紫薇遗传图谱构建及株型性状的QTL定位
2024-01-22姜聖姬王淑安张恩亮李林芳高露璐杨如同
姜聖姬, 王淑安, 张恩亮, 李林芳, 高露璐, 杨如同, 王 鹏
(江苏省中国科学院植物研究所,江苏 南京 210014)
紫薇(Lagerstroemiaindica),又称痒痒树,是千屈菜科(Lythraceae)紫薇属(Lagerstroemia)落叶灌木或小乔木,原产于中国,至今已有1 600多年的栽培历史,以其在仲夏持久开花而闻名,并作为木本观赏植物而倍受人们喜爱[1]。自20世纪80年代限制性内切酶片段长度多态性(RFLP)分子标记技术出现以来,DNA分子标记研究逐渐成为非常活跃的研究方向[2]。分子遗传图谱记录了基因组内基因和特定的多态性DNA分子标记的相对位置关系,是基因定位和克隆的基础。自从1994年Peltier等[3]首次利用分子标记、形态标记构建了矮牵牛(Petuniahybrida)的遗传图谱后,研究者相继开展了观赏植物的相关研究。基于遗传连锁图谱的数量性状座位(Quantitative trait locus, QTL)定位能有效结合基因型与表型来鉴定植株性状,显著提高选择的准确性和育种的高效性,已在植物育种方面得到广泛应用[4]。陈丹维等[5]利用扩增片段长度多态性(AFLP)、简单序列间重复(ISSR)标记构建了蜡梅(Chimonanthuspraecox)的分子遗传图谱,为QTL定位、基因克隆等研究奠定了基础。Cai等[6]构建了第1张牡丹(PaeoniaSect.Moutan)高密度遗传图谱,并获得了控制牡丹重要性状的QTL。由此可见,构建紫薇遗传连锁图谱的研究十分必要,不仅能够发掘控制重要观赏性状的QTL,而且有利于推进紫薇属植物基因组学的研究进程。
观赏植物的株型可用来营造特定植物景观,对环境的美学价值、公园景观规划具有较大影响,在过去的40年里,关于木本植物株型QTL的研究也越来越多[7]。Lee等[8]利用共显性标记单核苷酸多态性(SNP)和微卫星构建连锁图谱,对油棕(Elaeisguineensis)的株高进行QTL定位,鉴定出8个与株高有关的候选基因。Liu等[9]结合转录组与QTL分析对沙柳(Salixcheilophila)的株高、胸径等性状进行了初步定位,在6条染色体上鉴定出6个与株型相关的QTL位点。Du等[10]利用高密度连锁图谱对杨树(Populus)株型基因进行定位,将17个QTL位点定位于11个连锁群上。Souza等[11]在橡胶树(Heveabrasiliensis)遗传图谱中对与株型相关的性状进行定位,检测到18个在夏季、冬季与其生长差异相关的QTL。目前,关于紫薇株型性状的QTL定位研究鲜有报道。
目前,研究者已经在紫薇的基因克隆、转录组分析及遗传图谱构建等方面取得重要进展。冷嘉文等[12]利用简单重复序列(SSR)标记对30个来源于美国的紫薇品种进行遗传多样性分析,探讨紫薇的遗传规律,为开展紫薇种质资源评价及新品种培育提供了理论依据。Cai等[13]对55个紫薇品种进行了指纹图谱的构建及SSR标记的开发。贺丹等[14]利用AFLP、简单重复序列(SSR)标记构建了尾叶紫薇(Lagerstroemiacaudata)与紫薇种间的遗传连锁图谱,对株高、地径等株型性状进行了复合区间QTL定位。本研究旨在借助表达序列标签微卫星(EST-SSR)标记,以紫薇金幌(LagerstroemiaJinhuang)、紫薇堇秀(LagerstroemiaJiuxiu)的F1代分离群体为作图群体,构建1张紫薇种内遗传图谱,并对紫薇的3个株型性状进行QTL定位与分析,从而为紫薇的分子育种奠定基础。
