水热互作对多年冻土区高寒沼泽草甸土壤温室气体排放的影响
2023-12-04杨文丹赵秋梅翟泰雅毛天旭
杨文丹, 赵秋梅, 翟泰雅, 何 燕, 毛天旭, 张 涛
(1.贵州大学农学院, 贵州 贵阳 550025; 2.贵州民族大学生态环境工程学院, 贵州 贵阳 550025; 3.贵州大学林学院, 贵州 贵阳 550025; 4.贵州大学新农村发展研究院, 贵州 贵阳 550025)
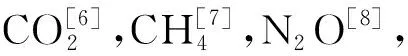
青藏高原是全球中、低纬度海拔最高,面积最大的多年冻土分布区,也是气候变化最敏感的地区之一[10],其变暖速率是全球陆地变暖速率的2倍[11]。气候变暖引起青藏高原冰川退缩、积雪融化和冻土退化,显著特征是多年冻土温度升高和活动层增厚[12-13]。多年冻土退化和活动层增厚必将导致多年冻土中大量赋存的地下冰转化为液态水,从区域尺度来看,1980—2018 年青藏高原多年冻土区降水和土壤含水量呈现显著增加趋势[12],为期55年的降水观测发现,青藏高原主体地区年降水量以6.24 mm每10年的趋势增加[14]。土壤水分含量决定了土壤氧气含量和氧化还原水平,对温室气体排放产生重要影响[15]。青藏高原多年冻土区呈现变暖和变湿的特征[16],这将加速微生物分解土壤有机碳和氮矿化过程,进而显著影响青藏高原多年冻土区温室气体排放过程[2-3,17]。高寒沼泽草甸是青藏高原上分布最广的草地生态系统之一,面积约5×104km2,占青藏高原高山湿地的79.8%,土壤有机碳储量达19.8×102Pg[18],其独特的自然环境条件及复杂的生物学过程在温室气体排放和吸收中发挥着重要作用[19]。
温度和水分是影响高寒沼泽草甸温室气体排放的主要环境因子[20],温度升高会导致土壤蒸散量增加,减少土壤水分,增加氧气可用性,加速土壤有机质分解,进而促进CO2排放[21-22],但也有研究表明,土壤处于淹水状态仍会促进CO2排放[23]。水分通过产生厌氧条件影响甲烷氧化菌群落,并改变CH4通量的气体扩散速率[24]。随着温度的季节性升高,CH4排放量也会显著增加[25],气候变暖通过增加土壤表层产甲烷菌数量和丰度来增加CH4排放[26],然而,也有研究发现,受土壤水分限制,增温会减少CH4排放[18,27]或没有显著影响[28]。N2O排放随着温度的升高而显著增加[3],也有研究发现,增温对N2O排放没有影响[29]或减少N2O的排放[30],在一定含水量范围内,硝化速率随水分增加而增加,当供氧不足时,反硝化速率增加,导致N2O排放减少,当土壤充水孔度为50%~60%时,土壤硝化速率最大,淹水条件将降低N2O排放[31]或没有显著影响[32]。可见,生态系统温室气体排放受水热双因子驱动,其通量存在很大的不确定性。
目前关于青藏高原草地温室气体排放开展了大量研究[28,33],但尚不明确水热互作对多年冻土区沼泽草甸土壤温室气体排放的影响,特别是气候暖湿化影响沼泽草甸季节性淹水过程,导致土壤在干燥、湿润和淹水间动态变化,进而影响土壤温室气体排放。本研究通过室内交互培养试验,对比不同水热条件对多年冻土区高寒沼泽草甸土壤CO2,CH4和N2O三种温室气体通量的潜在影响,为青藏高原多年冻土区草地土壤温室气体对气候变化的响应提供一定理论依据。
1 材料与方法
1.1 研究区概况
