Pool-ARMS:一种高效检测混合遗传样本中特定单核苷酸变异的方法
2024-01-20曹丽淼谭瑗瑗于清涛舒庆尧
曹丽淼 谭瑗瑗 汪 庆 钱 秋 于清涛 舒庆尧,5,*
(1浙江大学现代种业研究所/水稻生物育种全国重点实验室,浙江 杭州 310058;2浙江大学新农村发展研究院,浙江 杭州 310058;3无锡哈勃生物种业技术研究院有限公司,江苏 无锡 214100;4哈尔滨市农业科学院,黑龙江 哈尔滨 150029;5浙江省作物种质资源重点实验室,浙江 杭州 310058)
单核苷酸变异(single nucleotide variation,SNV),又称单核苷酸多态性(single nucleotide polymorphism,SNP),广泛存在于动植物基因组中,影响RNA 转录、剪切和蛋白质翻译及其功能。在水稻中,Wx基因第一内含子剪切供体位点SNV(G→T)使粳稻稻米直链淀粉含量降低[1-2];一个编码RNA 酶基因TMS5第71 位的一个SNV(C→A)产生了温敏雄性不育特性[3]。迄今,已有多种SNV分析方法在作物育种中应用。如将SNV 转化为基于酶切位点差异的酶切扩增多态性序列(cleaved amplified polymorphic sequence,CAPS)或衍生酶切扩增多态性序列(derived cleaved amplified polymorphic sequence,dCAPS)标记[4-5],基于DNA双链熔解特性差异的高分辨熔解曲线(high-resolution melting analysis,HRM)技术[6],以及利用DNA聚合酶延伸对引物3'端或附近与模板的错配高度敏感的特性建立的扩增阻滞突变系统PCR(amplification refractory mutation system PCR,ARMS-PCR)等[7]。其中,利用ARMS-PCR可以准确快速地对SNV进行分型,目前已成功应用于如水稻香味、光合效率相关基因[8-9]。但是,该方法只用于个体的基因分型,不能用于混合群体的突变检测。
通过诱变或碱基编辑创制特异SNV 位点是培育优异种质资源最直接有效的手段。由于诱发突变的频率极低,仅有7.5×10-6~9.8×10-6[10],美国科学家开发了一类混合样本中检测突变的技术,即定向诱导基因组局部突变技术(targeting induced local lesions in genomes,TILLING)[11],并衍生了如iTILLING(individualized TILLING)[12]、Seq-TILLING[13]、HRM-TILLING[14]等检测不同类型突变的技术体系。这些技术除基于二代测序(next generation sequencing,NGS)技术的Seq-TILLING 外,植株混合检测的倍数都很小(一般为1∶4)且检测的是各类随机突变,无法检测特定SNV。
在基因组编辑研究中,已开发基于PCR 片段分型和测序的多种突变检测技术[15]。但是,这些方法也无法对特定SNV 进行检测[16]。最新开发的引导编辑技术(prime editing,PE)可以精确地实现所有核苷酸之间的替换,但靶点的唯一性使其在特殊位点的编辑效率仍然很低,从而严重限制了该技术在植物育种中的应用[17-19]。
为了建立可以高效检出诱变和碱基编辑群体中携带目标SNV个体的技术体系,大幅降低两类群体中目标个体的检出成本,根据ARMS-PCR的基本原理,本研究设计一种适用于混合遗传样本中检测携带特定SNV个体的方法,简称Pool-ARMS。以水稻两个由单碱基变异引起的长链非编码RNA基因(long non-coding RNA,lncRNA)PMS1和PMS3为例开展实证性研究[20-22],即PMS1的一个G→T(pms1)和PMS3的一个C→G(pms3)使水稻获得了PGMS 特性,以期证明该策略的应用价值。
