基于加性-显性-上位性(ADAA)模型与主-多基因模型的陆地棉产量与品质性状的遗传分析
2024-01-30郭金成曹新川宋佳赵玉玲何良荣
郭金成 曹新川 宋佳 赵玉玲 何良荣





郭金成,曹新川,宋 佳,等. 基于加性-顯性-上位性(ADAA)模型与主-多基因模型的陆地棉产量与品质性状的遗传分析[J].江苏农业学报,2023,39(9):1793-1803.
doi:10.3969/j.issn.1000-4440.2023.09.001
收稿日期:2022-12-09
基金项目:国家自然科学基金项目(31550010);华中农业大学-塔里木大学科研联合基金项目(HNLH202007)
作者简介:郭金成(1999-),男,甘肃定西人,硕士研究生,研究方向为陆地棉遗传育种。(E-mail)526097667@qq.com
通讯作者:何良荣,(E-mail)hlrzky@163.com
摘要:本试验为进一步明确陆地棉(Gossypium hirsutum L.)杂交后代产量、纤维品质性状的遗传规律,以9个陆地棉品种为亲本设计不完全双列杂交(NCⅡ)。对2019-2021年的亲本、18个F1和F2组合及组合TH14-22×B7的127个F2单株分别采用加性-显性-上位性(ADAA)模型与主基因-多基因遗传模型,从基因整体与个体水平进行分析。NCⅡ分析结果表明:所有性状均存在基因型与环境的互作,主要通过显性×环境来体现,上位性×环境不可忽视。多数性状的遗传以加性效应为主,整齐度以显性效应为主,伸长率无显著的基因主效应。上半部平均长度与整齐度具有正向平均优势,其他性状无明显的平均优势。分离分析结果表明:各性状普遍受主基因控制。单株铃数、衣分与比强度的最适遗传模型为2MG-EA,单铃质量与上半部平均长度最适遗传模型为2MG-A,整齐度与马克隆值最适遗传模型为2MG-AD,伸长率最适遗传模型为1MG-AD。单铃质量、衣分、上半部平均长度、整齐度与比强度主基因遗传率较高,单株铃数、马克隆值与伸长率的遗传率较低。ADAA模型分析结果与主基因-多基因分析结果的相互补充,可有效了解各性状的遗传特征,为采用适当的育种改良策略提供参考。
关键词:陆地棉(Gossypium hirsutum L.);ADAA模型;主基因-多基因;遗传分析
中图分类号:S562 文献标识码:A 文章编号:1000-4440(2023)09-1793-11
Genetic analysis of yield and quality traits of upland cotton based on ADAA and major-polygene models
GUO Jin-cheng CAO Xin-chuan SONG Jia ZHAO Yu-ling HE Liang-rong
(College of Agronomy, Tarim University, Alar 843300, China)
Abstract:In order to further understand the genetic law of yield and fiber quality traits of hybrid progenies of upland cotton (Gossypium hirsutum L.), incomplete diallel cross (NCⅡ) was designed with nine upland cotton varieties as parents. Parents,18 F1 and F2 combinations and 127 F2 individuals from TH14-22 × B7 from 2019 to 2021 were analyzed by using additive-dominance-epistasis (ADAA) model and major gene-polygene genetic model from the whole gene level and individual level. The results of NCⅡ analysis showed that genotype-environment interaction existed in all traits, which was mainly reflected by dominance × environment. Epistasis × environment could not be ignored. The inheritance of most traits was dominated by additive effect, while the regularity was dominated by dominant effect. The elongation had no significant gene main effect. The average length and uniformity of the upper half had a positive average advantage, while the other traits had no obvious average advantage. The results of segregation analysis showed that all traits were generally controlled by major genes. The optimal genetic model for boll number per plant, lint percentage and specific strength was 2MG-EA, for boll weight and average length of upper half was 2MG-A, for uniformity and micronaire value was 2MG-AD, for elongation was 1MG-AD. The major gene heritability of boll weight, lint percentage, average length of upper half, uniformity and specific strength was higher, but the major gene heritability of boll number per plant, micronaire value and elongation was lower. The complementary results of ADAA model and major gene-polygene genetic model can effectively understand the genetic characteristics of the traits and provide a reference for the adoption of appropriate breeding improvement strategies.