供试土壤取自青藏高原风火山典型多年冻土区(92°50′~93°3′ E,34°40′~34°48′ N),该地区海拔4 600~4 800 m,年均温为-5.3℃,年降水量310.7 mm,其中80%降雨集中在生长季。该地区冻土厚度约50~120 m,活动层厚度约0.8~1.5 m。植被类型以高寒草甸和高寒沼泽草甸为主,高寒沼泽草甸主要优势物种为藏嵩草(KobresiatibeticaMaximowicz)、异针茅(StipaalienaKeng)和羊茅(Festucaspp)[34]等高寒植物。土地类型属高寒草甸土[35-36]。
1.2 样品采集
于2020年8月生长季旺盛期,在沼泽草甸土随机选择3个50 m×50 m植被覆盖均匀的样地,代表3个重复,用5点法进行采样,3个样地均用直径为5 cm土钻采集0~10 cm表层土,每个样地采集5钻土样并充分混匀,去除可见根系后过2 mm不锈钢筛,将过完筛的土样装入自封袋,保存在装有冰袋的保温箱里,然后存放于4℃冰箱待用。
1.3 试验设计
采用双因子试验设计,设置5℃,10℃,15℃(记为T5,T10,T15)3个温度梯度和75%,100%,130%土壤持水力(Water holding capacity,WHC,分别记作W75,W100,W130)3个水分梯度进行室内交互培养试验,共9个处理,每个处理3个重复。每个均质样品100 g放入容积为250 mL聚乙烯塑料瓶中,喷洒去离子水调整到培养所需的75%,100%,130% WHC含水量。培养开始前,土壤放入15℃恒温培养箱中预培养3天,以激活土壤微生物活性。预培养结束后,将土壤分别置于5℃,10℃,15℃恒温培养箱(LRH-100CA,上海),在暗环境下好氧培养90天,培养期间采用称重法每3天补水一次,以保持土壤水分恒定。
1.4 温室气体采集与测量
参考McDaniel等[37-38]的方法,在培养1,2,3,4,5,7,10,15,20,25,30,40,50,60,75,90天采集测量气体样品。用装有三通阀的气密盖密封瓶子,使所有瓶子内的空气流通,使用空气压缩机用新鲜空气彻底冲洗瓶子20 min后,立即使用带有三通阀的注射器收集气体样品(20 mL),调节三通阀气密盖密封,60 min后采集第2次气体,采集气体间隔时间均为60 min。采气之后打开瓶盖,直到下一个采样日期。使用气相色谱仪(GC-2014,日本)对采样气体进行测定分析。CO2和CH4为FID (氢火焰离子检测器),柱温55℃,检测器温度200℃,以高纯N2(纯度99.999%)作为载气,H2作为燃气;N2O为ECD(电子捕获检测器),柱温为55℃,检测器温度350℃,以高纯N2(纯度99.999%)作为载气,待测气体浓度通过标准气体和待测气体的峰面积计算得到。3种温室气体同时测定,每个样品测定时间为7.5 min。温室气体排放/吸收(CO2,CH4和N2O)速率F(μg·kg-1·h-1)由密封前后样品浓度差算得,计算公式如下[39]:
式中,ρ为气体标准状态下的密度(kg·m-3);dc/dt为单位时间培养瓶内气体浓度增加量(ppm·h-1);V是培养瓶中气体体积(m3);m为培养瓶内干土质量(kg);T为培养温度(℃)。
温室气体累积排放量计算:
式中:CE表示CO2,CH4,N2O的累积排放量(mg·kg-1)或(μg·kg-1);F为温室气体排放通量(μg·kg-1·h-1);w为两次采气时间间隔(d);n为采样次数。
全球增温潜势计算公式:
GWP=CECO2+CECH4×27+CEN2O×273
式中,CH4和N2O所引起的全球增温潜势(GWP/mg·kg-1,以CO2当量计)分别是CO2的27倍和273倍[1]。