1 材料与方法
1.1 常规材料和PGMS水稻
本研究试验材料使用粳稻包台型细胞质雄性不育(BT-cytoplasmic male sterility,BT-CMS)保持系浙粳7B[23]的黄叶突变体浙粳7BY,浙江大学水稻生物育种全国重点实验室(本实验室)采用花药培养育成的粳型温敏不育系TS949,野败型籼稻细胞质雄性不育系保持系龙特浦B的翠绿叶突变系翠玉B(CYB)[24],聚合抗稻瘟基因Pi1和Pigm的籼稻9311的近等基因系9311pg[25],以及粳型光周期敏感性雄性不育(photoperiod-sensitive genic male sterility,PGMS)系江79S[26]。
1.2 PGMS水稻的碱基编辑
为通过碱基编辑培育PGMS 水稻,本研究采用高效代理引导基因编辑器编辑目标SNV,在pegFinder 网站(http://pegfinder.sidichenlab.org)根据pms1和pms3的序列设计pegRNA 序列[27]。每一个表达片段都有2 个sgRNA(一个nicking sgRNA 用于切割未编辑的链以提高编辑效率,另一个作为pegRNA的一部分)、一个逆转录序列(reverse transcription,RT)和一个引物结合位点(primer binding site,PBS),使用多顺反子tRNA的策略以提高多重编辑能力和效率,从而产生pegRNA和nicking sgRNA 的串联序列[28]。用于这两个基因引导编辑的pegRNA 和nicking sgRNA 序列具体信息见表1。分别合成了382和377 bp的pegRNA-tRNA-sgRNA-tRNA串联序列(重复序列tRNA为77 bp,gRNA scaffold为76 bp)。为实现双靶基因的引导编辑,将两者合成为一个759 bp的表达片段。最后,将目的表达片段导入基于潮霉素和双草醚抗性基因的双代理引导基因编辑器PE3-DS载体[19]。具体方法为:Hind Ⅲ酶切PE3-DS 载体并回收,用In-fusion 无缝克隆方法将表达盒与PE3-DS 线性化载体连接,转化大肠杆菌DH5α,菌落PCR 筛选阳性克隆。重组质粒经测序和序列比对,若正确则用于后期转化。
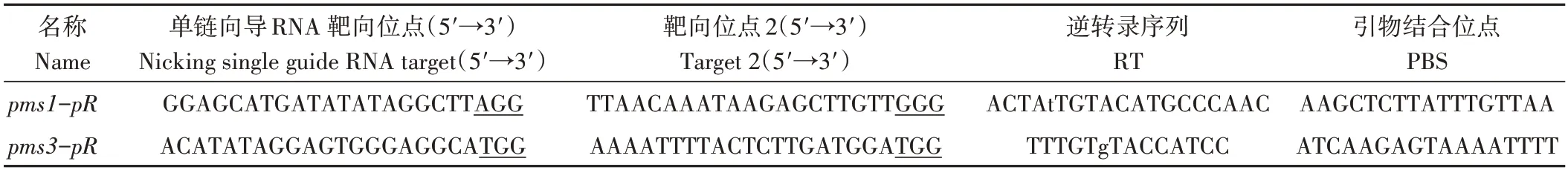
表1 用于PGMS基因PMS1和PMS3引导编辑的pegRNA和nicking sgRNA序列Table 1 pegRNA and nicking sgRNA sequences for prime editing of PGMS genes PMS1 and PMS3
从成熟胚诱导水稻愈伤组织,并用活化的携带碱基编辑重组质粒的农杆菌侵染生长状态良好的亮黄色致密胚性愈伤组织[29]。农杆菌浸染的愈伤组织在培养基上暗培养3 d后,用无菌水和含头孢的无菌水依次进行浸泡冲洗,沥干后将转到含500 mg·L-1头孢和50 mg·L-1潮霉素的第一轮选择培养基上,26 ℃、16 h 光照/8 h 黑暗条件下培养2 周。因为PE3-DS 载体是一个基于潮霉素Y46*和基于OsALSS627I组合的双代理系统,因此在抗性愈伤组织第二轮筛选培养基将潮霉素浓度提高至80 mg·L-1的同时加入0.65 µmol·L-1的双草醚(bispyribacsodium,BS),置于26 ℃、16 h光照/8 h黑暗条件下培养。之后,2 周换一次培养基,直至长出抗性愈伤组织。将抗性愈伤组织进行诱导分化和生根,获得碱基编辑的T0植株,取1~2 cm叶片用于后期检测。
