缩节胺和硝普钠对棉花幼苗根际土壤酶活性及细菌群落的影响
2024-01-16吴建飞刘茜李璇温天旺杨文亭汤飞宇
吴建飞,刘茜,李璇,温天旺,杨文亭,汤飞宇
(江西农业大学农学院/ 作物生理生态与遗传育种教育部重点实验室,南昌 330045)
棉花育苗移栽是我国南方棉区传统的棉花栽培技术,与直播相比,具有保苗、节种、促早熟、延长有效开花结铃期等优点,但工序繁杂、劳动强度较大、用工较多[1-2],难以满足当前棉花栽培轻简化、机械化发展的要求[3]。 以穴盘基质育苗为代表的轻简育苗技术在优化基质配方的基础上,辅助以化学控制,在提高幼苗质量和移栽成活率、 缩短缓苗期等方面取得了显著的进展[1,4-7]。缩节胺(1,1-dimethyl piperidinium chloride,DPC)是1 种植物生长延缓剂,可阻断赤霉素(gibberellin, GA)的生物合成,在棉花上主要用于控制徒长、塑造株型[8-10]。对棉花幼苗根系发育也具有促进作用,如DPC 浸种能促进侧根发生[11],提高根系活力[12]。 苗床喷施DPC 有利于培育壮苗,促进栽后早发,具有明显的增产作用[13]。 硝普钠(sodium nitroprusside, SNP)是一氧化氮(NO)的供体,NO 是1 种重要的气体信号分子,广泛参与植物的生长发育,如促进种子萌发、抑制幼苗下胚轴伸长[14-15]、影响根系形态建成等[16-17]。在棉花幼苗1 叶1 心期,叶片涂抹500 μmol·L-1SNP 可以改善根系生长, 如提高根总长、 根体积、根尖数及根系活力等,增加根系和全株干物质质量[18]。由此可见,DPC 和SNP 均有促进棉花幼苗根系生长发育的作用。NO 通过降低生长素(吲哚乙酸,in dole-3-acetic acid, IAA)氧化酶活性减少IAA 的降解, 抑制IAA 的向顶运输,诱导生长素效应抑制蛋白的降解,从而促进侧根和不定根的发生[19]。 DPC 通过脱落酸(abscisic acid, ABA)代谢及信号转导途径,调节IAA 合成、运输及信号转导,促进中柱鞘细胞的分裂产生侧根[11]。
根际是指受植物根系及其分泌物影响的土壤区域[20],被认为是植物的第二基因组,已成为研究根系- 土壤- 微生物三者互作的一个热点领域[21]。 植物根际的物质代谢和能量转化非常活跃,根际土壤的酶活性、微生物数量及活性均高于非根际土壤[20,22]。 土壤酶来源于土壤微生物及动植物活体的分泌物及其残体的分解物,其中微生物的分泌是最主要来源[23],其活性高低能反映土壤养分的转化能力、微生物的多样性及活性等土壤生物学特征,是评价土壤综合肥力的重要指标[22-24]。 植物通过根系分泌物影响根际微生物群落的结构与功能,反过来根际微生物也通过不同的机理影响植物的生长及对环境的适应性[25-26]。例如,植物根际促生菌(plant growth-promoting rhizobacteria, PGPR)能够调节植物根系构型,影响作物生长和产量[20]。 解淀粉芽孢杆菌(Bacillus amyloliquefaciens)SQR9 可以增强黄瓜根系色氨酸的分泌及IAA 的合成,从而促进黄瓜的生长[27]。