广东江门古兜山保护区光叶红豆种群径级结构与空间分布格局*
2024-01-14覃俏梅梅启明吴林芳曹洪麟
覃俏梅,梅启明,吴林芳,曹洪麟,王 锋
(1.广东生态工程职业学院,广东广州 510520;2.广州林芳生态科技有限公司,广东广州 510520;3.中国科学院华南植物园,广东广州 510650;4.广东省德庆林场,广东肇庆 526600)
种群作为生态系统的基本单元,其径级结构和空间分布格局是植物种群生态学研究的重要内容之一[1]。种群径级结构是种群动态变化的反映[2],有助于揭示种群与环境之间的复杂关系,对生态系统的管理和恢复具有重要的意义[3,4]。空间分布格局是种群生物学特性和生态学过程的综合体现[5]。空间分布格局不仅受物种自身生物学特性的影响,而且受环境条件和种间竞争的直接或间接影响[6]。分析种群空间分布格局有助于理解种内、种间的相互作用以及种群分布与环境的关系[7],为生物多样性的保护和物种的保育提供科学依据[8]。
濒危植物保护研究是生物多样性保护的重要内容[9]。对濒危植物种群及其所处群落进行研究,有助于了解其濒危机制,进而提出切实可行的保育方案[10]。光叶红豆Ormosiaglaberrima为豆科Fabaceae红豆属Ormosia常绿乔木,为中国特有植物,主要分布于广东、广西、湖南、江西、海南等省区的山地、沟谷疏林中,是华南地区珍贵的用材和造林树种,具有重要的生态和经济价值[11]。由于森林砍伐、栖息地环境恶化,光叶红豆种群日渐减少,已被列为国家二级保护野生植物,并列入《世界自然保护联盟濒危物种红色名录》中,保护级别为易危(VU)[12]。目前,已有较多研究报道红豆属植物群落组成[10]、种群特征[13-15]及遗传多样性[16,17],但对光叶红豆的研究仅见种苗质量分级[12]、遗传[18]、病原菌对其幼苗更新影响[19]等方面,有关其种群径级结构和空间分布格局的研究暂未见报道。
因此,本研究以广东江门古兜山省级自然保护区内天然分布的光叶红豆种群为对象,采用典型样地法进行野外调查,利用胸径大小进行种群径级结构分析,采用点格局分析方法研究光叶红豆种群的空间分布格局,探讨光叶红豆的种群数量动态特征和空间分布格局,以期为光叶红豆种群的保护、管理与资源开发提供科学依据。
1 材料与方法
1.1 研究地概况
广东江门古兜山省级自然保护区位于珠江三角洲西南部,地理范围为112°53'11″E-113°03'25″E,22°05'00″N-22°21'5″N,总面积11 753.02 hm2,是珠三角地区面积最大的自然保护区,属于南亚热带海洋性气候,年平均气温21.7℃,最冷1 月平均气温13.3℃,最热7 月平均气温28.3℃,年降水量为1 990-2 250 mm[20]。保护区内的植被主要以季风常绿阔叶林为主,生物多样性丰富,是广东省内保存得较好的原生性次生林植被[21]。
1.2 样地设置与调查
2017年,在研究地参照Center for Tropical Forest Science(CTFS)森林大样地建设技术规范,在光叶红豆天然种群所在的常绿阔叶林群落建立1 hm2固定监测样地,样地坐标为112°57'15.64″E,22°15'03.56″N,海拔为404 m。样地为100 m×100 m 的正方形,利用全站仪划分为25个20 m×20 m的样方,样方内再细分成16个5 m×5 m 的小样方,西南角为坐标原点。对样地内胸径(DBH)≥1 cm 的木本植物进行挂牌,测定空间相对位置坐标,记录种名,测量胸径和树高等生长指标信息。2022年,参照CTFS规范,对该样地进行第2 轮次调查,再次对2017年测量过的植物个体进行测量;确认2017-2022年期间死亡(包括枯立、倒伏和消失)的个体;对5年内新增的胸径≥1 cm 的个体进行定位、测量和挂牌。
1.3 种群径级结构分析
本研究参考徐俏等[22]的方法,采用径级结构代替年龄结构,结合光叶红豆的实际生长特征,把光叶红豆个体按胸径大小分为Ⅰ级(1 cm≤DBH<5 cm)、Ⅱ级(5 cm≤DBH<10 cm)、Ⅲ级(10 cm≤DBH<15 cm)、Ⅳ级(15 cm≤DBH<20 cm)、Ⅴ级(20 cm≤DBH<25 cm)、Ⅵ级(25 cm≤DBH<30 cm)、Ⅶ级(DBH≥30 cm)共7个径级。根据野外调查情况和光叶红豆的生长特点,将Ⅰ级个体划分为幼树,Ⅱ-Ⅲ级个体划分为壮龄树,Ⅳ-Ⅶ级个体划分为成树。