Key words:upland cotton (Gossypium hirsutum L.);ADAA model;major gene-polygene;genetic analysis
棉花(Gossypium spp.)是世界上重要的经济作物,是天然植物纤维的主要来源[1]。陆地棉作为世界上应用最广泛的四倍体栽培种,种植面积已达到全球棉花种植总面积的95%[2]。产量和纤维品质作为重要的育种目标性状,明确其遗传特点,是有效进行品种改良的基础,对育种实践具有重要的指导意义。
以往对产量、纤维品质性状的遗传研究多采用经典数量遗传方法,将控制性状的多个基因作为整体分析,将遗传效应分解为加性、显性与上位性效应。Roy等[3]认为加性基因在性状表达中发挥着重要作用,对育种后代的选择具有重要价值。秦鸿德等[4]指出可通过亲本的加性效应预测杂种F1的表现,加性方差分量越大,预测精度也越高。对于产量性状,衣分被认为主要受加性效应的影响[5],而单铃质量等性状,有研究结果表明以显性效应为主[6-7],还有研究者指出其受加性、显性效应共同控制[8-9],少数研究者认为其遗传以加性效应为主[10]。对于纤维品质性状,大部分研究结果表明主要受加性效应的作用[11-13]。这些研究大多利用加性-显性(AD)模型进行分析,利用加性-显性-上位性(ADAA)模型进行遗传分析的报道相对较少。努斯热提等[14]利用ADAA模型进行陆地棉机采性状对皮棉产量的遗传贡献分析,认为不同组合皮棉产量的加加上位性效应不同,但提高这些组合加加上位性效应的主要机采性状各不相同。然而经典数量遗传分析方法无法解析出控制某一性状的单个基因的行为和效应大小。盖钧镒等[15]提出了主基因-多基因遗传体系,认为单纯的主基因或多基因控制某一性状均为特例,主基因-多基因混合遗传模型才是数量性状的通用模型。该模型将混合分布理论与数量遗传学相结合,使用群体信息,能鉴别控制某一性状的主基因数量并估计相关遗传参数,在陆地棉产量、纤维品质相关性状均有所运用[16-19]。以分子标记为手段的QTL结果也提供了控制数量性状主基因存在的证据[20]。
前人对陆地棉产量、纤维品质性状开展了大量遗传研究,但利用ADAA模型,考虑加加上位性效应的研究报道并不多见。本研究利用9个陆地棉亲本,依据NCⅡ遗传设计组配18个组合,利用ADAA模型,对不同年份下亲本、F1与F2 3世代进行遗传分析,并将F2群体主基因-多基因分析与NCⅡ分析结果相互补充,以探讨陆地棉产量、纤维品质性状的遗传模式,为进一步明确各性状的遗传规律、采用适当的育种改良策略提供理论依据。
1 材料与方法
1.1 试验地概况
试验地点为新疆阿拉尔垦区十二团试验田(北纬40°33′15″,东经81°18′57″),属暖温带极端大陆性干旱荒漠气候。该地区热量和光照条件充足,且塔里木河及胜利、上游、多浪三大水库弥补了降水量稀少的短板;年均气温10.7 ℃,无霜期200~220 d,年均日照时长2 556.3~2 991.8 h,比较适合棉花生长。生产中播种、化学调控、收获均已实行机械化,在“矮密早”与膜下精量播种种植模式下,该地区棉花平均亩产量为450~500 kg,但纤维品质指标还有待提升。
1.2 试验材料
供试材料为南疆广泛种植的9个陆地棉品种(系),包括3个早熟母本耕野王(A1)、德农0908(A2)、TH14-22(A3);6个高产优质父本J206-5(B1,国审棉2016011)、创棉518(B2,国审棉20216002)、惠民6号(B3)、创棉517(B4,国审棉20200026)、B7(B5)、中棉641(B6)。
1.3 试验方法
2018年按照3×6的NCⅡ設计组配18个组合;2019年种植亲本及F1代,亲本自交留种,并再次配制18个杂交组合;2020年种植亲本、F1与F2;2021年种植亲本与F2代。