根系接种巴西固氮螺菌(Azospirillum brasilense)Sp245(能产生生长素)后,拟南芥次生根和根毛的数量增加,植株内源生长素的浓度提高[28]。 本研究中,假设DPC 和SNP 处理棉花幼苗后可能通过调节根系的分泌物影响到根际细菌的类型、多样性及活性等,进而影响土壤酶活性。 本研究利用穴盘基质育苗试验观察分析DPC 与SNP处理对棉花幼苗根系及生长的影响,及对根际土壤酶活性的影响,利用16S rRNA 测序技术分析根际细菌群落组成的差异,旨在探讨DPC 和SNP 对植物根际微生物多样性潜在的调控作用,为揭示两者促进棉花幼苗生长的机理提供新的认识。
1 材料与方法
1.1 试验设计
试验于2021 年在江西农业大学培养室内进行。 供试棉花品种为江西农业大学棉花课题组自育的陆地棉品系A201,长期自交保纯。 培养基质原材料为未耕种作物的表层红壤土, 风干过孔径2 mm 筛后与稻壳灰按体积比1︰1 均匀混合[7]。采用穴盘育苗,育苗盘为72 穴,盘体长54 cm,宽28 cm,穴深45 mm,穴上口长和宽均为40 mm,穴下口长和宽均为20 mm,容量为40 mL。 种子晒干后,经0.2%(质量分数,下同)的次氯酸钠消毒、漂洗晾干后播种,每穴1 粒种子,穴盘放置于塑料托盘内,定期适度浇水使幼苗处于适宜水分状态。 培养室条件为:温度28 ℃/24 ℃(昼/夜)、光周期14 h/10 h(光/ 暗)、光照强度180 μmol·m-2·s-1。 当棉花幼苗长至1 叶1 心时进行处理,共设置3 个处理,包括对照(CK,叶片涂抹蒸馏水),SNP 处理 (叶片涂抹500 μmol·L-1硝普钠)和DPC 处理(叶片涂抹50 mg·L-1缩节胺)。试验采用随机区组设计,每个穴盘视为1 个生物学重复,1 穴1 粒, 每个处理3 个穴盘即3 个重复。 培养条件同上。
1.2 取样
当棉花长至3 叶1 心期(约1 个月)时,在每个穴盘选取生长整齐一致的幼苗3 株。 由于基质比较疏松,根际土壤采用抖土法[29]采集,去除根部松散的土壤后,使用无菌刷子从根部收集残留的土壤即为根际土壤。 根际土壤混匀后分成2 份,一份于-80 ℃冷冻保存,用于高通量测序分析土壤细菌群落组成; 另1 份风干后过1 mm 筛用于土壤酶活性测定。 棉苗取样后清洗干净,在冰上将其分离为2 部分。 一部分为干样,调查完株高、茎粗后,将幼苗分为根、茎(含叶柄)、叶片3 部分, 经105 ℃杀青30 min 后于60 ℃下烘干至质量恒定; 另一部分作鲜样, 用Epson Expression 12000XL 扫描仪(日本长野)及时对根系进行扫描,获得根系构型参数。 每个处理取3 次生物学重复,每个重复测定2 次,取平均值。
1.3 测定项目及方法
1.3.1幼苗生长指标及根系构型参数。 利用直尺测量株高及下胚轴长度, 用游标卡尺测量茎粗(下胚轴中部直径)。 根系经Epson Expression 12000XL 扫描仪 (日本长野) 扫描后采用Win-RHIZO PRO 软件得到以下参数:根总长、根表面积、根体积、根平均直径、根尖数。