各径级光叶红豆个体的死亡率、更新率和增长率采用Condit 等[23]的方法进行计算,计算公式见(1)-(3)。
其中,测量间隔时间为t,本研究中t为5年;N为群落大小;S为存活个体数量。
1.4 空间格局分析
点格局分析最早由Ripley[24]提出,可分析连续尺度上种群格局的特征,其中K(r)函数是研究植物种群空间格局最常用的方法 [公式(4)]。G(r)函数[公式(5)]由Ripley的K(r)函数修正而来,是目前评估聚集程度的重要分析方法之一。
其中,A为样地面积;n为样地内林木株数;Ir(dij)为指示函数;Wij为边缘矫正的权重;r为距离尺度,单位为m;dij为圆心i和圆心j间的直线距离,当dij<r时,取值为1,否则为0。
本研究使用单变量函数G(r)分析不同空间尺度下光叶红豆的分布格局,采用的空间尺度为0-20 m,选用完全空间随机零模型,通过运行199 次Monte Carlo拟合检验,得到95%的置信区间(即上、下包迹线内的区域)。空间格局分析中,G(r)值高于置信区间的上包迹线,表示个体聚集分布;G(r)值在置信区间之内,表示个体随机分布;G(r)值低于置信区间的下包迹线,表示个体均匀分布[24]。
1.5 数据处理
光叶红豆的个体分布点图使用R 4.2.1(http://www.r-project.org)运行程序包ggplot2 绘制[25],空间格局分析计算通过R 程序包spatstat完成[26],其他数据处理采用Excel 2021软件完成。
2 结果与分析
2.1 种群径级分析
2017年,保护区内1 hm2样地共调查到胸径≥1 cm 的光叶红豆个体216 株,平均胸径为9.04 cm;2022年,光叶红豆个体数量有较大增长,总数为297株,平均胸径为8.74 cm。从图1(a)来看,光叶红豆种群的径级结构呈倒“J”型,占比最大的是胸径5 cm之下的Ⅰ级幼树,2017 年占当年总个体数的41.67%,2022年上升至45.79%;而径级≥30 cm 的成树2017 和2022 年分别仅占当年总个体数的1.39%和1.01%,其中最大胸径为39.6 cm。死亡植株均为Ⅰ-Ⅲ级幼树和壮龄树,死亡率最高的径级为Ⅱ级;随着径级增大,个体的存活率趋于稳定[图1(b)],当径级达到Ⅳ级时死亡率降为零。由于生长进阶补充,更新率和增长率最高的径级为25 cm≤DBH<30 cm 的Ⅵ级个体[图1:(c)、(d)]。综合分析,保护区的光叶红豆以幼树为主,成树较少,整体表现为增长型,且在一定时期内种群结构相对稳定。

图1 光叶红豆种群径级分析Fig.1 Dynamics of DBH class distribution of O.glaberrima population
2.2 空间分布格局
胸径1 cm以上的光叶红豆个体在样地的中部、西南部和东部聚集程度较高;特别是2022年,在壮龄树和成树周边有较多幼树产生,个体聚集的趋势更加明显(图2)。空间分布分析结果显示,2017 年和2022年样地中所有光叶红豆个体在小空间尺度(0-6 m)上呈现较强的聚集分布;随着空间尺度的增大,聚集程度下降,在6-20 m 尺度上呈随机分布。其幼树在0-7 m 尺度上呈聚集分布的趋势,在7-20 m 尺度上呈随机分布;壮龄树在4-6 m 尺度上表现出聚集分布,在0-4 m 和6-20 m 尺度上表现出随机分布;成树在各个空间尺度上都呈随机分布;2017与2022 年的光叶红豆空间格局没有显著差异(图3)。
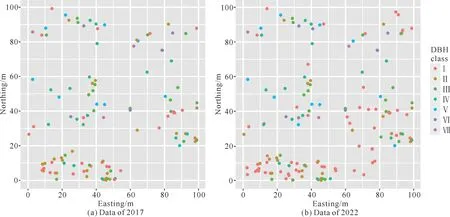
图2 不同径级光叶红豆个体的空间分布Fig.2 Spatial distribution of O.glaberrima individuals of different DBH classes

图3 不同径级光叶红豆种群的空间格局分析Fig.3 Spatial pattern of O .glaberrima population in different DBH classes