2020年将组合A3×B5(TH14-22×B7)F1自交留种,以进行主基因-多基因试验,2021年种植亲本及F2群体。
试验采用完全随机区组设计,亲本与F1各种植2行,F2种植6行。宽窄行(10 cm+66 cm+10 cm+66 cm+10 cm)株行配置,2次重复,行长2.5 m,株距10.7 cm。每年4月中旬进行人工播种,田间管理措施参照一般大田的管理措施。
1.4 数据采集
数据采集方法为小区调查,于每年8月20日后进行农艺性状调查,亲本与F1每小区随机选取5株,F2选取10株,测定单株铃数(Bolls number),取其平均值。棉花吐絮后于霜前每小区随机收获20个(2020年F1收获15个)棉株中部棉铃,皮辊轧花后考种,测定单铃质量(Boll weight)与衣分(Lint percent)。皮棉样品编号后送新疆第一师农业科学研究所棉花质量检测中心,测定上半部平均长度(Upper half mean length)、整齐度(Fiber uniformity)马克隆值(Micronaire value)、比强度(Fiber strength)与伸长率(Breaking elongation),检测仪器型号为HVI 9000。
由于F2代为分离世代,主基因-多基因试验所有性状均采用单株数据。2021年棉花吐絮后收获127个发育正常的F2单株,进行考种和纤维品质指标测定。
1.5 数据分析
NCⅡ数据以小区平均值为单位,采用ADAA模型对不同年份的表型数据进行联合分析。各项遗传参数采用Zhu[21-22]的软件及方法进行计算。运用MINQUE(1)法估算各性状遗传效应的方差分量,用Jackknife重复抽样方法计算各遗传参数的估计值及标准误,并进行t测验。表型值分解为:
Y=μ+E+A+D+AA+AE+DE+AAE+ε
式中,μ为群体均值;E为环境效应,E~N(0,VE);A为加性效应,A~N(0,VA);D为显性效应,D~N(0,VD);AA为加×加上位性效应,AA~N(0,VAA);AE为加性×环境互作效应,AE~N(0,VAE);DE为显性×环境互作效应,DE~N(0,VDE);AAE为加加上位性×环境互作效应,AAE~N(0,VAAE);ε为剩余效应,。
利用SPSS软件对亲本及F2群体进行表型统计分析及正态性检验。根据盖钧镒[23]、王建康等[24]提出的主基因-多基因分析方法,利用王靖天等[25]开发的R软件包SEA 2.0进行分析。根据AIC准测(Akaike’s information criterion)选择备选模型并进行适合性测验(U21、U22、U23均匀性检验,Smirnov检验和Kolmogorov检验),进而确定各性状的最适遗传模型。根据最小二乘法,由各成分分布参数估算方差、加性效应、显性效应、主基因遗传力等遗传参数。
2 结果与分析
2.1 NCⅡ试验遗传分析
2.1.1 陆地棉亲本与F1、F2组合的表型分析 将9个亲本与18个F1、F2组合各性状在3年中的表型平均值列于表1。由表1可知,2019年各性状表型值F1均高于亲本,2020年除单株铃数、马克隆值、比强度F1表型值低于亲本外,其余性状均高于亲本,这表明杂交F1在8个性状上基本表现出一定的正向杂种优势。2020年各性状F2表型值存在差异,比强度高于F1但低于亲本,单株铃数、单铃质量、上半均长、整齐度、马克隆值低于亲本、F1,衣分与伸长率高于亲本。2021年除比强度F2低于亲本外,其余性状F2表型值均高于亲本表型值。亲本与F1在不同年份表现较为一致,而亲本与F2在不同年份表现不一致,如单株铃数、单铃质量与上半均长的亲本平均值在2020年高于2021年,但F2却表现出在2021年高于2020年,说明这3个性状可能存在基因型与环境互作等复杂的遗传关系。