1.3.2土壤酶活性测定。 土壤脲酶活性的测定采用苯酚钠-次氯酸钠比色法; 土壤过氧化氢酶活性的测定采用高锰酸钾滴定法;土壤蔗糖酶活性的测定采用3,5- 二硝基水杨酸比色法;土壤碱性磷酸酶活性测定采用磷酸苯二钠比色法[30]。
1.3.3土壤细菌的16S rRNA 测序及多样性分析。 采用土壤专用的快速DNA 旋转提取试剂盒(Fast DNA Spin Kit for Soil,MP Biomedicals,Santa Ana,CA,USA)提取土壤样品基因组总DNA。 利用紫外分光光度计和0.1%琼脂糖凝胶电泳分别检测DNA 的浓度和质量。 采用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对16S rRNA的V3~V4 区进行扩增。 扩增产物用2%琼脂糖凝胶电泳检测,利用AxyPrepDNA 凝胶回收试剂盒(AXYGEN 公司)回收目的片段,将回收片段构建Miseq 文库,基于Illumina 平台采用双末端(paired-end)测序方法进行高通量测序。
对测序的原始序列进行质控和过滤,双端拼接后得到优化序列。 对优化序列提取非重复序列,去除没有重复的单序列,按照97%相似性对非重复序列(不含单序列)进行运算分类单元(operational taxonomic unit, OTU)聚类,获得OUT 的代表序列。 通过比对细菌分类学数据库(Silvar rRNA 数据库,http://www.arb-silva.de/)对OUT 进行细菌分类注释。 以OUT 聚类注释结果为基础, 对9 个样本进行Alpha 多样性、Beta 多样性及与土壤环境因子的关联分析。 不同处理(样本)间的比较即Beta 多样性采用主成分分析(principal component analysis,PCA)法分析,利用Adonis(称为非参数多因素方差)方法检验处理间的差异显著性, 通过偏最小二乘法判别分析(partial least squares discriminant analysis, PLSDA)进一步确认处理分组间的差异显著性。 为进一步探讨不同处理(样本)的土壤酶活性与细菌群落之间的关系,利用土壤酶活性与相对丰度前3 的优势菌群(科水平及属水平)进行冗余分析(redundancy analysis,RDA)及相关性分析。 所有分析均在美吉生物技术公司(中国上海)的微生物多样性云平台(www.major.com)进行。
1.4 统计分析
利用SPSS 20.0 软件对试验数据进行单因素方差分析,并采用邓肯多重范围检验比较各处理的平均值。使用Origin 8.5 软件作图。本试验所有数据均用“平均值±标准差”表示。
2 结果与分析
2.1 DPC 和SNP 对棉花幼苗生长及根系构型参数的影响
由表1 可知,SNP 和DPC 处理下棉花幼苗的茎粗、根干物质质量和植株干物质质量均显著高于对照;SNP 处理下棉花幼苗茎干物质质量显著高于对照,DPC 处理下棉花幼苗株高显著低于对照和SNP 处理。与对照相比,SNP 和DPC 处理的棉花幼苗根总长、根表面积、根体积和根尖数均显著增加。SNP 处理的棉花幼苗根平均直径显著高于对照,DPC 处理的棉花幼苗根平均直径也高于对照,但差异不显著(表2)。

