2.3.3.1 COG 注释 COG(Cluster of orthologous groups of protein)蛋白数据库根据细菌、藻类和真核生物完整基因组的编码蛋白系统进化关系分类构建而成。两株热杀索丝菌的COG 注释如图4 所示。BT27 和BT25 的基因组上分别有2 319 个和1 966 个编码基因被注释到22 个COG 功能分类上,两株热杀索丝菌的COG 功能注释结果相似,其中蛋白编码基因主要集中分布在氨基酸转运代谢(255,203 个基因)、碳水化合物转运代谢(245,175 个基因)、翻译、核糖体结构与生物合成(185,197 个基因)、转录(209,166 个基因)等功能分类上;而分布在细胞运动、胞外结构的编码基因数量较少。且在大部分COG 功能分类中,BT27 的基因数量均多于BT25。

图4 两株热杀索丝菌COG 功能注释Fig.4 COG functions prediction of two B.thermosphacta isolates
2.3.3.2 KEGG 注释 KEGG(Kyoto encyclopedia of genes and genome)是一个整合了基因组、化学分子和生化系统等方面的数据库,能够系统分析基因产物和化合物在细胞中的代谢途径以及这些基因产物的功能[23]。图5 为热杀索丝菌代谢相关通路注释结果,BT27 和BT25 分离菌株分别有1 460 个和1 175 个编码基因被注释到代谢功能中的12 个功能分类。除全局和概览通路外,2 株热杀索丝菌基因涉及最多的通路为碳水化合物代谢,其中BT27 有240 个基因,BT25 有175 个基因,其次是氨基酸代谢,两者均超过140 个基因。
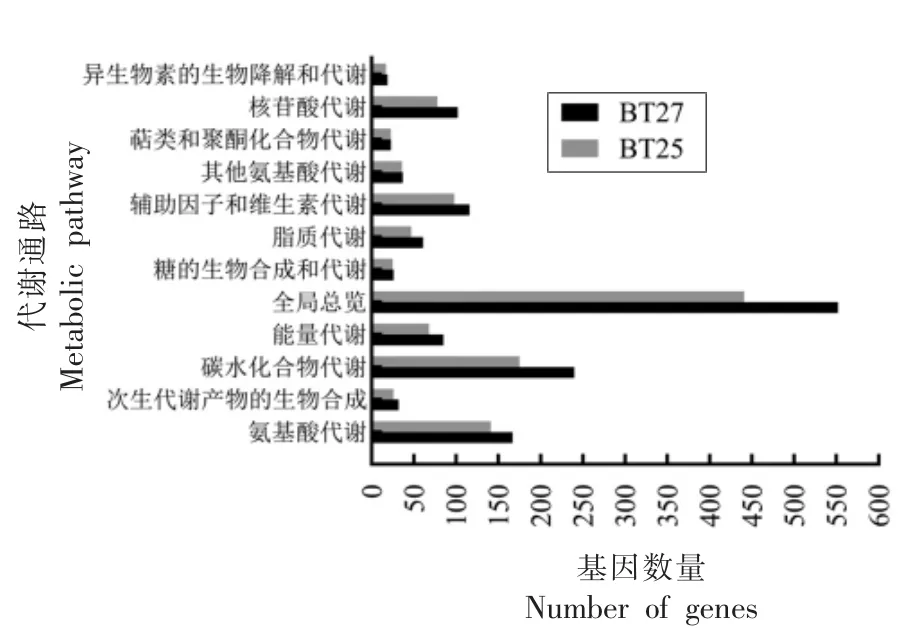
图5 两株热杀索丝菌KEGG 代谢功能注释Fig.5 KEGG functions prediction of two B.thermosphacta isolates
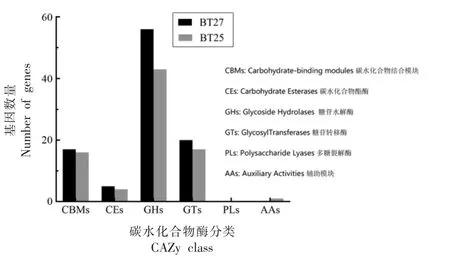
图6 两株热杀索丝菌碳水化合物酶的功能预测Fig.6 CAZymes functions prediction of two B.thermosphacta isolates
2.3.3.3 碳水化合物酶 碳水化合物活性酶(Carbohydrate-activeenzymes,CAZymes)为能催化碳水化合物降解、修饰、以及生物合成的相关酶系家族[24]。其包含6 个主要分类,在BT25 中发现1 个编码AAs 的基因。
两株腐败强、弱菌株的全基因组测序发现,BT27 和BT52 的核心基因组比例达到38.4%,表明两株菌同源性较高。CAZy 注释显示热杀索丝菌的基因组中存在大量参与碳水化合物代谢的编码基因和编码GHs 基因,GHs 家族基因编码的糖苷水解酶具备催化塔格糖苷、阿拉伯呋喃糖苷、甘露糖苷等糖类的功能[25]。BT27 的基因组中还有相关的基因为lacC(BT27_02234),araQ(BT27_00424),manA(BT27_00736)。Alexandrakis 等[26]研究显示葡萄糖、核糖、甘油、甘油-3 磷酸和肌苷是热杀索丝菌在肉中生长的底物,其中葡萄糖是首先被利用的碳源。