2.1.2 遗传方差比率和遗传率分析 根据ADAA模型,产量、纤维品质性状表型方差可分为加性方差(VA)、显性方差(VD)、上位性方差(VAA)与环境互作方差,环境互作方差可分解为加性×环境互作方差(VA×E)、显性×环境互作方差(VD×E)、上位性×环境互作方差(VAA×E)以及剩余方差(Ve)。各遗传组分方差占表型方差(VP)比率及遗传率见图1。单株铃数、单铃质量、衣分、马克隆值、比强度的VA/VP均达极显著水平,无显性与上位性效应(图1a)。整齐度的VD/VP达极显著水平,无加性与上位性效应。上半部平均长度的VA/VP与VD/VP均达极显著水平,VA/VP远大于VD/VP,同时检测出不显著的上位性效应。伸长率无显著的基因主效应,受环境影响较大。从环境互作来看,衣分、整齐度有极显著的VA×E/VP。单株铃数、上半均长、比强度存在极显著的VD×E/VP。马克隆值与伸长率的VA×E/VP、VD×E/VP均达极显著水平,VD×E/VP远大于VA×E/VP。单铃质量、衣分、上半部平均长度、整齐度还存在显著或极显著的VAA×E/VP。所有性状的Ve/VP均达极显著水平,说明产量、纤维品质性状还受其他试验因素的影响。遗传率分析结果(图1b)表明,比强度有较高的广义遗传率(h2B)与狭义遗传率(h2N),遗传率达到45.2%;整齐度h2B最高,达到58.9%。与环境互作遗传率中,互作狭义遗传率(h2NE)普遍较低;伸长率与单株铃数有较高的互作广义遗传率(h2BE),分别达到61.7%与51.1%,表明这些性状在早代不同环境中选择具有一定成效。
2.1.3 参试亲本产量、纤维品质性状的遗传效应分解 在基因传递过程中,加性和加加上位性的遗传效应是相对稳定的,杂交组合的表型受亲本加性效应的影响。本研究8个性状均未檢测到显著的加×加上位性效应,因此,将亲本各性状的加性效应预测值列于表2。结果(表2)显示,B4在单铃质量有显著的正向加性效应;A1、A2、A3与B1在衣分均有显著或极显著的正向加性效应;A2、B2与B5在上半部平均长度有显著或极显著的正向加性效应;马克隆值一般认为在3.7以上时,数值越低越好,参试亲本中只有A3有极显著负向加性效应;A2、A3、B2与B5在比强度均有显著或极显著的正向加性效应。利用加性效应较高的材料作亲本,是选育出强势组合的重要途径之一,育种实践中可利用这些亲本提高单铃质量、衣分、比强度等,并适度利用马克隆值的负向加性效应将其控制在优质范围之内。整齐度未检测到加性效应,可在杂种优势利用中发挥其显性效应。伸长率加性与显性效应均不存在,宜在不同的环境中根据表型加以选择。
2.1.4 陆地棉杂交后代群体的杂种优势分析 利用ADAA模型预测基因型与环境互作的效应值及F1和F2群体的杂种优势。结果(表3)显示,基因型与环境互作在单铃质量、衣分、整齐度与比强度的F1和F2均达到极显著水平,表明这4个性状的基因型易受环境条件的影响。整齐度有极显著的正向平均优势,优势率F1为3.1%,F2为1.5%;上半部平均长度在F2有极显著的正向平均优势,优势率为1.4%;其余性状均无明显的平均优势,可见陆地棉杂交后代产量、纤维品质性状的杂种优势并不强。整齐度在F2存在极显著的正向超高亲优势,其余性状均具有极显著的负向超高亲优势。
2.2 主基因-多基因遗传分析
2.2.1 F2群体表型分布统计 对双亲及F2群体8个性状进行统计分析并构建频数分布直方图,结果(图2)显示,所有性状均呈连续分布,符合数量性状遗传。各性状变异系数范围为1.34%~41.27%,其中整齐度变异系数最小,单株铃数变异系数最大。K-S正态性检验结果表明,单铃质量、整齐度与伸长率的P<0.05,其遗传可能存在效应较大的主基因;其他性状P>0.05,符合正态分布,可能受到主基因或多基因控制。单株铃数、马克隆值与伸长率存在正向超高亲优势,比强度有正向平均优势,其他性状均表现出负向平均优势。
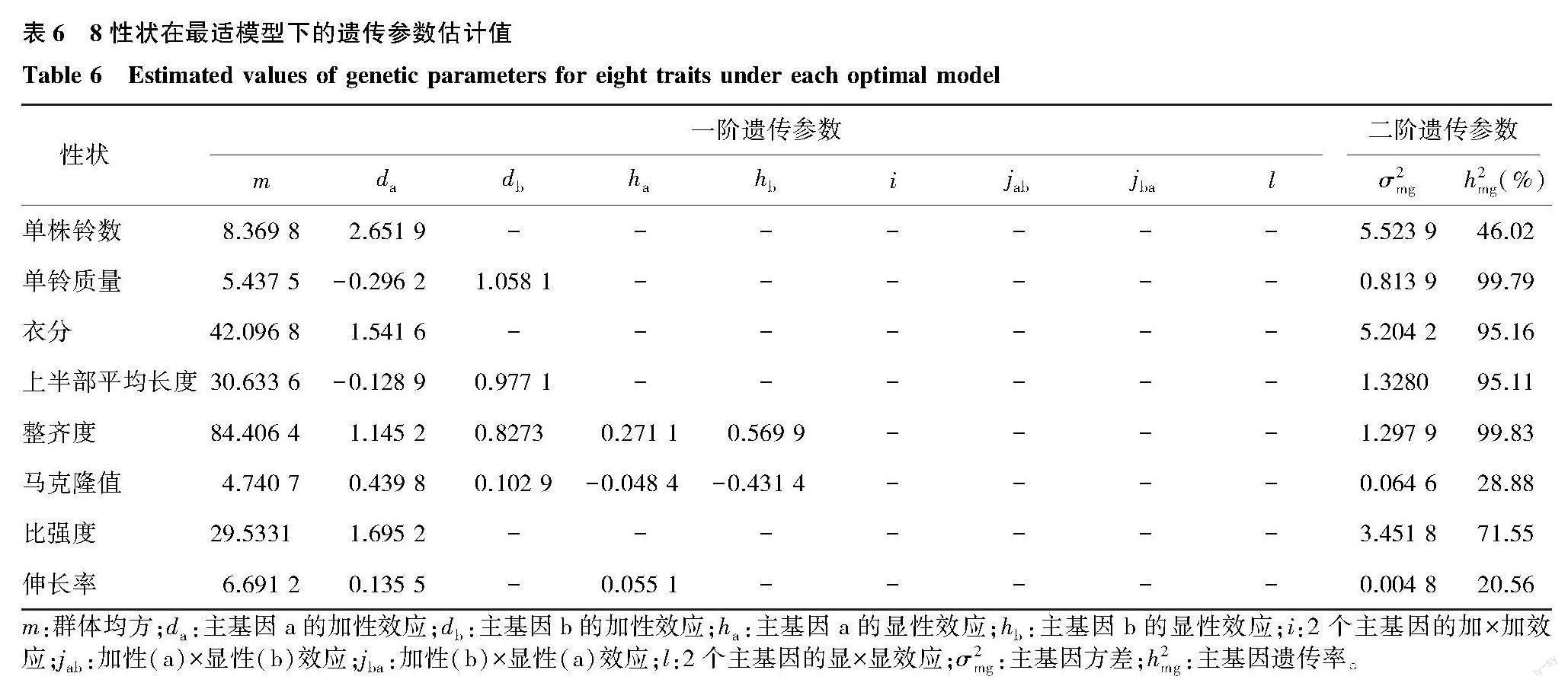