用范特霍夫方程计算土壤温室气体排放的温度敏感指数Q10:
Re=α×eβ×T
Q10=e10β
式中:Re为温室气体排放速率(μg·kg-1·h-1);a,b为回归参数;T为培养温度(℃)。
1.5 土壤理化性质测定
土壤pH和电导率(EC)采用水浸提,称5 g的土加25 mL蒸馏水(土水比1∶2.5),充分搅拌均匀后静止30 min,用酸碱度测试仪(PHS-3C,中国)测定;土壤微生物生物量碳(MBC)和土壤微生物生物量氮(MBN)采用氯仿熏蒸提取法[40](Vario TOC,德国);有机质(SOM)采用重铬酸钾氧化——外加热法,过氧化氢酶(Catalase)采用KMnO4滴定法测定[41],土壤蔗糖酶(Invertase)和脲酶(Urease)采用分光光度法测定[42],3,5-二硝基水杨酸比色法测定蔗糖酶,苯酚-次氯酸钠比色法(GEN10S UV-VIS,USA)测定脲酶。
1.6 统计分析
使用Excel 2019和SPSS 26.0软件进行数据统计分析。采用Two-way ANOVA分析温度、水分及其交互作用对温室气体排放量及全球增温潜势的影响。采用指数模型、二次项模型、和二元线性模型分别拟合温度、水分及二者交互作用与温室气体累积排放量关系。采用最小显著差数检验法(LSD法)分析各处理间的显著性差异。采用Pearson相关分析温室气体排放与土壤因子的相关性。方差分解分析由RStudio“Vegan”包中的varpart函数实现。显著性水平均定义为P<0.05。
2 结果与分析
2.1 温室气体排放的时间变化特征
W75处理各温度梯度下CO2排放速率随着培养时间增加呈先升高后降低,最终趋于平稳趋势(图1a),W100处理T15条件下CO2排放速率随着培养时间增加快速降低后趋于稳定,而T10和T5缓慢降低(图1b),W130处理CO2排放速率则表现出相反趋势,各温度梯度下随着培养时间增加呈逐渐升高趋势(图1c)。W75和W100处理各温度梯度下CH4排放速率随着培养时间增加上下浮动(图1 d~1e),W130处理,培养前15天各温度梯度下CH4排放速率平稳,20天后,T15条件下CH4排放速率显著提升,在第60天达到峰值后立即下降(图1f);不同温度和水分处理下N2O排放速率存在较大差异,W75和W130处理各温度梯度下N2O排放速率没有明显规律(图1g~1i),W100处理,T15和T10条件下N2O排放速率分别在培养15和30天出现最大峰值(图1h),而T5条件下一直趋于平稳。
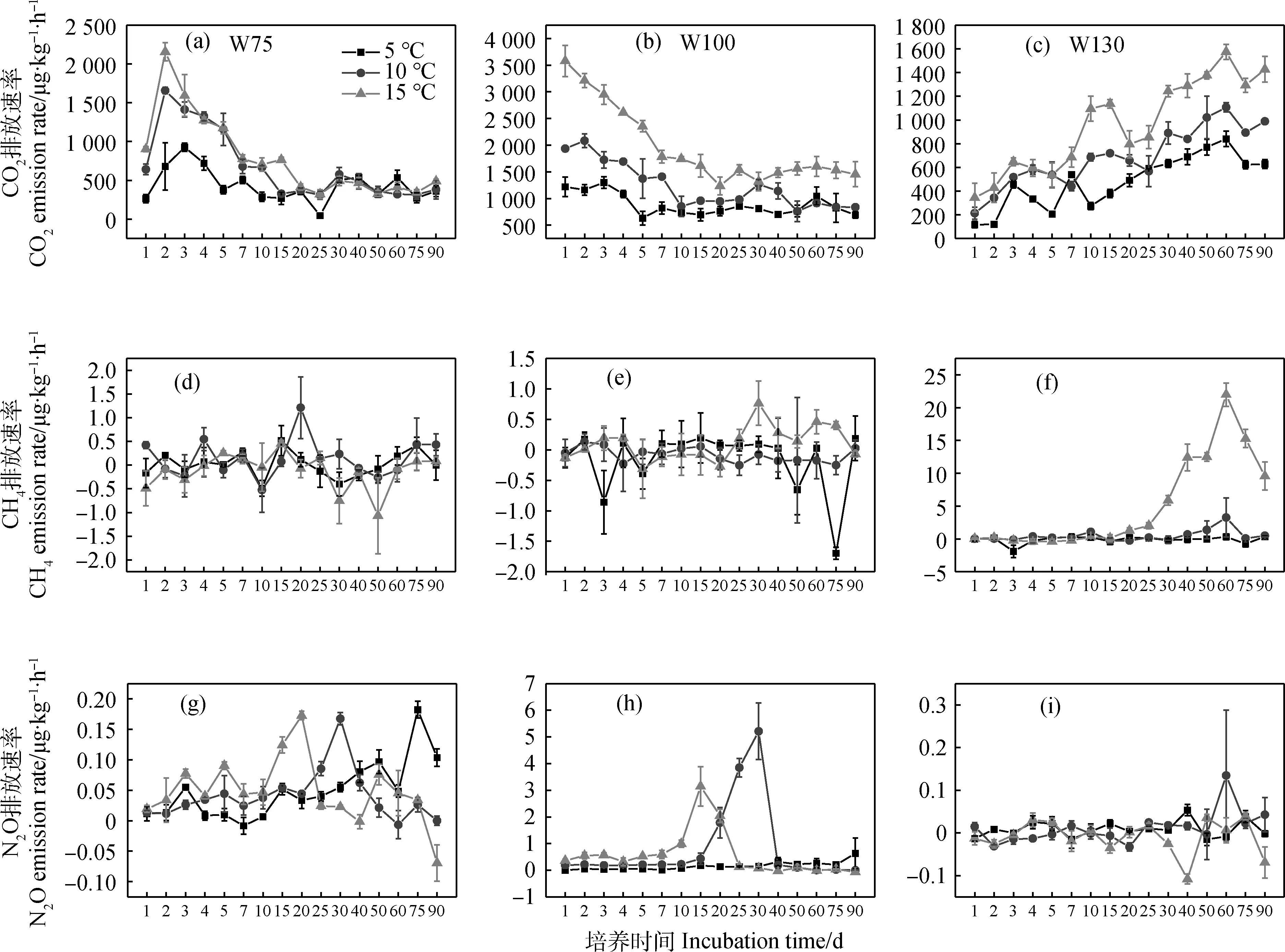
图1 高寒沼泽草甸土壤温室气体排放速率的时间变化Fig.1 Temporal variation of greenhouse gas emission rates from soil of alpine swamp meadow
2.2 温室气体累积排放量变化
各处理CO2累积排放量在整个培养期间变化趋势一致,表现为稳步增加,各水分处理下,CO2累积排放量均随温度升高呈增加趋势(图2a~2c)。各处理CH4累积排放量在培养前期变化趋势一致,前15天趋近于0,培养后期出现较大分异,W75和W100处理T5和T10与T15累积排放量在培养后期表现出相反趋势(图2d~2e),W130处理,T15累积排放量在30—90天期间显著高于T10和T5(图2f)。W75处理,各温度梯度下N2O累积排放量在前10天趋近于0,随后T5和T10逐步增加,而T15稳步增加到75天后开始降低(图2g);W100处理各温度梯度下N2O累积排放量在前7天变化不大,随后T10和T15快速上升,25天左右趋于稳定,T5则缓慢上升(图2h);W130处理,前30天各温度梯度下累计排放量趋近于0,30天后T5和T10逐渐增加,而T5呈波动式减少(图2i)。