表1 对照、缩节胺和硝普钠处理下棉花幼苗生长参数的比较Table 1 Comparison of cotton seedling growth parameters among the control, DPC, and SNP treatments

表2 对照、缩节胺和硝普钠处理下棉花幼苗根系构型参数的比较Table 2 Comparison of the root architecture parameters of cotton seedlings among the control, DPC,and SNP treatments
2.2 DPC 和SNP 对棉花土壤酶活性的影响
由图1 可知,DPC 处理的脲酶和蔗糖酶活性均显著高于对照和SNP 处理,SNP 处理的蔗糖酶活性显著高于对照,而脲酶活性显著低于对照。 DPC 和SNP 处理的过氧化氢酶和碱性磷酸酶活性与对照均无显著差异。
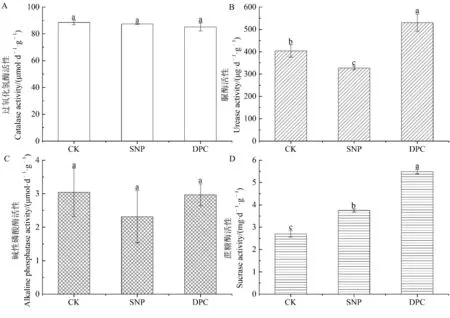
图1 对照、缩节胺和硝普钠处理根际土壤酶活性的比较Fig.1 Comparison of the soil enzyme activities in the rhizoshpere among the control, DPC, and SNP treatments
2.3 DPC 和SNP 对棉花育苗基质土壤细菌群落构成的影响
2.3.1不同处理下细菌群落组成及丰度。 不同处理下土壤细菌在门(phylum)分类水平上的相对组成如表3 所示。 相对丰度大于1%的优势种群有变形菌门(Proteobacteria)、 放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidota)、厚壁菌门(Firmicutes)、芽单胞菌门(Gemmatimonadota)、酸杆菌门(Acidobacteria)、粘菌门(Myxococcota ) 和髌骨菌门(Patescibacteria)。DPC 处理下髌骨菌门的相对丰度(1.63%)显著高于对照(0.69%),SNP 处理下髌骨菌门的相对丰度与对照相比没有显著差异。DPC 和SNP 处理下绿弯菌门、 酸杆菌门的相对丰度分别为6.50%、2.00%和7.73%、2.21%, 显著低于对照组(14.38%、5.45%)。

表3 在门水平上的相对丰度大于1%的根际土壤细菌Table 3 Bacteria with relative abundance more than 1% of rhizosphere soil on phylum level%
2.3.2不同处理下根际土壤细菌群落的Alpha多样性分析。 不同处理下细菌Alpha 多样性分析结果如表4 所示。 其中,ACE 指数(abundancebased coverage estimator)、Chao1 指数、覆盖指数(coverage index)和Sobs 指数表征微生物群落丰富度,不同处理间均无显著差异。香农-维纳多样性指数(Shannon Wiener's diversity index,简称为香农指数) 和辛普森多样性指数 (Simpson's diversity index,简称为辛普森指数)综合表征微生物群落丰富度和均匀度。DPC 处理的香农指数显著小于对照,但辛普森指数显著大于对照。SNP处理的香农指数和辛普森指数与DPC 处理和对照相比均无显著差异。