2.2.2 主基因-多基因最适遗传模型选择 对8个性状进行主基因-多基因分离分析。根据AIC准则选取AIC值最小的3个模型作为备选模型(表4)。对备选模型进行样本实际分布与最优遗传模型理论分布间的适合性检验,根据检验结果(表5)确定最适遗传模型。伸长率选择检验达到显著水平个数最少的模型为最适模型,其他性状U21、U22、U23、nW2和Dn均未达到显著,表明模型在统计学意义上有较好的拟合水平,选择AIC值最小的模型作为最适模型。表明伸长率的最适遗传模型为1MG-AD,整齐度与马克隆值的最适遗传模型为2MG-AD,单铃质量与上半部平均长度的最适遗传模型为2MG-A,单株铃数、衣分与比强度的最适遗传模型为2MG-EA。
2.2.3 主基因-多基因遗传效应分析 明确各性状最适遗传模型后,对F2群体进行遗传效应值估计,结果(表6)显示,单铃质量与上半部平均长度的第一对主基因加性效应为负向效应,其他性状的主基因加性效应均为正向增效。其中单株铃数、衣分与比强度受两对主基因控制,两对主基因加性效应值相等,分别为2.651 9、1.541 6与1.695 2。单铃质量与上半部平均长度为加性遗传,受两对主基因控制,两对主基因加性效应方向均为一正一负,正向效应均大于负向效应。整齐度与马克隆值为加性-显性遗传,受两对主基因控制,主基因加性效应均大于显性效应。伸长率符合一对主基因加性-显性遗传模型,主基因的加性效应与显性效应分别为0.135 5与0.055 1。单铃质量、衣分、上半部平均长度、整齐度与比强度的主基因遗传率较高,表明这些性状受环境影响较小。而单株铃数、马克隆值与伸长率的主基因遗传率较低,可能受到主基因与多基因的共同控制。
3 讨 论
3.1 陆地棉产量、纤维品质性状遗传分析模型的选择
以往的数量性状分析多采用AD模型,假定不存在基因的上位性效应。本试验采用ADAA模型进行遗传分析,虽各性状均未检测到显著的VAA,但单铃质量、衣分、上半部平均长度与整齐度的VAAE均达显著或极显著。由于各性状VAA均未显著,因此以AD模型再次分析,结果显示,ADAA模型与AD模型的基因主效应基本一致,但基因与环境互作项更为准确。前人[26-27]利用遗传图谱进行QTL定位并对上位性进行分析,证明上位性是陆地棉的重要遗传基础。Wade等[28]也指出上位性以及基因与环境互作对复杂遗传体系的性状要比对简单遗传体系的性状更为重要。因此,对于陆地棉产量、纤维品质性状遗传体系的探索,应充分考虑其上位性效应及其与环境互作。主基因-多基因模型能够鉴别控制某一性状的主基因数量并估计相关遗传参数,但是无法分解出基因型与环境的互作。本研究利用两种分析方法对陆地棉遗传效应进行分析,得到的结论更为可靠、全面。
3.2 陆地棉产量、纤维品质性状的遗传效应与育种策略
明确目标性状的遗传特性,才能正确地选择育种策略。遗传分析结果表明,单株铃数、单铃质量、衣分、马克隆值与比强度的遗传以加性效应为主,与前人结论[9-10]一致。整齐度以显性效应为主,这与汤飞宇等[29]、王保勤等[30]结果相符。上半部平均长度同时存在加性效应与显性效应,加性效应大于显性效应,这与李春森等[31]结论相同。伸长率无显著的基因主效应,这与前人结论不同,有研究结果[30,32]表明其以显性效应为主,也有研究者[33-34]指出以加性效应为主,这可能是由于品种或地域原因导致结果不一致。育种实践中,对于加性效应为主的性状可通过常规育种世代选择加以利用,使加性效应稳定遗传;显性效应为主的性状,则可考虑在杂种优势利用中发挥其显性效应;而对于伸长率,更适合在不同的环境中进行表型选择和改良。此外,本研究根据亲本的加性效应预测值,筛选出在各性状有较高加性效应的亲本材料,为进一步的品种改良和优势组合的测配提供了参考。