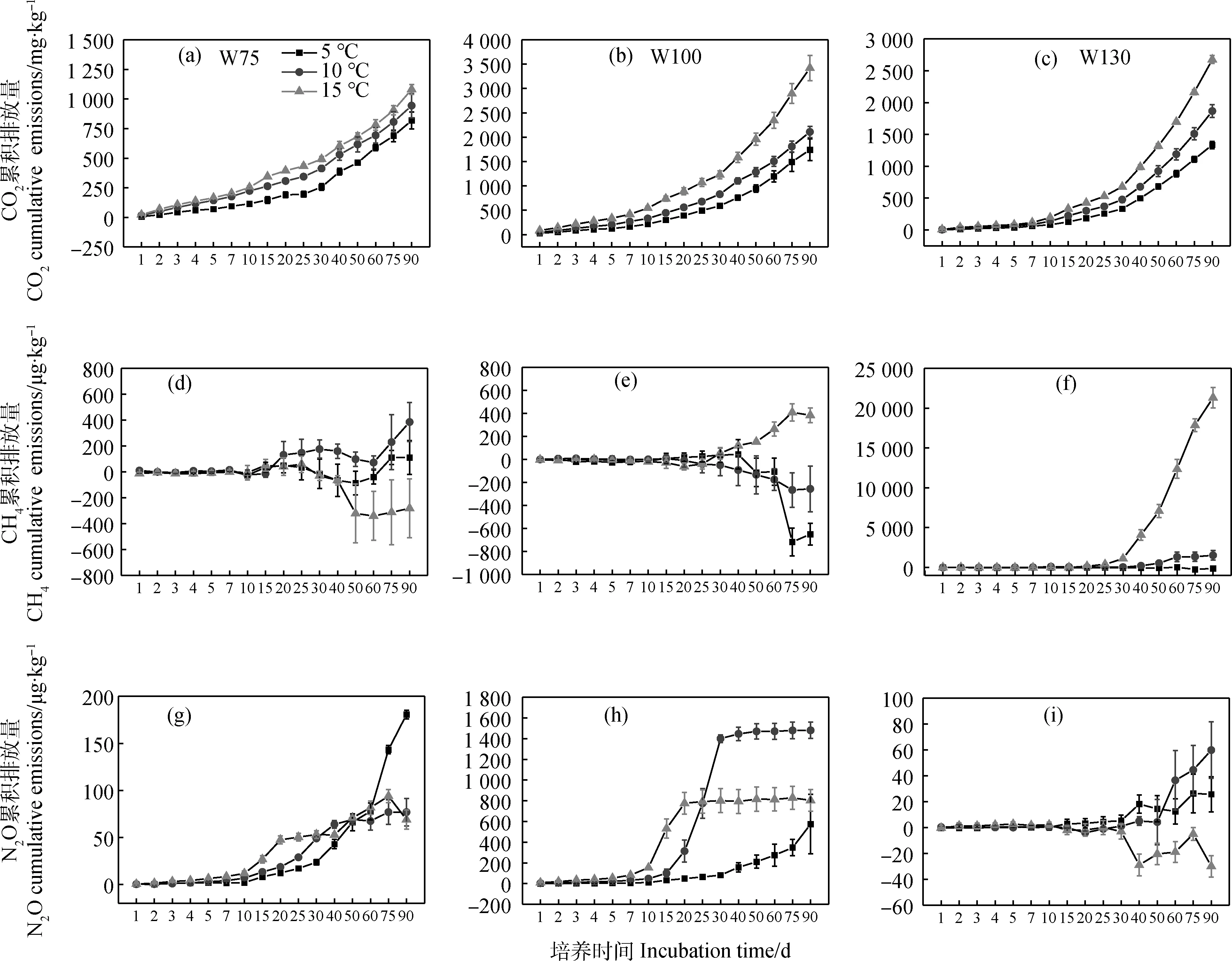
图2 高寒沼泽草甸土壤温室气体累积排放量Fig.2 Cumulative greenhouse gas emissions from soil of alpine swamp meadow
不同水分处理下CO2累积排放量均表现为T15>T10>T5,不同温度梯度下表现为W100>W130>W75(图3a),即随温度升高而增加,随水分增加先增加后减少;T15处理不同水分处理间差异显著(P<0.05),W130处理各温度处理差异显著(P<0.05),W100处理T15与T10和T5差异显著(P<0.05),W75处理各温度间差异不显著。W100和W130处理CH4累积排放量随温度增加而增加,W75处理随温度增加先增加后减少(图3b);W100和W130处理T15显著高于T10和T5(P<0.05),W75处理T10与T15间差异显著(P<0.05)。W75处理N2O累积排放量随着温度增加而降低,W100和W130处理随着温度增加先增加后降低(图3c);各温度梯度下,N2O累计排放量随着水分的增加呈先增加后减少的趋势;T10和T15处理N2O累积排放量均表现为W100处理显著高于W75和W130处理(P<0.05)。温度与水分变化对GWP影响与CO2累积排放量影响类似,表现为随温度增加而显著增加,随水分增加先增加后降低的趋势(图3d),T10处理各水分处理间差异显著(P<0.05)。
图3 90天培养期间温室气体累积排放量(平均值±标准误)Fig.3 Cumulative greenhouse gas emissions during the 90 day incubation (mean ± standard error)注:不同小字母表示同一水分下不同温度处理间差异显著(P<0.05);不同大写字母表示同一温度处理下不同水分含量间差异显著(P<0.05)Note:Different lowercase letters for the same moisture indicate a significant difference between different temperature treatments under the same moisture at the 0.05 level;different capital letters a significant difference between different moisture contents in the same temperature treatment at the 0.05 level
2.3 土壤理化性质特征