3 讨论
3.1 种群径级结构
物种自身的生物学、生态学特性及环境因子对种群特征有着重要的影响[27]。本研究中,光叶红豆种群径级结构呈倒“J”型分布,与同属南亚热带地区的广东象头山博罗红豆OrmosiaboluoensisY.Q.Wang et P.Y.Chen[10]、五桂山软荚红豆O.semicastrataHance[28]、罗浮山紫花红豆O.purpureifloraL.Chen[29]的种群径级结构相似。本研究的光叶红豆幼树个体数量多,表明种群自我繁殖能力较强,具有较大的增长潜能,这与光叶红豆结实量大有关[30]。同时,由于广东江门古兜山省级自然保护区曾是经营性的国有林场,光叶红豆等红豆属优良材用树种曾是主要的砍伐目标,导致该种群成树个体数量少。
森林更新过程中,幼苗阶段尤为关键和敏感[31,32],幼树个体数量的持续增长是种群演替的核心[33,34],对于濒危物种而言更是如此[35]。光叶红豆Ⅰ-Ⅲ级幼树和壮龄树个体存活率低、死亡率高,这可能与物种自身的生物学特性和环境条件有关。光叶红豆种子较重,扩散的距离有限[12],因此幼苗集中于母树周围,在高湿、低光强的条件下,个体间竞争激烈;随着树龄增长,个体需光量增加,个体生长严重受限[30]。另外,光叶红豆母树周围存在土壤病原菌,也抑制了幼苗个体的生长[19]。因此,针对光叶红豆种群生长特性和生境特点,保护和管理幼树个体是促进光叶红豆种群自然恢复和更新的关键。
3.2 空间分布格局
种群空间分布格局的形成受很多因素的影响[36]。自然条件下,不同尺度上各种生物因子(种子扩散机制、繁殖特性、种内和种间竞争等)和环境因子(地形、土壤、温度、水分、光照等)长期的相互作用被认为是主要驱动力[37,38]。已有研究表明,小尺度上种群的空间分布格局主要受物种的生物学特性、种子扩散机制、种内和种间竞争等因素的影响,大尺度上种群的空间分布格局主要受环境异质性的影响[39]。本研究中,光叶红豆个体在小尺度上呈现较强的聚集分布;随着空间尺度的增大,聚集程度下降,个体呈随机分布,与广东象头山博罗红豆种群的空间分布格局一致[10]。光叶红豆种群的集群分布与其生物学特性有关。光叶红豆种子呈扁圆形或长圆形,种皮坚硬,通过鸟、昆虫及风媒传播困难,离母树越远,种子越少;其幼树生长需要较多光照,一般聚集在林窗和林缘。
种群的空间分布格局在不同发育阶段也存在差异。胡尔查等[40]和Jiang等[41]的研究表明,随着植物种群径级增加,种群的聚集程度逐渐降低,空间分布格局由聚集分布向随机分布转变。本研究结果表明,在小尺度上,随着径级增加,空间分布从聚集分布趋向随机分布,光叶红豆幼树呈聚集分布,成树呈随机分布。光叶红豆种群在生命初期主要受到种子扩散限制,容易在母树周围聚集;同时,小径级种群个体生存所需的资源较少,聚集分布有助于发挥群体效应,更利于种群的稳定性。因此,光叶红豆种群初期一般为聚集分布格局。随着植株个体的生长发育,其对光照、养分、水分等生存资源的需求增大,种内、种间竞争加剧,因密度制约出现自疏和它疏现象,部分个体死亡,种群密度降低,聚集强度减弱,种群空间分布格局逐渐趋向于随机分布。
因此,本研究认为光叶红豆种群的空间分布格局在生活史初期主要由其生物学特性、生态学特性决定;随着个体的生长发育,其空间分布格局更多地受环境资源条件、种内和种间竞争的影响。激烈的竞争和密度制约是影响光叶红豆种群空间分布格局变化的重要因素。
4 结论
广东江门古兜山省级自然保护区光叶红豆种群的径级结构呈倒“J”型分布,说明种群的幼树和壮龄树数量较多,但该种群的幼树和壮龄树死亡率也较高,需针对种群的生长特性和生境特点进行幼树的保护与管理,促进光叶红豆种群高效地自然恢复和更新。在小空间尺度上,光叶红豆个体呈聚集分布,而随着空间尺度的增大,聚集程度下降,呈随机分布;随着径级增加,空间分布从聚集分布趋向于随机分布。在光叶红豆野生种群的保护与管理中,需解决其种群存在的种内和种间竞争,适当地开辟林窗、除去林下灌丛,营造适宜的栖息地,提高其就地保护成效。此外,还可以适当地对光叶红豆幼苗开展迁地保护,通过将幼苗移植到合适环境,降低密度制约的影响,提高幼苗的保存率。