1 材料与方法
1.1 试验材料
以紫薇品种金幌(母本)、堇秀(父本)为亲本,其中父本紫薇堇秀速生,花色淡紫,叶色深绿;母本金幌生长速度慢,花色红色,叶色金色。
于2011年7-8月5:30-6:30,当紫薇母本花瓣未完全展开时去雄、套袋并将采集的父本紫薇雄蕊放置于直径90 mm的塑料培养皿中,于9:00-11:00用毛刷采集父本花粉并将其置于母本柱头上进行授粉,最后套袋,避免花粉污染。每个花序留5~10朵花。10-11月陆续收集紫薇种子。2013年3月播种,用穴盘培育后,移栽至试验田中定植。2014年春季采集金幌、堇秀紫薇杂交所得F1代实生苗200株个体的健康幼叶,于-80 ℃保存待用。引物由上海英骏生物技术有限公司合成。
1.2 DNA提取、扩增与检测
紫薇基因组DNA的提取采用十六烷基三甲基溴化铵(CTAB)法[15]。PCR扩增程序参考张恩亮等[16]的方法,SSR扩增产物的检测:8%聚丙烯酰胺凝胶电泳,210 V电泳90 min,电泳产物经银染显色完全后,统计条带数据。
1.3 紫薇株型性状的测量
在秋季新梢停止生长后,对181个F1代单株进行分枝数、地径和植株高度的调查和统计。分枝数的统计:由每个单株主干最下部第1个分枝数起,至主干末端,侧枝分枝不计数。地径的测量:使用游标卡尺在离地面高1 cm处进行测量,每个单株在不同方位进行测量,共计测量3次,取均值。高度的测量:测量每个单株从地面至主枝顶端的高度。
1.4 数据处理
数据的统计:筛选照片中有多态性的清晰的电泳条带,将有带的记为“1”,无带的记为“0”,不清楚或缺失的记为“-”。
将得到“1”和“0”组成的原始矩阵数据转换成“CP” 类型要求的数据格式,然后将数据输入Joimnap 3.0,用Kosambi函数计算图谱距离,选择对数优势比(LOD)=3.0~10.0的标记作为连锁群,构建紫薇的连锁图谱。用绘图软件MapChar 5.0绘制连锁图。用SPSS 17.0(SPSS Inc.,Chicago,USA)软件对株型性状进行统计分析,获得分枝数、地径、植株高度等性状表型值的相关系数与试验统计数据的柱形正态分布图。应用MapQTL 5.0作图软件,采用区间作图法对株型性状进行QTL分析和作图(LOD>3为阈值)。
QTL命名:q+T(目标性状)+ “-” +所在染色体(连锁群)。如果同一染色体包含同一目标性状的多个QTL位点,在染色体编号后用“-”+“数字”表示QTL数量,QTL全名通常以斜体表示[17]。
2 结果与分析
2.1 SSR引物筛选与带型分析
本研究从1 600对EST-SSR引物中筛选出337对引物,用其扩增得到清晰可辨的486.00个多态性位点。在337对多态性引物中,每对引物扩增出1.00~5.00个位点,平均扩增出1.44个位点,其中引物NBJ1050扩增的位点数最多,可以扩增出5.00个位点。
根据林木遗传作图群体的“双假测交”理论,将486个多态性位点分为以下3种类型:第1种为lm×ll分离类型(母本基因型为杂合的lm,父本基因型为隐性纯合的ll),共有208个位点,由116对引物扩增而来;第2种为nn×np(母本基因型为隐性纯合的nn,父本基因型为杂合的np)分离类型,共有204个位点,由196对引物扩增而来;第3种为hk×hk(父母本基因型均为杂合的hk)分离类型,共有74个位点,由48对引物扩增而来。
2.2 紫薇分子遗传图谱的构建