另一方面,COG 注释显示两株菌也有编码氨基酸代谢的基因,然而数量较少,如sdhA基因(BT27_00143)编码的L-丝氨酸脱水酶,oppB基因(BT27_00600)编码寡肽转运系统渗透蛋白。然而,它们可利用的氨基酸范围非常窄。Illikoud等[27]发现热杀索丝菌基因组仅存在编码降解亮氨酸以产生异戊酸盐和3-甲基丁醇所需酶的基因。基于两株强、弱腐败株的比较基因组学发现,腐败能力较强的BT27 被注释到碳水化合物代谢和碳水化合物酶的编码基因数量均多于腐败能力较弱的BT25,提示热杀索丝菌的腐败潜力取决于碳水化合物的代谢能力。相似地,猪肉来源热杀索丝菌基因中也有大量磷酸烯醇式丙酮酸依赖型磷酸转移酶系统(PTS)的底物特异性基因,同时还存在编码核糖、麦芽糖和肌醇转运体的基因[28],提示热杀索丝菌的碳源利用能力极强。
PTS 可以将多种糖类及其衍生物进行磷酸化,然后运输到细胞内。绝大多数细菌PTS 的基本组成几乎都包含2 个磷酸转移酶:酶I(EI)和组氨酸磷酸载体蛋白(HPr),以及依赖于不同种类糖的特异性酶II 复合物(EIIA,EIIB,EIIC,EIID)[29]。编码EI 和HPr 蛋白的基因通常在同一个操纵子上,即ptsHI 操纵子。EI(约570 个残基)由ptsI 基因(BT27_00962)编码,HPr(约90 个残基)由ptsH(BT27_00963)基因编码[30]。BT27 和BT25 中涉及编码PTS 的基因多达17 种,bglF、celA、crr、dhaK、dhaL、dhaM、fruA、gmuC、malX、manX、manY、manZ、mtlA、nagP、ptsG、treP、ulaA,而致腐能力更强的BT27 中的基因数量更多。它们编码的蛋白质主要用于能量运输过程中碳水化合物(如葡萄糖、果糖、甘露糖、乳糖、海藻糖和纤维二糖等)的吸收,并将糖类物质催化转化为相应的磷酸酯。
此外,BT27 中还存在多种糖类的转运系统,如malG(BT27_01417)编码麦芽糖转运系统渗透蛋白,在麦芽糖进入细胞内后由 malP(BT27_00184)编码的麦芽糖羧化酶催化进入相应代谢途径。rbsA(BT27_02029)、rbsB(BT27_02027)和rbsC(BT27_02028)基因分别调节核糖与ATP的结合以及细胞膜渗透。阿拉伯糖的降解需要araB、araA 和araD 3 个基因,分别编码核酮糖激酶、阿拉伯糖异构酶和核酮糖-5-磷酸差向异构酶[31],在BT27 相应的基因为BT27_01643、BT27_00421和BT27_00424。
3 结论
热杀索丝菌是冷鲜肉中常见的腐败菌,近期研究发现猪肉、牛肉、羊肉及腌制肉中均有存在[32],表现出很强的代谢多样性和广泛的定殖环境,然而,对热杀索丝菌致腐机制的研究仍较少。本研究从冷鲜牛肉样品中共分离得到29 株热杀索丝菌,分离株的TVB-N 形成较弱,而产乙偶姻能力较强,且基于乙偶姻差异获得强、弱腐败菌BT27 和BT25。通过比较基因组学解析了冷鲜牛肉分离的热杀索丝菌降解碳水化合物分解酶的基因。2 株热杀索丝菌核心基因组中涉及调控基因比例较高,其致腐能力与碳水化合物基因编码的酶和蛋白质相关,从基因层面上阐明BT27 菌株的强致腐能力的分子基础。研究为热杀索丝菌分离株的基因组结构中差异性研究提供了线索,为后续热杀索丝菌代谢特征、致腐机理和调控通路研究奠定了良好的基础。