由表1可知,培养结束后,W75处理pH随温度增加而增加,W130随温度增加而降低,W100则随温度增加先增后减,水分变化对pH影响显著(P<0.05)。W75和W130处理EC随温度增加而增加,W100处理T5较T10和15差异显著(P<0.05),T10处理各水分间差异显著(P<0.05)。整体上,温度和水分对有机质含量影响较小,W75处理T5与T15较T10差异显著(P<0.05)。温度与水分变化均显著影响微生物量碳含量(P<0.05),各水分处理下均随温度增加而减少。W75处理T15温度下微生物量氮含量最高,水分显著影响微生物量氮含量(P<0.05),但温度以及温度与水分的交互作用对微生物量氮含量影响均不显著。温度和水分及其交互作用对蔗糖酶影响均不显著,T5处理过氧化氢酶随水分增加而增加,W75与W100和W130差异显著(P<0.05),T5和T15处理脲酶随水分增加而降低,且T5和T15处理各水分间差异显著(P<0.05),温度、水分及其交互作用显著影响脲酶活性(P<0.05)。
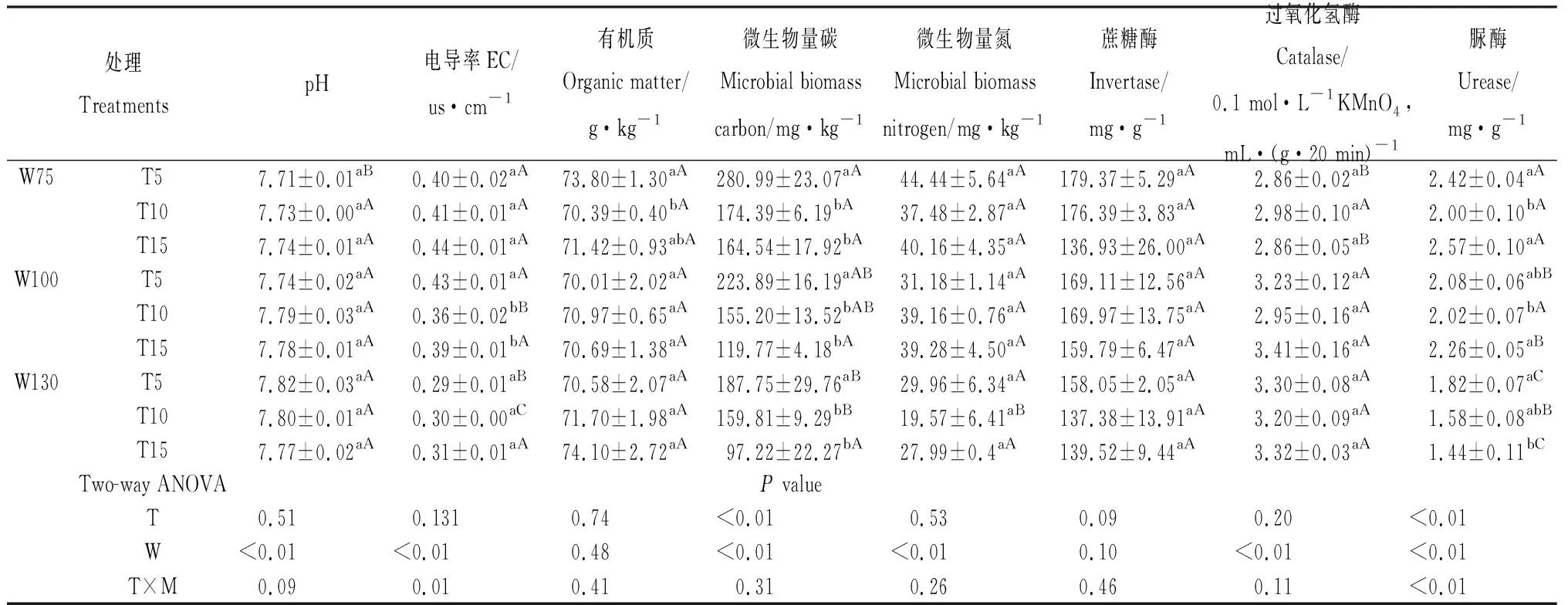
表1 不同温度和水分含量对土壤性质的影响Table 1 Effects of different temperature and moisture content on soil properties
2.4 温室气体通量影响因子
方差分析结果表明,温度、水分及二者交互作用与CO2,CH4,N2O累积排放量及GWP均有显著影响(P<0.01,表2)。模型拟合结果表明,温度、水分及二者交互作用与CO2累积排放量具有显著指数关系(P<0.01),N2O累积排放量与温度具有显著指数关系(P<0.01);温度和水分交互作用对CO2和CH4均具有显著影响(P<0.05),而对N2O影响不显著。CO2排放速率的温度敏感系数Q10值分别为1.7(W75处理),2.32(W100处理),1.98(W130处理)(表3)。
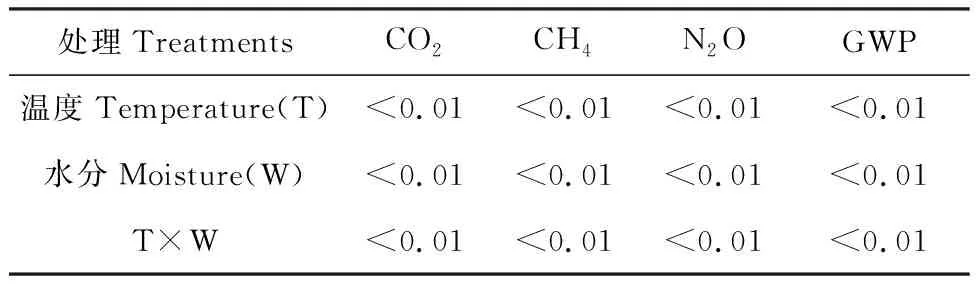
表2 温度、水分及二者交互作用与温室气体累积排放量相关性分析Table 2 Correlation analysis of temperature,moisture and their interaction with cumulative greenhouse gas emissions
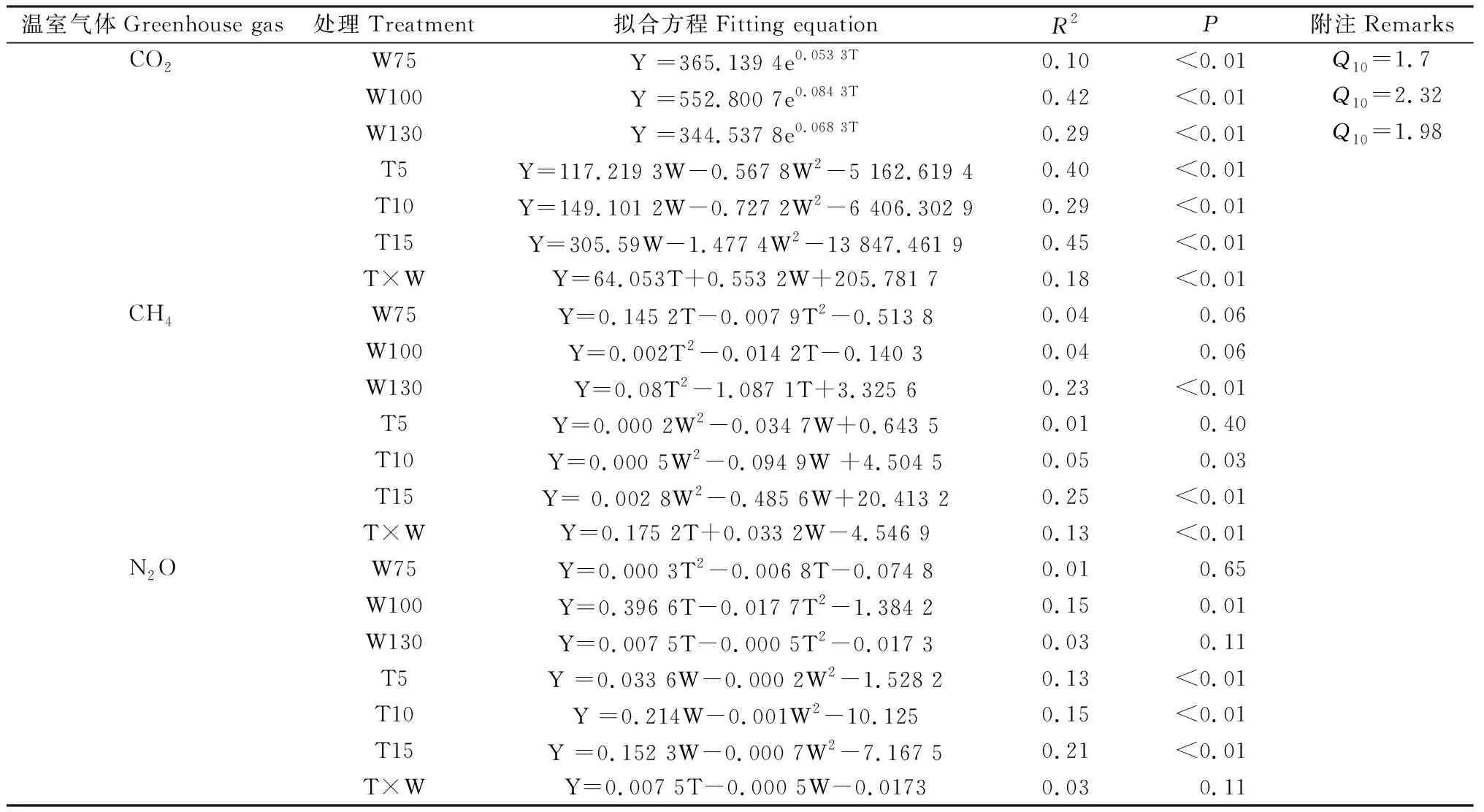
表3 CO2,CH4和N2O与温度、水分及二者交互作用拟合关系分析Table 3 Fitting relationship analysis of CO2,CH4 and N2O with temperature,moisture and their interactions
Pearson相关性分析表明,土壤CO2累积排放量与MBC呈显著(P<0.05)负相关,与pH值呈显著(P<0.05)正相关,与过氧化氢酶呈极显著(P<0.01)正相关;CH4累积排放量与EC呈显著(P<0.05)负相关,与脲酶呈极显著(P<0.01)负相关;而N2O与土壤因子均没有达到显著水平(图4)。方差分解分析发现,水热、土壤性质及其相互作用分别解释了GWP变异的31.95%,14.49%和14.72%(图5),表明GWP不仅受土壤温度和水分相互作用的调节,还受土壤化学性质的综合影响。
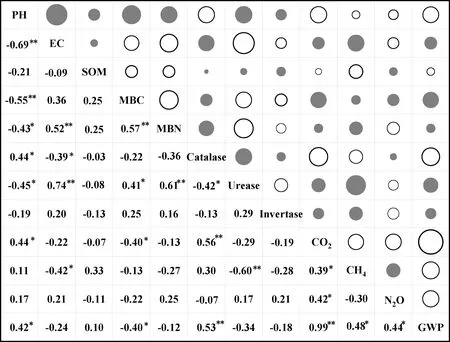
