蚕豆有限生长习性的遗传分析与种质筛选
2023-12-13龙珏臣张继君王萍武云霞杜成章
龙珏臣 张继君 王萍 武云霞 杜成章







摘 要 蚕豆是重要的冷季豆类作物,目前生产中应用广泛的品种多为无限生长习性,成熟期常发生倒伏,难以满足机械化收获的要求。团队创制出有限生长习性的蚕豆材料,并通过构建F1、F2、BC1群体进行遗传分析。杂交F1表型分析结果表明,蚕豆生长习性受核基因调控,与细胞质遗传无关,且无限生长习性相对有限生长习性为显性;χ2测验结果及杂交F2群体的表型分析结果表明,蚕豆生长习性的性状受1对基因调控,且无限生长习性相对有限生长习性为显性,BC1群体的表型分析结果进一步印证了F2群体的表型分析结果。创制的有限生长习性材料为蚕豆机械化生产提供了坚实的种质保证,同时厘清了蚕豆生长习性的遗传规律,为蚕豆在生长习性方向的育种工作打下了理论基础。还筛选得到了3个适宜机械化收获的优良蚕豆种质材料。
关键词 蚕豆;生长习性;遗传分析;种质筛选
中图分类号:S643.6 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2023.17.012
无限生长习性是豆科作物的主要特点,这可以延长作物生殖生长的持续时间,在部分生殖器官受损的情况下重新获得营养和生殖器官,并形成产量[1],但无限生长习性也导致了营养生长和生殖生长对同化产物的过度竞争,使产量和收获指数变得不稳定[2]。有限生长习性作物的特征是顶端花序完成分化后茎的生长也随之停止[3],这使得株高和倒伏率显著降低,促进同化产物在营养生长和生殖生长之间更好地分配,从而获得更稳定的收获指数[1]。此外,荚果在时间和空间上更加集中,便于田间管理和机械化收获[4]。目前生产中常用的控制作物株高的方法,仍以喷施植物生长调节剂为主,但生长调节剂残留对农产品质量安全造成了巨大的隱患,还会影响后茬作物的正常生长,不利于土地周年生产。通过育种手段控制作物株高,加快农业机械化生产进程势在必行。
蚕豆(Vica fabea.L)是重要的冷季豆类作物之一,是主要的粮食和饲用作物,多年以来,育种家在蚕豆新品种选育特别是高产、高品质、短生育期品种等方面开展了大量工作,但目前生产上主推的蚕豆品种仍以无限生长习性品种为主,成熟期易发生倒伏,不适宜机械化收获;同时随着城镇化进程的加快,我国农村劳动力短缺现象日渐显现,并逐渐成为限制蚕豆产业发展的关键因素。为应对目前出现的蚕豆机械化收获难题,笔者通过大量工作创制了一批有限生长习性的蚕豆品系,为打破生长习性的制约提供了物质基础。目前关于蚕豆有限生长习性的遗传规律研究较少,因此本研究通过课题组现有材料构建了F1、F2和BC1群体,对蚕豆生长习性的遗传规律进行探究。
1 材料与方法
1.1 试验材料
本研究采用的是课题组自有的稳定亲本材料:GS-40(有限生长习性)和2016-327(无限生长习性)。通过GS-40×2016-327的正、反交构建F1、F2群体和BC1群体。GS-40为有限生长习性,结荚位靠植株上部,集中在顶端,株型紧凑直立。2016-327为无限生长习性材料,结荚位在中上部,株高较高。
1.2 试验方法
亲本材料于2018年秋季种植在隔离大棚中,2019年春季配置GS-40×2016-327正、反交组合,并于当年获得正、反交F1群体。2019年秋季将获得的F1群体种植于隔离大棚令其自交,于2020年春季开展回交,当年获得F2群体及BC1群体。在各世代成熟期前,对顶端分化情况进行观察判定其生长习性,应用遗传学方法记录各群体生长习性的分离比,并应用χ2测验进行分析。
1.3 遗传数据分析
本研究中χ2值的计算公式:
式(1)中,Ai为F2代第i个分离性状观测值,npi为预测F2代第i个分离性状预期值,k为分离性状总个数。采集的分离性状数据在EXCEL中进行统计,χ2测验采用SPSS软件进行。
1.4 种质筛选
2021年将课题组现有的76个有限生长习性蚕豆材料按单株种植于重庆市农业科学院渝西作物实验基地,每个单株形成株行种植,行距50 cm,穴距25 cm,每穴2苗。参考《蚕豆种质资源描述规范和数据标准》进行记载,于成熟期对各株行的荚质、荚姿、株高、分枝数、主茎节数、单株荚数、单荚粒数、百粒重及株行产量进行考察(见表1)。从中筛选高产、软荚的有限生长习性的优质种质材料。
2 结果与分析
2.1 亲本稳定性检验