试验中所有性状均检测到基因与环境的互作效应,其中显性×环境占比较大,可见基因与环境互作主要通过显性×环境来体现,孔广超等[35]与梅拥军等[36]也有相同结论。单铃质量、衣分、上半部平均长度与整齐度还存在一定比例的上位性×环境,其中单铃质量的VAAE/VP达到9.5%,这对研究基因型与环境互作是不可忽视的,在今后的研究中需加强对上位性及其与环境互作的深入研究。对于普通狭义遗传率高的性状(比强度),可在早代进行选择;广义遗传率高的性状(整齐度),需在特定环境中加以选择。与环境互作遗传率高的性状(伸长率、单株铃数),在早代不同环境中进行选择具有一定成效。
3.3 F1、F2群体杂种优势
杂种优势是自然界普遍存在的一種现象。已有研究结果表明,棉花杂种F1可比常规种增产15%左右,并可改善品质和增强抗逆性[37],裴小雨等[38]指出F2杂种仍具有一定的利用价值。邢朝柱等[39]研究结果表明,陆地棉品质性状杂种优势不明显,产量性状杂种优势明显。本研究利用ADAA模型预测产量、纤维品质性状F1和F2的杂种优势,结果表明,仅有上半部平均长度与整齐度存在显著的群体平均优势,且仅有上半部平均长度与整齐度存在显性效应,这表明显性效应可能在杂种优势中起到决定性作用,这符合显性假说理论[40],群体超高亲优势除整齐度外均表现负向优势。F2分离群体中127个单株的统计分析结果表明,单株铃数、马克隆值与伸长率呈正向超高亲优势,比强度存在正向平均优势,其余性状均表现负向平均优势。这表明F2群体优势的实际情况与ADAA遗传模型预测值并不完全一致,实际上虽大多数性状F2出现杂种优势的衰退,群体平均值低于亲本中亲值,但仍存在超中亲优势很强的个体和组合。因此,只要F2性状表现不低于双亲,育种过程中加强对优势组合和个体的选择,利用F2的杂种优势是有可能的,这对降低棉花生产制种成本具有实际意义。
3.4 主基因-多基因分离分析
利用主基因-多基因遗传模型分析各性状遗传模式,可为后续的基因定位与分子标记辅助育种提供理论依据。分离分析结果显示,所有性状均存在一对或两对主基因,表明陆地棉产量、纤维品质性状符合主基因-多基因遗传模型,各性状普遍存在主基因,这与前人结论[41-42]一致。由于本研究所用数据来自F2群体,与前人分析世代不同,因此结果与前人存在异同。对于单株铃数,殷剑美等[43]、李成奇等[18]认为其遗传以多基因为主,而马雪霞等[19]认为其存在两对主基因,与本研究结果一致。对于单铃质量,龚举武等[44]认为一般情况下以多基因遗传为主,而本研究中单铃质量符合两对主基因加性模型,狄佳春等[17]研究结果也表明铃重在两个组合中均存在两对主基因,但不同组合主基因作用方式有所不同。龚举武等[16]认为衣分一般有两对主基因,与本研究结果一致。对于纤维品质性状,王淑芳等[45]认为多数品质性状由一个主基因和多基因控制,而艾先涛等[46]认为纤维长度与整齐度符合多基因遗传,比强度与伸长率遵循两对主基因+多基因遗传模式。本研究结果表明,整齐度与马克隆值符合两对主基因加性-显性模型,上半部平均长度符合两对主基因加性模型,比强度符合两对主基因等加性模型,伸长率符合一对主基因加性-显性遗传模型。研究结果与前人不完全一致,还需更深入的研究与探讨。不同性状主基因遗传率存在差异,单铃质量、衣分、上半部平均长度、整齐度与比强度主基因遗传率均较高,单株铃数、马克隆值与伸长率主基因遗传率较低,这与NCⅡ遗传率分析结果相似。
理论上单一分离世代可以对主基因-多基因遗传模型的基因组成进行分析,但由于不能获得误差的估计因而不能将多基因信息剖分出来,无法肯定是主基因还是主基因+多基因遗传模型,改进的方法是加入不分离群体P1、P2和F1世代,以估计试验误差[15]。今后的研究需进一步考虑加入不分离世代和回交世代进行多世代联合分离分析,进一步提高遗传模型及遗传参数的准确性,并对本研究结论进行完善。
参考文献:
[1] 王 健,董俊哲,陈 浩,等. 全球棉花进出口贸易分析及展望[J]. 棉纺织技术,2018,46(3):81-84.