图4 培养90天后土壤性质与温室气体累积排放量的关系Fig.4 Relationship between soil properties and greenhouse gas emissions for 90 days of incubation注:图中白色圆圈表示正相关,灰色圆圈表示负相关,圆圈大小表示相关性R2的大小,*显著相关(P<0.05); **极显著相关(P<0.01)Note: The white circle indicates positive correlation, the gray circle indicates negative correlation, and the size of the circle indicates the size of the correlation R2, *mean a significant correlation at the 0.05 level, **means a very significant correlation at the 0.01 level
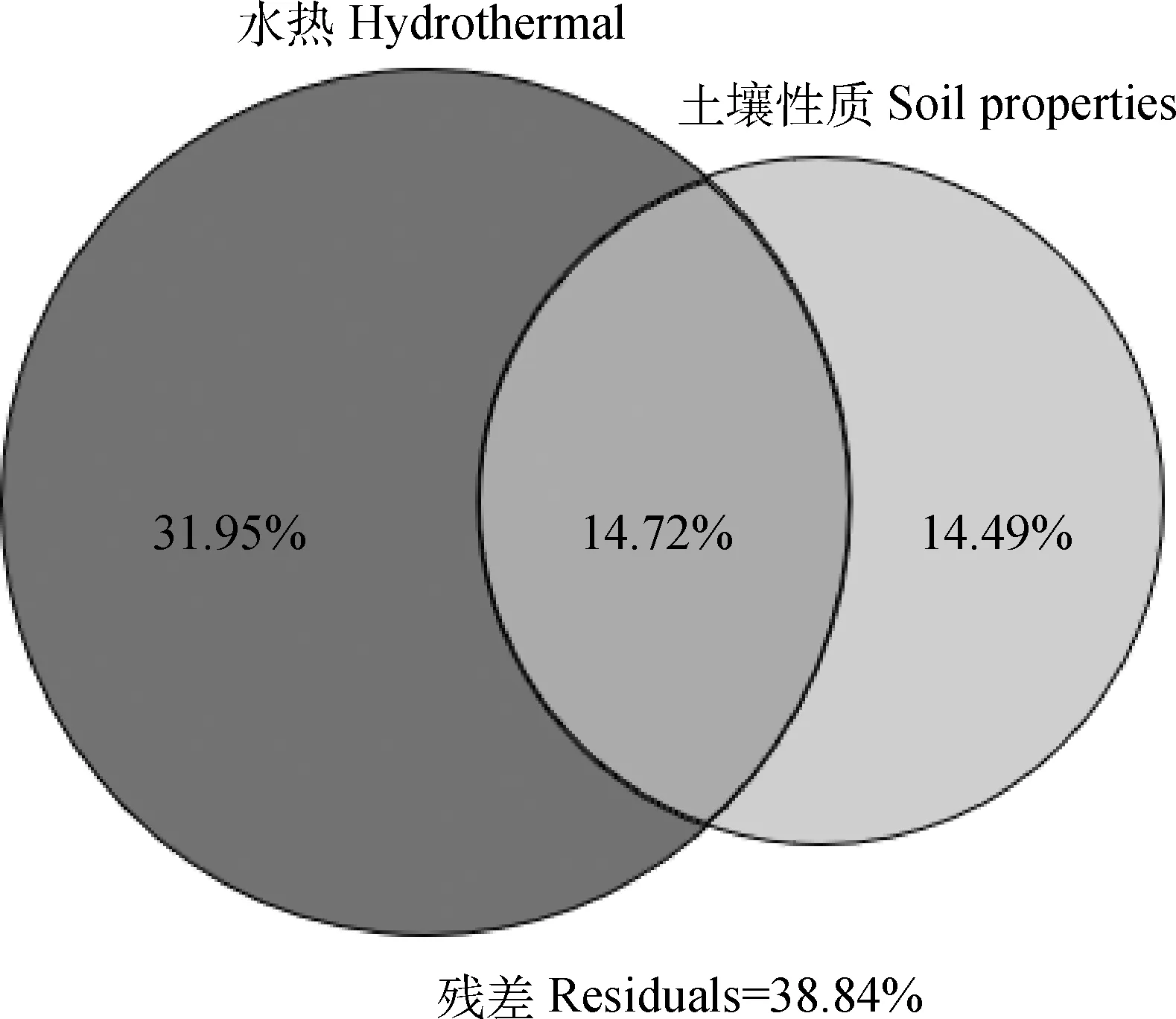
图5 土壤水热因子和土壤性质对全球增温潜势(GWP)影响的方差分解分析Fig.5 The variance decomposition analysis of global warming potential (GWP) influenced by hydrothermal factors and soil properties注:图中重叠代表共同影响,残差代表无法解释的变化Note:The overlap represents the common influence from hydrothermal and soil factors and the residual represents the unexplained variation
3 讨论
3.1 温度变化对土壤温室气体排放的影响
随着温度的增加,高寒沼泽草甸土壤CO2通量显著增加,这与Chen等[28,43]原位试验研究结果一致,其原因是增温增加了土壤微生物的活性和数量,使CO2排放速率增加33.4%[30]。增温显著降低微生物生物量碳含量(表1),且CO2累积排放量与微生物量碳含量负相关(图4),表明土壤温度升高促进土壤活性碳的分解,从而增加了CO2排放。100%WHC处理各温度梯度下CO2累积排放量均最高(图3a),此时土壤不受淹水和干燥胁迫,以及土壤氧浓度的限制,表明增温促进高寒沼泽草甸CO2的排放,但其受到土壤水分含量和含氧量的共同调控。也有研究表明,温度对CO2通量的直接影响很小[44]或没有影响[25],其原因可能因为温度对土壤有机碳分解还受制于水分状况等其它因素。
CH4累积排放量受温度显著影响(表2)。已有研究发现,CH4累积排放与土壤温度呈正相关关系,但不显著[45],增温处理可使CH4平均排放量增加28%[46];较高温度可增加产甲烷菌的多样性和丰度,从而使CH4排放提升[47-48]。也有研究发现,增温使高寒草甸的CH4吸收增加16.2%[49]。本研究W130处理下,T15处理在培养后期CH4排放速率和累积排放量较T5和T10处理显著增加(图1f和图2f),其原因是淹水条件下升高温度使土壤可溶性碳(DOC)和微生物胞外酶活性增加,有利于产甲烷菌和甲烷氧化菌的活动[50],此外,淹水条件会降低土壤通透性和土壤O2可利用性,进而影响CH4氧化和扩散[24]。