1.3 扩增阻滞PCR引物设计与程序优化
采用十六烷基三甲基溴化铵(cetyltrimethylammonium bromide,CTAB)法提取水稻基因组DNA[30]。根据ARMS-PCR原理,采用Primer Premier 5.0软件设计一条与模板DNA 序列完全一致的引物,即用于扩增PMS1/pms1的pms1/R 和用于扩增PMS3/pms3的pms3/F,一条用于区别目标SNV与非目标SNV等位基因的带错配碱基的引物(图1)。设计携带错配的引物时,3'末端的最后一个核苷酸与目标SNV 一致,在3'末端倒数第二位或第三位引入一个错配碱基,错配碱基为除本身碱基外另三种碱基类型(图1)。
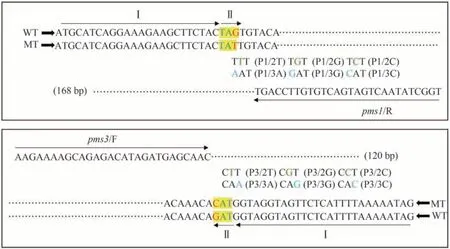
图1 两个水稻PGMS基因(PMS1和PMS3)携带SNV的片段及其ARMS-PCR引物设计示意图Fig.1 Diagram of the fragments of two rice PGMS genes PMS1 and PMS3 and their respective SNVs and the primers designed for ARMS-PCR
为确定最佳引物和扩增条件,采用梯度PCR进行扩增。PCR 体系为20 µL,体系为:引物各0.8 µL,Green Taq Mix(P131,南京诺唯赞生物科技股份有限公司)10 µL,无菌水7.4 µL,模板1 µL(200 ng·µL-1)。扩增程序为:95 ℃预变性3 min;95 ℃变性15 s,退火15 s(温度梯度为56~66 ℃,PMS1为62 ℃,PMS3为63 ℃),72 ℃延伸,共30个循环;72 ℃延伸5 min。PCR产物1 %琼脂糖凝胶电泳,拍照保存。
经检测扩增成功的PCR 产物送浙江有康生物科技有限公司进行Sanger 测序,使用DNAMAN 6.0 软件分析每个目标位点的Sanger测序图谱。
1.4 混池样本筛选SNV检测
为建立混合样本SNV检测体系,以浙粳7BY(PMS1和PMS3)和江79S(pms1和pms3)为材料,按三种方式进行DNA 混样:1)DNA 混样:先提取浙粳7BY 和江79S叶片的DNA,按1∶9、1∶19、1∶49、1∶99的比例混合成样;2)叶片混样:用1.5 mm 打孔器取单株叶片样本,再按不同比例混合制成叶片样本,提取DNA;3)芽鞘混样:将种子浸种2 d、催芽2 d以获得萌动的种子,取其胚芽鞘4 mm,再按不同比例混合制成芽鞘样本,提取DNA。叶片混样和芽鞘混样两种混合样本中江79S∶浙粳7BY的比例设为1∶1、1∶9、1∶19、1∶24、1∶49。以1.3确定的最佳ARMS-PCR条件检测目标SNV。
2 结果与分析
2.1 不同水稻材料的PMS1和PMS3基因型
为保证试验材料的准确性,先对包含PMS1和PMS3基因SNV 位点的片段测序。结果证实,江79S的SNV-pms1和SNV-pms3分别为T和G,与已报道的PGMS材料一致;其余四份材料的SNV-pms1和SNV-pms3分别为G和C,均为野生型PMS1和PMS3(图2)。

图2 不同水稻材料PGMS基因PMS1和PMS3片段序列分析Fig.2 Sequence analysis of PGMS genes PMS1 and PMS3 in different rice accessions
2.2 PGMS等位基因特异性扩增
为建立PGMS 等位基因特异性扩增体系,以浙粳7BY 和江79S 两种材料,分析了12 对PCR 引物特异性扩增光敏SNV 的效果。结果发现,在56~64 ℃温度范围内,用倒数第三位碱基错配的引物扩增可以明显区分PMS1和pms1;在61~63 ℃温度范围内,用倒数第三位碱基错配的引物扩增可以明显区分PMS3和pms3。在这两个PGMS基因位点上,用3'末端倒数第二位碱基的错配的引物区分效果均差于倒数第三位碱基错配的引物。