[2]FANG L, WANG Q, HU Y, et al. Genomic analyses in cotton identify signatures of selection and loci associated with fiber quality and yield traits[J]. Nature Genetics,2017,49:1089-1098.
[3]ROY U, PALOTI M, TIGGA A, et al. Genetic variability studies in the F2 populations of interspecific cotton(G. hirsutum L.×G. barbadense L.) hybrids[J]. International Journal of Genetics,2019,11:660-663.
[4]秦鴻德,冯常辉,张友昌,等. 基于部分NCⅡ设计的陆地棉F1表现预测[J]. 中国农业科学,2021,54(8):1590-1600.
[5]LI B T, SHI Y Z, GONG J W, et al. Genetic effects and heterosis of yield and yield component traits based on Gossypium barbadense chromosome segment substitution lines in two Gossypium hirsutum backgrounds[J]. PLoS One,2016,11:e0157978.
[6]金骏培,武耀廷,张天真. 皖杂40杂交棉产量与品质性状的杂种优势表现及遗传分析[J]. 中国农业科学,2004,37(10):1428-1433.
[7]詹有俊,杨 涛,孙建船,等. 特早熟陆地棉的遗传效应及杂种优势分析[J]. 农业现代化研究,2012,33(4):493-497.
[8]成 磊,梅拥军,郭伟锋,等. 中熟×早熟陆地棉F1产量及形态性状的遗传分析[J]. 安徽农业科学,2008,36(15):6249-6251,6260.
[9]杨六六,刘惠民,曹美莲,等. 棉花产量和纤维品质性状的遗传研究[J]. 棉花学报,2009,21(3):179-183.
[10]JENKINS J N,WU J,MCCARTY J C,et al. Genetic effects of thirteen Gossypium barbadense L. chromosome substitution lines in topcrosses with upland cotton cultivars:II. fiber quality traits[J].Crop Science,2006,46:1169-1178.
[11]郑巨云,王俊铎,艾先涛,等. 陆地棉产量与纤维品质性状的遗传相关分析[J]. 新疆农业科学,2013,50(6):995-1002.
[12]代勇强,张新宇,孙 杰. 北疆棉花三系主要选育指标的遗传效应分析[J]. 新疆农业科学,2017,54(1):1-9.
[13]JENKINS J N, MCCARTY J C, WU J X, et al. Genetic variance components and genetic effects among eleven diverse upland cotton lines and their F2 hybrids[J]. Euphytica,2009,167:397-408.
[14]努斯热提·吾斯曼,喻树迅,范术丽,等. 陆地棉机采性状对皮棉产量的遗传贡献分析[J]. 棉花学报,2012,24(1):10-17.
[15]盖钧镒,章元明,王建康. 植物数量性状遗传体系[M]. 北京:科学出版社,2003.
[16]龚举武,刘爱英,李俊文,等. 陆地棉衣分性状的主基因-多基因遗传分析[J]. 棉花学报,2019,31(3):192-200.
[17]狄佳春,陈旭升,赵 亮,等. 利用主基因+多基因混合遗传分析解析陆地棉铃重与铃壳率杂种优势的遗传基础[J]. 棉花学报,2016,28(2):115-121.
[18]李成奇,李玉青,王清连,等. 不同生态环境下陆地棉生育期及产量性状的遗传研究[J]. 华北农学报,2011,26(1):140-145.
[19]马雪霞,丁业掌,蒋 峰,等. 亚洲棉纤维品质和产量性状的主基因与多基因遗传分析[J]. 植物遗传资源学报,2008,9(2):212-217.