用Joimnap 3.0对486个SSR位点进行连锁分析,构建了1张包含429个SSR标记位点、分属36个连锁群的紫薇遗传连锁图谱(图1、图2)。连锁群的LOD为3.0~10.0,遗传图谱总距离为1 998.81 cM,位点间平均距离为4.63 cM。36个连锁群的平均遗传距离为55.5 cM,其中LG1连锁群的遗传距离最长,为177.6 cM,LG27连锁群的遗传距离最短,为19.3 cM。不同连锁群上的标记位点数为4~37个,最多的是LG1连锁群,有37个位点,最少的是LG17连锁群、LG19连锁群,均只有4个位点。36个连锁群中标记位点间平均间隔距离最小的连锁群是LG2,为3.2 cM,平均间隔距离最大的是LG19连锁群,为12.5 cM。在36个连锁群中发现了8个大于20 cM的间隔区域,分布在LG1(29.3 cM)、LG6(21.6 cM)、LG15(22.6 cM)、LG16(23.6 cM)、LG20(20.6 cM)、LG22(22.2 cM)、LG23(23.2 cM)和LG30(22.5 cM)上。
2.3 偏分离标记分析
经过对486个SSR多态性位点的卡方检测(χ2<χ20.01,1=2.71),发现偏分离标记位点共计124个(占比25.51%),其中位于连锁群上的偏分离标记共86个,占偏分离标记总数的69.35%,占连锁群上标记总数的20.05%。lm×ll型偏分离标记55个,占偏分离标记总数的44.35%,连锁群上标记35个,占该类型偏分离标记总数的63.64%,占连锁群上标记总数的8.16%。nn×np型偏分离标记51个,占偏分离标记总数的41.13%,其中位于连锁群上的标记38个,占该类型标记总数的74.51%,占连锁群上总标记数的8.86%。hk×hk型偏分离标记18个,占偏分离标记总数的14.52%,其中位于连锁群上的标记13个,占该类型标记总数的72.22%,占连锁群上标记数的3.03%。
2.4 紫薇株型性状的表型值分析
对181株紫薇的株高、地径和分枝数等性状进行统计和分析。从表1、图3可以看出,2个亲本金幌与堇秀在株高、地径及分枝数等性状上均存在显著差异。株高、地径和分枝数的变异范围分别是8.00~176.00 cm、0.10~1.70 cm和4.00~44.00个,变异系数分别为0.50、0.45和0.50,说明作图群体中单株间存在差异。株高、地径及分枝数的偏度均>0且<1,均为正偏离,说明3个株型性状的分布偏向母本金幌。

左侧数据为遗传图距,右侧编号为SSR位点。图1 紫薇的简单重复序列(SSR)遗传图谱(1)Fig.1 Genetic maps of Lagerstroemia indica based on simple sequence repeats (SSR) (1)

左侧数据为遗传图距,右侧编号为SSR位点。图2 紫薇的简单重复序列(SSR)遗传图谱(2)Fig.2 Genetic maps of Lagerstroemia indica based on simple sequence repeats (SSR) (2)
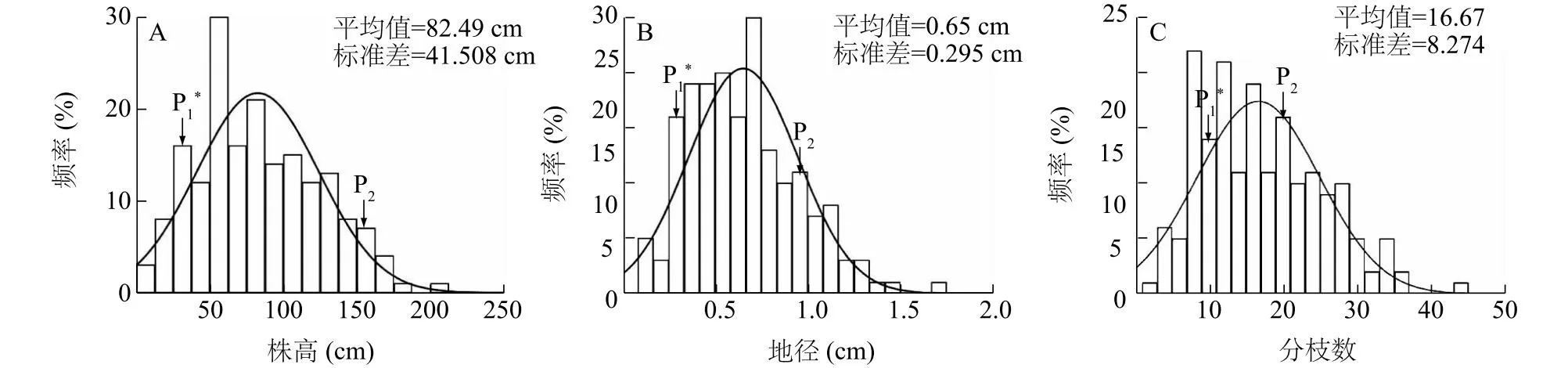
A:株高;B:地径;C:分枝数;P1:金幌;P2:堇秀;*表示P1与P2在0.05水平上有显著差异。图3 株高、地径和分枝数的频率分布Fig.3 Frequency distribution of plant high, ground diameter and branch number