本研究发现,温度对N2O排放的影响,在不同含水量下存在很大差异(图2g~2i),如在W130处理下,T15处理表现为N2O的吸收,T5和T10处理表现为N2O的排放(图2i),与Chen等[28]研究结果类似。温度是影响土壤N2O排放的重要因素[3],已有研究发现,温度升高加速土壤氮矿化增加无机氮可利用性,进而促进N2O排放[51],也有研究发现,增温降低多年冻土区高寒草甸N2O排放或没有影响[52-53],温度低于15℃时N2O排放通量随温度升高呈递减趋势[54]。因为氮的可利用性和氧气可利用性限制,高寒沼泽草甸土壤既是N2O汇,又是N2O源[29]。
3.2 水分变化对土壤温室气体排放的影响
W75和W100条件下,CO2累计排放量随水分增加而增加,W100增加至W130排放降低(图1a~1c),与Gao等[55]的研究结果基本一致。土壤水分通过改善土壤通气性和氧气浓度,增强土壤微生物活性,进而刺激SOC分解和CO2排放[56-57]。适宜的土壤水分有利于O2和可溶性基质的扩散,土壤水分过高或过低都会抑制土壤中可溶性有机碳的扩散,致使土壤中微生物因缺乏能量来源物质而活性降低[58]、数量减少[59],从而抑制CO2排放。土壤水分含量100% WHC,CO2累积排放趋向于最大值(图3a),可能是因为土壤水分含量饱和时,此时土壤微生物呼吸速率最大[60]。研究发现,与未淹水(土壤含水量为25%~40%)的沼泽土壤相比,淹水厌氧处理使土壤CO2排放率降低6%~44%[55],当水分过高时,土壤处于厌氧环境,抑制了微生物的活性,使其分解土壤有机质速率降低[61],从而导致CO2通量排放降低。
W75和W100处理CO2排放主要集中在培养前期,可能是添加水分后不稳定有机碳的异养消耗所致[62],也有研究发现,土壤水分减少使CO2排放速率降低47.4%[30]。本研究,水分变化对土壤pH值、微生物量碳和过氧化氢酶影响显著(表1),且土壤pH值和过氧化氢酶与CO2累计排放量呈显著正相关,微生物量碳与CO2排放呈显著负相关(图4),这表明土壤水分增加,提高了土壤pH值和过氧化氢酶活性,从而促进微生物繁殖与代谢过程,降低微生物量碳含量,导致更多碳损失。本研究土壤呼吸的温度敏感性Q10值随着土壤水分增加先增加后减小,其原因可能是淹水厌氧条件限制土壤呼吸底物有效性、土壤酶活性等。
产甲烷过程是有机物再矿化的最终步骤,由严格厌氧古菌参与完成[63]。一般而言,CH4排放与土壤水分呈显著正相关[45]。土壤水分增加使CH4吸收降低50.64%[49],土壤水分通过影响产甲烷菌的多样性和丰度[64],从而影响CH4的排放和氧化。本研究,随土壤水分增加,CH4累积排放先降低然后剧增,这是因为沼泽草甸土壤淹水条件,为产甲烷菌产生CH4提供了有利的厌氧条件[22,57],从而产生大量CH4,但在湿润条件下,厌氧层厚度会影响O2可利用性,阻碍CH4运输和扩散,导致CH4排放降低。此外,本研究,水分显著影响土壤EC(表1),且土壤EC与CH4排放呈显著负相关(图4),表明水分变化可以通过调节土壤EC值来改变CH4氧化菌种群的pMMO酶活性强度来影响CH4排放[65],杨文柱等[66]发现,EC越低,CH4吸收通量越低,可能是该研究地点本就是盐碱地,较低的EC,土壤甲烷氧化菌比活性高,CH4排放减少。
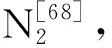
3.3 温度与水分交互作用对土壤温室气体排放的影响