表4 对照、SNP 和DPC 处理下根际土壤细菌群落Alpha 多样性指数Table 4 Alpha diversity index of soil bacterial community in the rhizoshpere among the control, DPC, and SNP treatments
2.3.3不同处理下土壤细菌群落Beta 多样性分析和样本分组分析。 基于OUT 水平的PCA 结果(图2A)表明,不同处理的根际细菌组成存在显著差异(P=0.038),PC1 轴和PC2 轴对结果的解释度分别为20.37%和15.36%。 PLS-DA 表明SNP 处理组和对照组在COMP1 轴的方向上分离, 和DPC 处理组在COMP2 轴的方向上分离(图2B)。 利用非参数多因素方差分析进一步检测样本分组的可行性,结果显示组间差异的距离值显著大于组内差异的距离值(P=0.025<0.05)(表5),因而样本分组是有意义的,即3 个处理下土壤细菌群落组成存在差异。
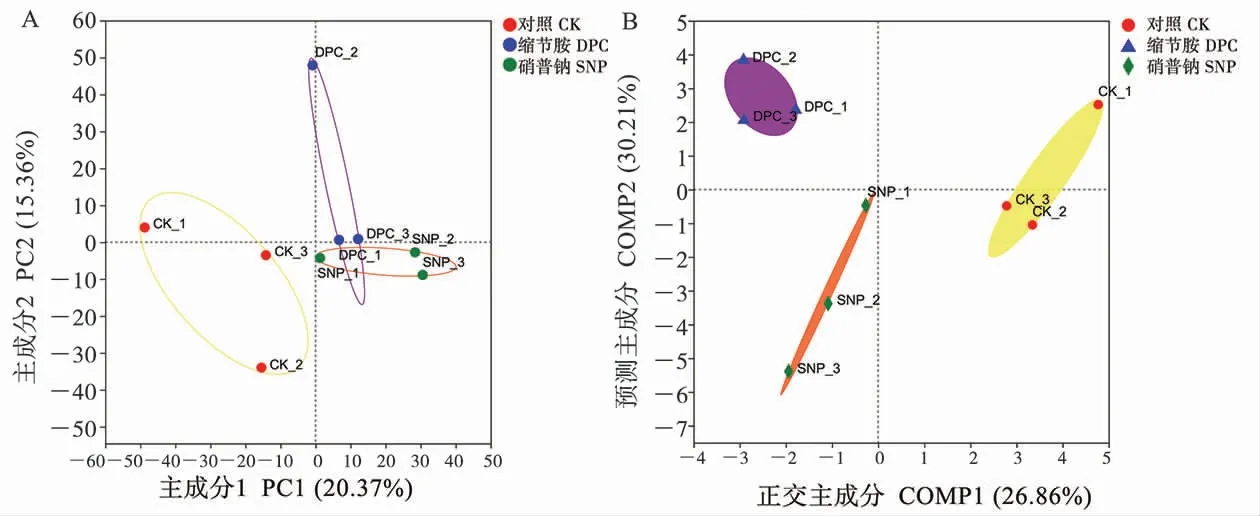
图2 不同处理土壤细菌群落的PCA(A)和PLS-DA(B)分析Fig.2 PCA (A) and PLS-DA (B) of soil bacterial community under diverse treatments

表5 非参数多因素方差分析Table 5 Nonparametric multi-variant analysis of variance
2.3.4不同处理下土壤细菌与土壤酶活性之间的关系。 为进一步探讨不同处理(样本)的土壤酶活性与细菌群落之间的关系,利用土壤酶活性与相对丰度前3 的优势菌群 (科水平及属水平)进行RDA 及相关分析。 图3 红色箭头表示4 个土壤酶活性(环境因子),3 个蓝色箭头指向相对丰度前3 的优势菌群。 菌群在酶箭头方向上的投影位置离原点越远,表示该酶活性越高、菌种丰度越高,即呈正相关,反之为负相关。 由图3A 和表6 可知, 从科水平来看, 酵母菌科(Saccharimonadaceae)细菌相对丰度与脲酶、蔗糖酶活性呈一定的正相关关系,且与脲酶活性的正相关性达到显著水平,与过氧化氢酶活性呈一定的负相关关系。 根瘤菌科(Rhizobiaceae)细菌相对丰度与蔗糖酶活性呈一定的正相关关系,与过氧化氢酶活性呈一定的负相关关系。 硝化螺旋菌科(Nitrospiraceae)细菌相对丰度与过氧化氢酶、碱性磷酸酶活性呈一定的正相关关系,与蔗糖酶活性呈一定的负相关关系。 菌种到样本的距离越近,表明该菌种在样本中的丰度越高。由图3A 可知,DPC处理组的酵母菌科细菌相对丰度比SNP 处理组和对照组高,DPC 和SNP 处理组的根瘤菌科细菌相对丰度高于对照组,对照组的硝化螺旋菌科细菌相对丰度比DPC 和SNP 处理组高。 同时RAD 表明土壤细菌群落受土壤酶活性影响程度不同,酶箭头越长,表示该酶对细菌群落分布的影响越大。 由图3A 可知,过氧化氢酶、蔗糖酶、脲酶是贡献率最大的3 个因素,说明各处理土壤细菌群落的分布主要受到该3 个土壤酶活性的影响。