为确定最佳退火温度以及错配碱基的类型,对用倒数第三位碱基错配的引物区分SNV 的效果做了进一步研究。发现倒数第三位碱基错配为G的引物区分PMS1/pms1和PMS3/pms3的效果最好,最佳退火温度分别为62 和63 ℃。因此,建立了特异性扩增PGMS 等位基因pms1和pms3的引物和PCR参数(表2)。

表2 PCR扩增条件的参数Table 2 Optimized PCR amplification conditions
采用以上引物和扩增条件,多次对4份非PGMS材料进行扩增,结果均无扩增条带(图3),说明可以特异且高效检测PGMS基因。

图3 水稻PGMS基因pms1和pms3等位基因型检测Fig.3 Genotyping of two PGMS genes pms1 and pms3 in rice
2.3 混合样本SNV检测
2.3.1 叶片组织DNA样本 叶片是提取DNA最常用的材料。比较先提取DNA 再混合成DNA 池和取等量叶片组织混合后直接提取DNA 两种方法的检测效果。采用先提取再根据DNA 浓度调整后按不同比例混合形成混样DNA模板,发现以1∶99混合时仍可扩增出特异性条带(图4)。在采用打孔器从叶片中均匀取样、混合,提取的DNA 用于PCR 模板时,无论是pms1还是pms3,检出水平降为1∶49(图5)。上述方法均经过多轮、多次试验,结果均能很好重现。

图4 叶片提取DNA按不同比例混合成样本的突变检测电泳图Fig.4 Mutation detection of mixed DNA from leaves samples in different proportions using agarose electrophoresis

图5 不同比例叶片混合样本的突变灵敏度检测电泳图Fig.5 Mutation detection of mixed leaf samples in different proportions using agarose electrophoresis
2.3.2 萌动种子芽鞘组织混合DNA 样本 由于芽鞘组织量很少,保证取完胚芽鞘后的种子仍能正常生长,本研究采用先混合芽鞘组织的样本、再提DNA 的方法。多次重复试验结果表明,pms1和pms3位点的检出效率为1∶24的比例(图6)。

图6 不同比例胚芽鞘混合样本的突变检测电泳图Fig.6 Mutation detection of mixed coleoptile samples in different proportions using agarose electrophoresis
2.4 碱基编辑材料的pms1和pms3 SNV检测
本试验共获得pms1和pms3双靶点基因编辑载体转基因T0植株63株,其中CYB转化苗15株,浙粳7BY转化苗48株。采用打孔器叶片取样后提取单株的DNA,利用本研究建立的特异性PCR体系,扩增并检测目标条带。结果显示,试验组均没有扩增出特异性条带(图7)。为验证Pool-ARMS 结果的准确性,对63株植株单独提取DNA 并采用常规方法扩增目标片段,进行Sanger 测序分析。结果显示(图8),这些植株的PGMS基因与亲本一致,均为正常可育基因型,与Pool-ARMS 法确定的结果一致。

图7 两个PGMS基因碱基编辑材料的Pool-ARMS检测电泳图Fig.7 Pool-ARMS detection for the mutation of two PGMS genes in base editing plants