2.5 株型的QTL定位
用MapQTL 5.0软件的区间作图模块对株型性状进行QTL分析,检测到4个控制紫薇植株高度的QTL位点,分别将其命名为qPH-1-1、qPH-6-2、qPH-22-3和qPH-29-4(图4、表2)。其中,qPH-1-1位于LG1连锁群上,在NBJ951-360与NBJ104-118之间,加性效应值为-1.72,来自母本金幌,可解释表型变异的4.32%;qPH-6-2位于LG6连锁群上,在NBJ1133-220和NBJ1137-235之间,加性效应值为0.23,来自父本堇秀,可解释表型变异的56.93%;qPH-22-3位于LG22号连锁群上,位于NBJ202-100和NBJ42-120之间,加性效应值为1.49,来自父本堇秀,可解释表型变异的55.61%;qPH-29-4位于LG29号连锁群上,在标记NBJ957-160和标记NBJ1133-315之间,加性效应值为-9.99,来自母本金幌,可解释表型变异的29.82%。

表1 株高、地径和分枝数的表型
本研究共检测到2个控制地径的QTL位点,分别命名为qPD-1-1、qPD-22-2,分别位于LG1、LG22连锁群上(图3、表2)。qPD-1-1在标记NBJ14-240和标记NBJ440-240之间,加性效应值为-1.33,来自母本金幌,可解释表型变异的56.43%;qPD-22-2位于标记NBJ1212-260和标记NBJ151-100之间,加性效应值为-1.72,来自母本金幌,可解释表型变异的59.02%。
本研究还发现了控制分枝数的1个QTL位点,命名为qBN-22-1,位于LG22连锁群上,在标记NBJ133-205和标记NBJ782-310之间,加效应值为-2.32,来自母本金幌,可解释表型变异的54.60%(表2)。

表2 株高、地径和分枝数3个性状的数量性状座位(QTL)分析

图4 株型的数量性状座位(QTL)定位Fig.4 Quantitative trait locus (QTL) mapping of plant architecture
3 讨 论
3.1 分子标记的选择
目前常见的分子标记大致分为4类:基于杂交的分子标记、基于PCR技术的分子标记、杂交和PCR技术相结合的分子标记及单核苷酸多态性标记[18-20]。简单重复序列(SSR)标记具有操作简便且数量丰富的特点,能够揭示较高的多态性,同时具有多等位基因特性,能够提供较高的信息量,不受外界环境干扰,检测高效快速,被广泛应用于作物、林木等植物的遗传图谱构建[20-21]。本研究以181株F1代分离群体为作图群体,利用SSR标记进行作图分析,构建了1张包含429个SSR位点的紫薇遗传连锁图谱,覆盖紫薇基因组总长度的1 998.81 cM。该连锁图谱框架结构己初步形成,并可为后续相关基因及QTL定位提供基础。
3.2 偏分离标记与遗传图谱
在DNA标记连锁图谱的制作过程中,常常会遇到大量DNA标记偏离孟德尔分离比例的现象。番茄[22]、水稻[23]、棉花[24]等作物均有关于偏分离标记在特定连锁群上集中分布或偏向于某一亲本的描述。在桃[25]、杧果[26]等多年生果树的遗传图谱作图过程中也普遍存在DNA标记的偏分离现象。Bradshaw等[27]研究发现,偏分离的标记和正常分离的标记几乎具有相同的作图效率。在本研究中,偏分离标记位点共计124个,占多态性位点总数的25.51%,其中连锁群上的偏分离标记共计86个,占偏分离标记总数的69.35%,占总连锁群上标记数的20.05%,分别低于水稻、棉花等作物的偏分离标记水平[28-29]。