本研究发现交互作用对三种温室气体均存在显著影响(表2)。一些研究表明,温度和土壤水分的交互作用对CO2排放速率没有显著影响[55],但当温度从5℃上升到35℃时,沼泽土CO2排放速率在非淹水处理下增加3.3~3.7倍,淹水处理下增加2.2~2.3倍[55]。也有研究表明,温度与水分相互作用加剧了净碳量的损失[44]。W75处理下,所有增温处理表现为CO2排放增加(图2a),表明高温和干燥条件促进CO2排放,与Shi等[30]研究结果相反,可能是因为该研究中土壤含水量范围为22%~35%,而本研究土壤含水量较高为75%,水分胁迫对CO2影响较小。W130条件下,CO2排放随温度升高而增加(图2c),其原因可能是淹水环境促进可溶性有机碳积累[72],增加土壤微生物活性和可利用碳底物含量,进而促进CO2排放[73]。各温度梯度下,随着水分增加CO2排放先增加后减少,其原因淹水条件会降低O2可利用性,使参与土壤有机质分解和转化过程的微生物酶活性降低而抑制土壤微生物呼吸[21]。方差分解分析发现,水热因素和土壤性质共同解释了GWP变异的61.16%(图5),表明GWP不仅受土壤温度和水分相互作用的调节,还受土壤性质影响。
水热交互作用显著影响CH4排放速率[55],研究发现,温度和水分共同作用使CH4平均排放量显著降低58%[46]。本研究,W130处理条件下,15℃培养的土壤比5℃和10℃培养的土壤具有相对较高的CH4排放速率(图1c),模型拟合结果显示,不同温度下,W130处理与CH4排放速率显著相关(P<0.01),其R2(0.23)最大(表3),15℃时土壤CH4累积排放量较高,表明温度从10℃升高到15℃可能对CH4产生过程影响较大。CH4通量在生长季节受土壤含水量的调节,而在非生长季与土壤温度密切相关[74]。也有研究发现,温度和水分对土壤CH4吸收并没有显著交互作用[49]。W130处理下,T5温度条件CH4排放速率为负值(图2f),表明高寒沼泽土壤可能是大气CH4汇,而温度的升高使高寒沼泽土壤由CH4吸收汇转变为CH4排放源[47]。W75条件下,T15处理表现为CH4吸收,而T5和T10处理表现为CH4排放(图3b),表明高温和干燥条件会减少CH4排放,与Yang等[46,75]的研究结果一致。当土壤淹水时土壤透气性和O2有效性降低可能会抑制CH4氧化,而在相对潮湿的土壤中,增温可以改变土壤充满水的孔隙空间并缓解厌氧条件,这有利于CH4扩散[76]。
土壤N2O的排放主要取决于微生物硝化和反硝化作用[77]。温度升高和水分增加会增加活动层厚度,使多年冻土N2O排放量增加,并导致强烈的非碳气候变化反馈[78]。N2O累积排放受温度与土壤水分交互作用的显著影响(表2),而交互结果模型结果不显著(表3),这可能是因为该模型对N2O解释度较低。W75处理下,增温处理整体表现为N2O排放减少(图2 g),表明高温和干燥条件共同抑制N2O排放。已有研究发现,土壤水分的减少抵消了气候变暖的正效应,导致土壤微生物种群规模和气体代谢的负响应[69]。此外,在高海拔地区,土壤水分对土壤氮库的影响比增温的影响要大得多[79]。
4 结论
高寒沼泽草甸土壤温室气体排放受温度、水分及其交互作用显著影响,75%~130% WHC下CO2累计排放量均随着温度增加而增加,130% WHC淹水条件下,CO2和CH4累计排放量显著增加,T10和T15处理N2O累积排放量均表现为100% WHC处理显著高于70%和130% WHC。各水分处理下,GWP均随温度增加而增加,其中T10处理100% WHC显著高于75%和130% WHC;GWP受水热互作和土壤理化性质综合影响,二者共同解释61.16%的GWP变异,表明多年冻土区沼泽草甸土壤温室气体排放受水热过程和土壤理化性质共同驱动。气候变暖背景下,全球增温潜势呈增加趋势,但其增幅强度受土壤水分调控。