[20]李成奇,郭旺珍,马晓玲,等. 陆地棉衣分差异群体产量及产量构成因素的QTL标记和定位[J]. 棉花学报,2008,20(3):163-169.
[21]ZHU J. Mixed model approaches for estimating genetic variances and covariances[J]. International Journal of Biomathematics,1992,7:1-11.
[22]朱 军. 遗传模型分析方法[M]. 北京:中国农业出版社,1997.
[23]盖钧镒. 植物数量性状遗传体系的分离分析方法研究[J]. 遗传,2005,27(1):130-136.
[24]王建康,盖钧镒. 利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应[J]. 遗传学报,1997,24(5):432-440.
[25]王靖天,张亚雯,杜应雯,等. 数量性状主基因+多基因混合遗传分析R软件包SEA v2.0[J]. 作物学报,2022,48(6):1416-1424.
[26]张先亮,高俊山,宋国立,等. 陆地棉中G6主要性状主效和上位性QTL分析[J]. 分子植物育种,2009,7(2):312-320.
[27]林忠旭,冯常辉,郭小平,等. 陆地棉产量、纤维品质相关性状主效QTL和上位性互作分析[J]. 中国农业科学,2009,42(9):3036-3047.
[28]WADE M J. A gene’s eye view of epistasis,selection and speciation[J]. Journal of Evolutionary Biology,2002,15:337-346.
[29]汤飞宇,王晓芳,莫旺成,等. 陆地棉棉铃与纤维品质性狀的遗传协方差分析[J]. 核农学报,2013,27(9):1285-1292.
[30]王保勤,刘书梅,李 宾,等. 棉花永久F2群体纤维品质性状的遗传分析[J]. 湖北农业科学,2016,55(22):5758-5760.
[31]李春森. 陆地棉品质性状遗传效应分析[J]. 新疆农垦科技,2013,36(9):3-4.
[32]石玉真,刘爱英,李俊文,等. 陆海种间杂交纤维品质性状的遗传及其F1群体优势分析[J]. 棉花学报,2008,20(1):56-61.
[33]刘 杰,曹新川,胡守林,等. 陆地棉品质性状的遗传模型分析[J]. 新疆农业科学,2015,52(7):1195-1199.
[34]詹有俊,杨 涛,孙建船,等. 特早熟陆地棉熟性产量品质的遗传相关分析[J]. 农业现代化研究,2013,34(1):118-121.
[35]孔广超,秦 利,徐海明,等. 棉花F2群体构建及其在纤维品质遗传和杂种优势研究中的应用[J]. 作物学报,2010,36(6):940-944.
[36]梅拥军,张改生,叶子弘,等. 海岛棉不同果枝品种间杂交产量性状的遗传及F1和F2群体优势分析[J]. 作物学报,2004,30(10):1026-1030.
[37]邢朝柱,靖深蓉,邢以华. 中国棉花杂种优势利用研究回顾和展望[J]. 棉花学报,2007,19(5):337-345.
[38]裴小雨,刘艳改,周晓箭,等. 陆地棉杂交组合F1、F2竞争优势研究[J]. 中国棉花,2021,48(1):16-19,41.
[39]邢朝柱,喻树迅,郭立平,等. 不同生态环境下陆地棉转基因抗虫杂交棉遗传效应及杂种优势分析[J]. 中国农业科学,2007,40(5):1056-1063.
[40]JONES D F. Dominance of linked factors as a means of accounting for heterosis[J]. Proceedings of the National Academy of Sciences,1917,3:310-312.
[41]YUAN Y L, ZHANG T Z, GUO Z W, et al. Major-polygene effect analysis of super quality fiber properties in upland cotton(G. hirsutum L.)[J]. Acta Genetica Sinica,2002,29:827-834.
[42]李成奇,郭旺珍,张天真. 衣分不同陆地棉品种的产量及产量构成因素的遗传分析[J]. 作物学报,2009,35(11):1990-1999.
[43]殷剑美,武耀廷,朱协飞,等. 陆地棉产量与品质性状的主基因与多基因遗传分析[J]. 棉花学报,2003,15(2):67-72.
[44]龚举武,刘爱英,段 丽,等. 不同环境下‘中棉所70’RIL群体棉铃重的主基因+多基因遗传分析[J]. 中国农学通报,2019,35(15):128-137.
[45]王淑芳,石玉真,刘爱英,等. 陆地棉纤维品质性状主基因与多基因混合遗传分析[J]. 中国农学通报,2006,22(2):157-161.
[46]艾先涛,李雪源,王俊铎,等. 新疆高品质陆地棉纤维品质性状遗传分析研究[J]. 棉花学报,2009,21(2):107-114.
(责任编辑:成纾寒)