标记偏分离的原因可能是多方面的,如亲本遗传差异较大、分离群体偏小、致死基因的存在及亲本杂交过程中存在染色体结构重排、缺失、插入和突变等[30-35]。此外,值得注意的是,紫薇图谱中86个偏分离标记出现在12个连锁群的端部,与水稻[36]、番茄[37]、咖啡[38]等作物的研究结果相似。本研究还对部分黄化F1代植株长势、地径和植株高度3个性状进行QTL的定位与分析,推测有部分黄化F1代植株在幼年时期因叶片黄化而死亡,这可能导致SSR标记出现偏分离的现象。
3.3 QTL在连锁群上的分布特点
株高、分枝数、地径等性状是紫薇株型的重要指标。在相同环境条件下,紫薇金叶品种金幌与野生型品种堇秀在株高、地径等性状上存在显著差异,其F1代群体中在不同个体之间同样存在较大差异,这种非环境因素造成的差异必然存在分子水平的调控机制。因此,对株高、地径等性状进行QTL定位研究有利于了解其在分子水平的调控机制。
前人研究发现,株高相关的QTL位点具有较高的环境稳定性,来自父母本的等位基因均可能起到增效作用[39-41]。在本研究中,笔者鉴定了4个与株高相关的QTL位点,效应值为-9.99~1.49,其中qPH-1-1、qPH-29-4来自母本金幌,表型贡献率分别为4.32%、29.82%,qPH-6-2、qPH-22-3来自父本堇秀,表型贡献率分别为56.93%、55.61%。上述结果表明,紫薇中存在控制株高的主效QTL位点。
地径相关的QTL定位研究主要集中在木本植物中,如在杨树[42]、香榧[43]等木本植物中均检测到与地径相关的QTL位点。在本研究中,我们共检测到2个与地径相关的QTL位点,分别为qPD-1-1、qPD-22-2,分别位于LG1、LG22连锁群上,加性效应值分别为-1.33、-1.72,均来自母本金幌,分别可解释表型变异的4.32%、59.02%。上述结果说明,紫薇中存在控制地径的主效QTL位点。
张福敏[42]检测到2个与杨树萌枝数相关的QTL,均位于父本银白杨图谱上,分别为NS-1、NS-2,均位于al-2连锁群上,NS-1可解释8.25%的表型变异,2个QTL共解释了16.38%的表型变异。在本研究中,笔者检测到1个紫薇分枝数QTL位点(qBN-22-1),位于LG22连锁群上,加性效应值为2.32,来自母本金幌,可解释表型变异的54.60%,较高的表型贡献率表明该位点可能是控制分枝数的主效QTL位点。
3.4 紫薇中存在功能相关基因的紧密连锁效应
在检测数量性状QTL的过程中,QTL在分布上有区域化趋势,表现出一因多效或紧密连锁效应[44]。Kenis等[45]发现,与苹果果实性状相关的QTL集中在第10、16连锁群上。这一现象在梨[46]、玉米[47]、水稻[48]和冰草[44]等植物上均有报道。上述证据表现出一因多效或紧密连锁效应,也可以从遗传上解释不同性状间存在的相关性,进一步说明具有相关功能的基因成簇分布。对紫薇分枝数、地径与植株高度3个与生长相关的性状进行QTL定位与分析发现,3个性状中检测到的7个QTL位点集中分布在LG1、LG22和LG29等4个连锁群上,在连锁群LG1、LG22上均有4个QTL共分离或紧密连锁的现象。其中,共分离或紧密连锁的QTL数目最多的是LG22,均定位于标记位点NBJ157-145附近,说明控制同一性状的QTL位点分布在相同连锁群的不同或相邻位置。上述结果表明,可能存在与株高、地径和分枝数等性状相关的关键基因。