本研究中的2个亲本材料(GS-40和2016-327)在2017年分别通过隔离自交获得自交F1群体,2018年考察F1群体表现生长习性。结果表明,GS-40自交F1群体共计107个,均表现为有限生长习性;2016-327自交F1群体共计143个,均表现为无限生长习性。通过稳定性测验,表明本研究中的亲本遗传稳定性良好,可用于后期遗传分析配置杂交组合。
2.2 杂交F1群体生长习性表现
通过对GS-40×2016-327正反交F1群体的生长习性表现进行观察统计发现,正反交获得的所有F1个体均表现为无限生长习性,正反交结果一致。这表明蚕豆生长习性受核基因调控,与细胞质遗传无关,且无限生长习性相对有限生长习性为显性(表2)。
2.3 杂交F2群体生长习性表现
于始荚期对F2群体的生长习性进行统计,其中GS-40×2016-327组合共获得292个单株,有限生长习性的有64个,无限生长习性的有228个,分离比为1∶1.36。2016-327×GS-40组合共获得217个单株,有限生长习性的为58个,无限生长习性的为159个,分离比为1∶2.7。χ2检验结果表明,两个组合群体的分离比均小于[χ20.05]=3.84,因此通过χ2检验可确定两个组合的分离比符合1∶3。这表明,生长习性的性状受1对基因调控,且无限生长习性相对有限生长习性为显性(见表3)。
2.4 BC1群体生长习性表现
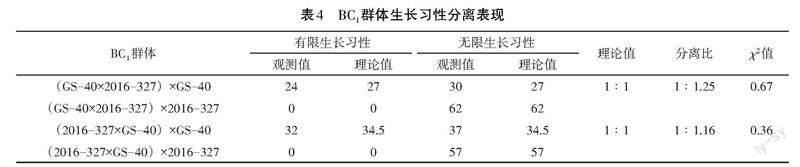
利用有限亲本和无限亲本分别与正反交F1群体展开回交获得BC1群体,其中(GS-40×2016-327)×GS-40获得54个单株,其分离比为1∶1.25;(GS-40×2016-327)×2016-327获得62个单株,均为无限生长习性。(2016-327×GS-40)×GS-40获得69个单株,分离比为1∶1.16;(2016-327×GS-40)×2016-327获得57个单株,均为无限生长习性。通过χ2检验对BC1分离群体的分离比进行测验,结果显示均小于[χ20.05]=3.84,分离比符合1∶1。这表明有限生长习性不受细胞质基因调控,而受1对核基因调控,且无限生长习性为显性,也进一步证明了F2群体中得到的结果(见表4)。
2.5 种质筛选
于成熟期对76个株系进行了室内考种,结果表明,单株产量在30 g以上的有17个材料,百粒重在90 g以上的材料有23个,适宜机械化收获的软荚材料共23个,底荚高度大于15 cm的材料26个;其中同时满足以上条件的材料有3个,分别为YS-1、YS-13和YS-56,这3个种质材料产量高,又适宜机械化收获(见表5)。
3 小结与讨论
有学者认为生长习性是受少数基因控制的质量性状[5-6],关于大豆的研究中,有限生长习性的材料与无限生长习性的材料杂交后,F1代表现为无限生长习性,这表明在大豆中无限生长习性相对于有限生长习性表现为显性,这一点与本研究结果是一致的[7]。笔者猜想,这一现象是否在豆科作物中普遍存在,还需要进一步试验验证。本研究结果表明蚕豆有限生长习性受单基因调控,这与大豆的相关研究结果一致[7]。有的作物生长习性还会受光周期影响,在光照不足或光周期明显缩短的情况下,表现出明显的无限生长特性;而光周期适宜时,表现为有限生长习性[8-10]。本课题组在长期实践中并未发现蚕豆的生长习性受光周期调控的案例,在对不同纬度的适应性鉴定中,仅表现为株高和节间长度的差异。针对这一差异,课题组将持续关注并开展相关研究。
蚕豆生长习性的相关研究目前还处于起步阶段,生产中的大部分蚕豆品种为无限生长习性,机械化收获难题亟待解決。随着农业现代化发展加快,农村劳动力短缺严重,机械化生产必然是农作物收获的主要方式。目前,缺乏适宜机械化收获的品种是限制蚕豆产业高质量发展的一个主要问题。本研究通过构建遗传群体对蚕豆生长习性进行了遗传分析,厘清了有限生长习性的遗传规律。同时本研究中的有限生长习性亲本材料可以作为稳定的品系改良成为适宜机械化收获的品种,为蚕豆机械化收获打下物质基础。此外,筛选创制了一批有限生长习性的材料,从中筛选得到3个具有重大开发价值的种质材料,将在今后的蚕豆产业宜机化发展进程中扮演重要角色。