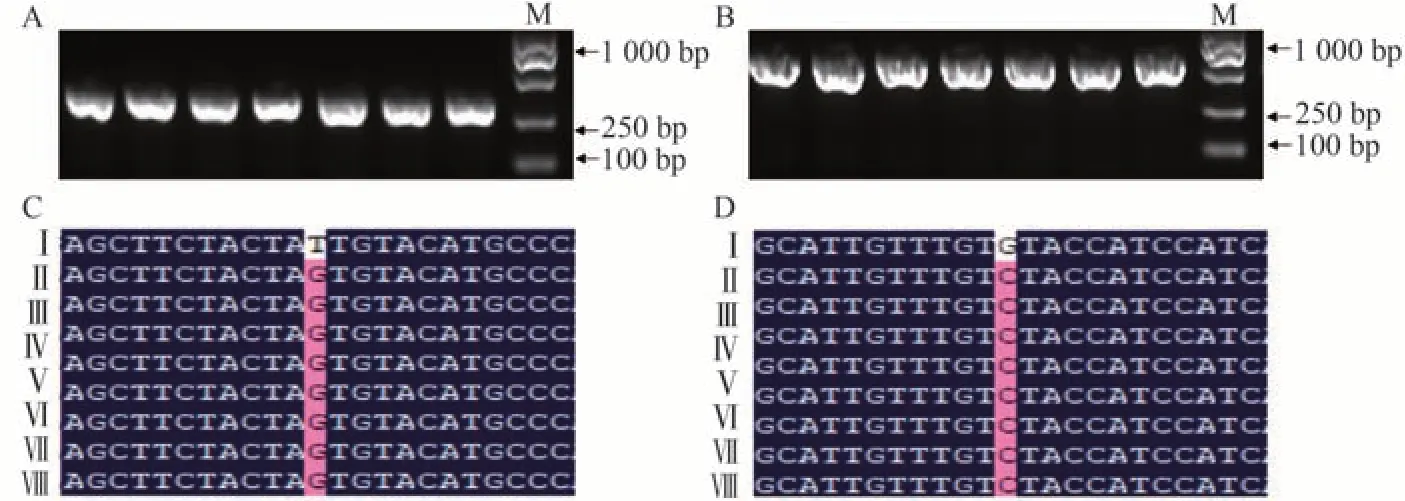
图8 两个PGMS基因PMS1(A)和PMS3(B)碱基编辑水稻T0植株的PCR片段和序列比对图Fig.8 PCR amplification and sequence alignment of two PGMS genes PMS1(A)and PMS3(B)in T0 rice plants transformed with base-editing vectors
3 讨论
ARMS-PCR 作为一种操作简便、准确性较高且低成本的SNV 分型技术,可以准确地将单碱基突变的样本进行区分,实现快速、简单、低成本和高通量的基因分型[31],具有广阔的应用前景和发展潜力。在特异性SNV 筛查中,只需确定群体中是否含有特定SNV 的个体,无需对特异和非特异SNV基因型进行分型,因此目标是建立特异性扩增目标SNV的体系。基于这一需求,本研究通过针对目标SNV 设计一条特异扩增引物,即在该引物的3'末端的倒数第三位碱基处增加一个错配,结合适宜的退火温度,实现了对群体中特异SNV的灵敏检测。
目前,多数基因检测技术都是用于个体等位基因分型,尚缺少可以用于群体中特异SNV 筛查的廉价技术体系。在医药和环境科学中,数字PCR(digital PCR,dPCR)可以对群体中稀有突变等位基因进行定量分析。但是,dPCR仪器昂贵,所需试剂耗材成本高,需要事先优化流程并研制出相应的探针,该方法在植物育种中的应用前景尚有待进一步研究。深度测序技术理论上也可以用于混池样本稀有突变的检测,但在实际应用中依赖具有生物信息分析技能的专业人员,而且该方法检出的假阳性比例较高,对筛查出的混样需要通过单个样本的进一步分析后确定[32]。因此,建立基于普通PCR 程序、操作简便快速、对样品的纯度要求较低的混样SNV 检测技术体系,具有广泛的应用前景。本研究将ARMS-PCR 的原理应用于群体SNV 的检测,建立了Pool-ARMS 体系,该方法可以大幅提高检测效率并缩小成本,为筛选特定SNV 建立了一种高效、低成本的方法,在突变群体和碱基编辑植株特定SNV筛查中具有良好的应用前景。
本研究针对实际应用场景确定了利用叶片和芽鞘组织筛选特定SNV 个体的技术体系。对水稻碱基编辑植株这样的群体,可以通过等量混合叶片样本提取DNA,开展Pool-AMRS分析,在50株T0植株的混样中检出目标SNV。对水稻诱变群体,特定SNV的比例极低,需要对高达10万个M2个体开展检测[10]。本研究建立的利用芽鞘组织的技术体系,可以实现当天取样,当天或次日出结果,从而减少田间种植、取样这一费时费力、易错的环节,大幅简化突变体筛选流程。按照1∶24 的检出能力,10 万粒种子仅需建立约4 000 个混样,进行约400 次PCR 和相应的琼脂糖电泳检测,将工作成本变得可控。
本研究建立的pms1和pms3特异SNV 检测体系主要用于混池群体的检测,同时也可以在其他育种方式中应用。例如,在水稻的花培育种中,加倍单倍体植株为纯合野生型或纯合突变型。因此,利用本研究的筛选方法,如果一个植株同时检测到pms1-SNV 和pms3-SNV,可以推定该材料即为PGMS水稻。
4 结论
本研究以碱基编辑和诱发突变的群体为目标应用场景,以控制光周期敏感性雄性不育(PGMS)水稻SNV为例,优化PCR引物和反应程序,建立了一种通过混池筛查高效检出目标SNV 的方法Pool-ARMS。利用叶片和芽鞘组织混样提取DNA 检测突变的体系,将突变检出水平分别确定为1∶49和1∶24,实现经济、高效、简便的目标,可以运用到水稻等农作物育种特别是利用诱发突变和碱基编辑创制种质资源中,实现对目标变异的定向筛选。