图3 科(A)及属(B)水平上的冗余分析结果Fig.3 Redundancy analysis (RDA) results on family (A) and genus (B) levels
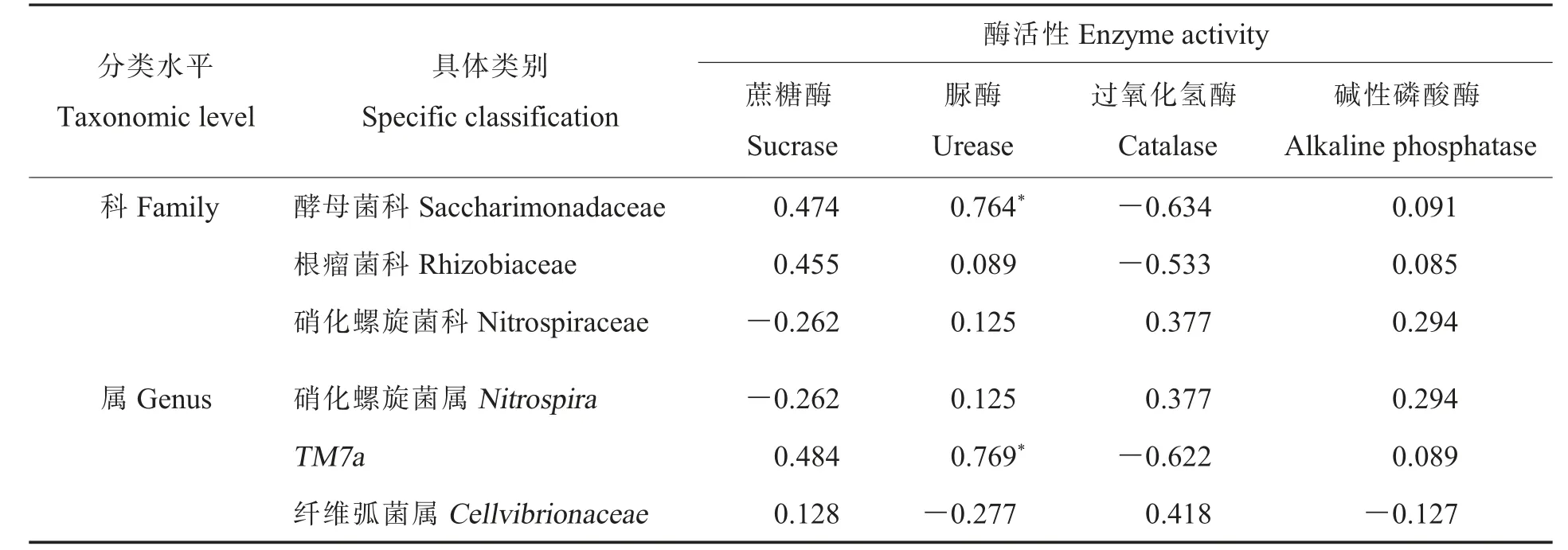
表6 细菌优势科属与土壤酶活性之间的相关系数Table 6 Correlation coefficents between bacterial dominant families and genera and soil enzyme activity
由图3B 和表6 可知,TM7a细菌相对丰度与脲酶、蔗糖酶活性呈一定的正相关关系,且与脲酶活性的正相关性达到显著水平(P=0.016),与过氧化氢酶活性呈一定的负相关关系。 纤维弧菌属(Cellvibrionaceae)细菌相对丰度与过氧化氢酶活性呈正相关关系,与脲酶活性呈负相关关系,但均不显著。 DPC 处理组的TM7a细菌相对丰度高于SNP 处理组和对照组,对照组的硝化螺旋菌属(Nitrospira)细菌相对丰度高于DPC 和SNP 处理组,SNP 处理组的纤维弧菌属细菌相对丰度高于DPC 处理组和对照组。
3 讨论
DPC 和SNP 处理促进了棉花幼苗根系的发生和生长,提高了根系及整株干物质质量。与前人报道[11,13,16-17]基本一致。DPC 处理显著提高了土壤蔗糖酶活性和脲酶活性,蔗糖酶是参与土壤碳循环的关键酶,负责将蔗糖水解为葡萄糖和果糖,为微生物繁殖提供能量,有利于微生物数量的增加[31]。 脲酶负责将土壤有机态氮转变为有效态氮,其活性增强说明土壤供氮水平提高[32-33]。 土壤酶活性的升高有利于棉花幼苗的生长。 例如,棉花与小麦、玉米等同穴互作育苗能显著提高育苗土壤中蔗糖酶、脲酶等的活性,降低土壤中有害根系分泌物的积累,进而提高棉苗素质[34]。蒜棉套作在全生育期耕作层土壤脲酶和蔗糖酶活性显著高于单作棉田,有利于土壤中细菌、放线菌的繁殖而抑制真菌增殖,说明提高棉田土壤相关酶活性提高有利于土壤健康[35-37]。
DPC 和SNP 处理后改变了棉花幼苗根际细菌群落的组成,绿弯菌门和酸杆菌门的相对丰度显著低于对照,DPC 处理的髌骨菌门相对丰度显著高于对照。 酸杆菌是土壤微生物的重要成员,通常占细菌总量的20%左右[38-39],有的超过50%[40],具有嗜酸、寡营养、难培养的特点[41]。 多数情况下土壤酸杆菌的相对丰度与土壤pH 呈显著负相关关系[39]。 DPC 和SNP 处理棉花幼苗后,有可能刺激根系分泌物质使根际营养环境改善,pH 升高,因而导致酸杆菌相对丰度下降。 绿弯菌广泛分布于生物圈各种环境,因独特的3-羟基丙酸双循环(3-hydroxypropionate bicycle,3-HP)固定CO2为人们所熟知[42],目前已发现绿弯菌参与了亚硝酸盐的氧化,即硝化作用的第二步[43-44]。 DPC 和SNP 处理后有可能导致幼苗根际周围氮素的硝化活动相对减弱。 