本研究采用的亲本材料(GS-40和2016-327)系稳定材料,可用于后续研究。杂交F1群体生长习性均表现为无限生长习性,这表明蚕豆生长习性受核基因调控,与细胞质遗传无关,且无限生长习性相对有限生长习性为显性。杂交F2生长习性的分离比符合1∶3,表明蚕豆生长习性的性状受1对基因调控,且无限生长习性相对有限生长习性为显性。2个BC1群体的生长习性分离比均符合1∶1,这表明有限生长习性不受细胞质基因调控,而受1对核基因调控,且无限生长习性为显性,F2群体的分离结果和BC1群体的分离结果相互印证。综上所述,蚕豆有限生长习性受1对核基因调控,而不受细胞质基因调控,且无限生长习性为显性。筛选得到3个适宜机械化收获的优良材料,分别为YS-1、YS-13和YS-56。
参考文献:
[1] AVILA C M, ATIENZA S G, MORENO M T, et al. Development of a new diagnostic marker for growth habit selection in faba bean (Vicia faba L.) breeding[J]. Theoretical and Applied Genetics, 2007, 115(8): 1075-1082.
[2] HUYGHE C. Genetics and genetic modifications of plant architecture in grain legumes: a review[J]. Agronomie, 1998, 18: 383-411.
[3] FILIPPETTi A. Inheritance of determinate growth habit induced in Vicia faba major by ethyl methane sulphonate (EMS)[J]. Fabis News, 1986, 15: 12-14.
[4] 范淑英,张彦斌,谷成日,等.摘心对菜用蚕豆生长的影响[J].长江蔬菜,2006(3):43.
[5] BERNARD R L. Two genes affecting stem termination in soybean[J]. Crop Science, 1972, 12: 235-239.
[6] 胡喜平,赵海红,郭泰,等.半矮秆大豆育种组配方式的研究[J].大豆科学,2009,28(5):935-937.
[7] LIU B, WATANABE S, UCHIYAMA T, et al. The soybean stem growth habit gene Dt1 is an ortholog of arabidopsis TERMINAL FLOWER1[J]. Plant Physiology, 2010, 153: 198-210.
[8] 吴存祥,刘金,李兴宗,等.扁茎大豆的花序形态受光周期调控[J].中国油料作物学报,2004,26(1):36-41.
[9] LI X M, WU C X, MA Q B, et al. Morphology and anatomy of the differentiation of flower buds and the process of flowering reversion in soybean cv. Zigongdongdou[J]. Acta Agronomica Sinica, 2005, 31: 1437-1442.
[10] XU M, XU Z, LIU B, et al. Genetic variation in four maturity genes affects photoperiod insensitivity and PHYA-regulated post-flowering responses of soybean[J]. BMC Plant Biology, 2013, 13: 91.
(责任编辑:易 婧)
收稿日期:2023-05-13
基金项目:重庆市农业科学院绩效激励引导专项项目(cqaas2021jxjl05);国家产业技术体系(CARS-08);重庆市技术创新和应用发展专项重点项目(cstc2021jscx-gksbX0011);重庆市作物种质永川豆类高粱荞麦资源库(ZWZZ2020012)。
作者简介:龙珏臣(1991—),男,重庆合川人,硕士,副研究员,主要从事豆类作物遗传育种及植保研究。E-mail: longjuechen@163.com。
*为通信作者,E-mail: duchengzhang@qq.com。