从科及属水平分析棉苗根际细菌群落,DPC 处理下酵母菌科、TM7a菌属细菌的相对丰度高于SNP 处理和对照,对照的硝化螺旋菌科、 硝化螺旋菌属细菌相对丰度比DPC 和SNP 处理组高,SNP 处理下的纤维弧菌属的相对丰度高于DPC 处理组和对照,DPC 和SNP 处理组的根瘤菌科细菌的相对丰度高于对照组。 根瘤菌将氮素转变为植物可吸收利用的氨根离子,既有共生固氮也有非共生固氮的细菌[45]。 根瘤菌能促进豆科植物如大豆、蒺藜苜蓿侧根的发育[46-47],对非豆科植物如水稻、白三叶植株上接种根瘤菌(Rhizobiumsp.)POA3 能够增加其根系生物量[48]。接种根瘤菌IRBG74 可促进拟南芥侧根的发生,但会抑制初生根的生长[49]。 多数纤维弧菌属细菌可以合成几丁质酶,分解壳聚糖[50-51]。 某些纤维弧菌经甲壳素诱导能分泌产生多种多糖水解酶,进而分解和活化根际糖类物质[52]。 还有些纤维弧菌细菌具有固氮作用,能促进根系对氮素的吸收[53-54]。DPC 和SNP 对根系发育及整株生物量的促进作用可能分别与根瘤菌、纤维弧菌相对丰度的提高有关。 硝化螺旋菌是一类具有亚硝酸盐氧化功能的细菌,负责将亚硝态氮(NO2-)转化成硝态氮(NO3-)。对照组同时含有较高丰度的硝化螺旋菌及绿弯菌门细菌,两者都参与了氮硝化过程的第二步,说明对照组根际细菌的硝化活动比较活跃。
土壤微生物分泌物是土壤酶的主要来源[22-23]。脲酶活性与酵母菌科、TM7a菌属细菌相对丰度显著正相关,暗示提高这两类细菌的丰度有利于增强土壤脲酶活性, 从而改善土壤的供氮能力,但未检测到脲酶活性与具有硝化功能的硝化螺旋菌属的相对丰度间的相关性。 根系分泌物也是土壤酶的来源之一[55]。根系分泌物通过提供碳源、群集信号(quorum sensing)、化学引诱物调控根际微环境等影响土壤微生物生物量、群落结构及土壤酶活性[21,26,56]。棉花根系分泌物可通过促进土壤细菌和真菌的繁殖来提高土壤磷酸酶和蔗糖酶活性[57]。 下一步的工作应该是比较DPC 和SNP处理后棉花根系分泌物组成和含量的变化,分析根系分泌物、土壤酶活性、微生物组成三者之间的关系。 为从根际微生物生态学方面揭示DPC和SNP 促进棉花根系生长和幼苗长势的机理提供更深入的认识。
4 结论
棉花幼苗于1 叶1 心期叶片涂抹DPC 和SNP 可以促进棉花根系生长、提高茎粗及整株干物质质量。DPC 处理能够提高土壤蔗糖酶和脲酶的活性,SNP 处理可提高土壤蔗糖酶活性但会降低脲酶活性。DPC 处理显著提高根际土壤细菌群落的辛普森指数以及髌骨菌门的相对丰度,但会显著降低细菌群落的香农指数以及绿弯菌门和酸杆菌门的相对丰度。 DPC 处理组的酵母菌科、TM7a属细菌相对丰度比SNP 处理组和对照组高,SNP 处理组的纤维弧菌属细菌相对丰度高于DPC 处理组和对照,DPC 和SNP 处理组的根瘤菌科细菌相对丰度高于对照,对照的硝化螺旋菌科、 硝化螺旋菌属细菌丰度比DPC 和SNP 处理组高。脲酶活性与酵母菌科、TM7a菌属细菌相对丰度呈显著正相关关系。 DPC 和SNP 对棉花幼苗生长的调控可能与根际土壤酶活性及细菌群落多样性的变化存在关联性。
