基于高密度遗传图谱的多环境玉米穗长QTL 定位
2023-11-23马晓杰赵延明王军燕周苗苗彭欣潘乃菲高惠敏刘宸铭苏成付
马晓杰,赵延明,王军燕,周苗苗,彭欣,潘乃菲,高惠敏,刘宸铭,苏成付
(青岛农业大学农学院,山东 青岛 266109)
玉米是重要的粮食和饲料作物,也是一种生物质能源材料[1-2],在人们生产生活中占据重要地位。 果穗是玉米的主要收获器官,穗长是影响玉米籽粒产量的重要性状之一。 Jenkins 等[3]早期研究表明,玉米自交系产量与穗长性状显著正相关;梁晓玲等基于17 个玉米杂交种[4]、李泉木等基于20 个杂交组合[5]的研究结果也证明穗长性状与产量间存在正相关关系。 因此,定位玉米穗长的QTL 具有重要意义,不仅有助于理解玉米穗长性状的遗传基础和分子调控机制,而且可为挖掘玉米产量潜力奠定基础。
前人已对玉米穗长性状开展了一些QTL 定位研究,但因穗长是复杂的数量性状,不同试验材料在不同环境下的研究结果不同[6]。 王帮太等[7]以87-1 和综3 为试验材料构建染色体单片段代换系(SSSL),并基于两种环境下的表型数据,定位到20 个穗长QTL 位点,解释表型贡献率12.10%~19.18%。 胡利宗等[8]基于NX110 与NX5314 组合的BC2F2回交群体,定位到4 个控制穗长的QTLs,解释表型变异1.32%~23.50%。 Li等[9]基于Dan232 和N04 杂交组合的F2、BC2F2和RIL 群体,在4 个环境下鉴定出14 个控制玉米穗长的QTLs,解释表型变异4.5%~8.7%。 Sabadin等[10]基于玉米自交系L-06-05F 与L-14-4B 组合获得包含400 个家系的F2∶3群体,定位到5 个控制玉米穗长的QTLs,解释表型变异3.1%~7.9%。汤继华等[11]以综3 与87-1 为材料配制杂交组合,利用F2群体,在两种环境中鉴定到8 个穗长QTLs,解释表型变异3.11%~11.86%。 Ross[12]利用玉米自交系SE-40 与LE-37 杂交获得的F2和F2∶3群体,分别定位到9 个和26 个与穗长相关的QTLs。 Veldboom 等[13]利用Mo17/H99 组合衍生的F2∶3群体定位到5 个穗长QTLs。 代国丽等[14]利用L26/095 组合的F2群体鉴定到3 个玉米穗长QTLs。 谢惠玲等[15]利用黄C/许178 组合获得的重组自交系,定位到8 个穗长QTLs。 霍冬敖[16]利用玉米自交系TY6 分别与Mo17 和W138组配F2分离群体以及F2∶3群体,鉴定到11 个玉米穗长QTLs,解释表型变异0.9%~15.6%。 李庭锋[17]基于吉846/掖3189 组合的重组自交系群体,定位到19 个玉米穗长QTLs,解释表型贡献率4.09%~14.49%。 王辉等[18]以郑58 和HD568 为亲本构建RIL 群体,在不同种植密度下进行QTL定位研究,检测到8 个穗长QTLs,分别位于1、2、3、4、9 号染色体,可解释表型变异4.37% ~10.50%。Zhou 等[19]以郑58/昌7-2 组合的F2∶3群体、D276/D72/A188/Jiao51 进行四元杂交获得的四交群体为材料,结合SSR 分子标记,在两环境下共检测到14 个穗长QTLs。 李卫华等[20]以昌7-2的单片段代换系为材料,共检测到22 个穗部性状QTLs,其中控制穗长性状的QTLs 有9 个,单个QTL 可解释8.6%~14.25%的表型变异;且发现了前人研究中同样被检测到的3 个玉米穗长QTLs 和1 个穗粗QTL 稳定表达位点。 Huo 等[21]以Mo17×TY6、W138×TY6 构建两套F2∶3家系群体,共检测到11 个穗长QTLs,可解释0.9% ~15.6%的表型变异;在1 号染色体发现一个一因多效位点qEL1.10,同时调控穗长及行粒数性状。Zhao 等以廊黄×TS141 和昌7-2×TS141 两组亲本构建的两组F2∶3家系为材料,检测到9 个穗长QTLs,单个QTL 可解释4.0%~17.2%的表型变异[22];应用同样的试验材料,在不同环境下,利用复合区间作图法(CIM)及基于混合线性模型的复合区间作图法(MCIM)相结合,定位到2 个调控穗长的MQTLs[23]。 Shi 等[24]构建了240 个家系的DH 群体,结合高密度遗传图谱,定位到5 个与穗长相关的QTLs,并在1 号染色体上定位到成簇QTLs,穗长QTL(qEL1)、穗粗QTL(qED1)和穗轴粗QTL(qCD1)被重叠定位,3 个QTLs 分别解释15.7%、28.3%和22.6%的表型变异。
在穗长基因克隆上,张人予[25]克隆了玉米穗长基因EL3,通过CRISPR/Cas9 技术进行基因敲除和超表达、结合转录组技术验证了该基因。 Jia等[26]克隆了一个调控行粒数QTLqKNR6,研究结果表明该QTL 主效基因同时影响玉米穗长与行粒数性状。 Luo 等[27]运用全基因组关联分析(GWAS)技术,定位了一个未知蛋白编码基因YIGE1,通过基因敲除及过表达分析,验证了YIGE1基因对玉米花序分生组织(IM)、行粒数(KNR)和穗长(EL)具有正调控作用;进一步分析发现该基因可能通过参与糖和生长素信号途径,进而调控玉米IM 的发育,影响行粒数和穗长,最终增加玉米的产量。
通过多环境QTL 定位研究可以发现控制穗长的稳定QTL 位点,既为进一步精细定位、克隆相关主效候选基因并深入研究其调控机制奠定基础,也为玉米基因组学研究提供重要的试验材料和数据支持,有助于进一步深入解析玉米穗长遗传调控机制。 虽然目前已有大量研究者利用不同作图群体检测了数百个穗长QTLs,但多为初定位,未经多环境验证其稳定性,很难进一步克隆利用。 本研究以穗长性状差异显著的玉米自交系为亲本,通过配制杂交组合,获得F2代分离群体。基于F2代群体构建高密度遗传图谱,结合多环境穗长表型数据,采用WinQTLCart2.5 中的复合区间作图法对目标性状QTL 进行定位,旨在对玉米穗长性状进行遗传剖析,研究穗长的遗传机制,提供新的玉米穗长QTL 位点,为高产玉米新品种选育提供理论参考,加速玉米育种进程。
1 材料与方法
1.1 分离群体构建及表型调查
2013 年夏以穗长差异显著的玉米自交系SG5 和SG7 为试验材料配制杂交组合,获得F1代杂交种子,自交F1得到F2代种子。 2014 年冬在海南省三亚市的盘县玉米育种试验站种植由199株单株组成的F2分离群体,并自交,得到F2∶3家系。 2018 年和2019 年将F2∶3家系按穗行的方式,种植于盘县玉米育种试验站,采用完全随机试验设计,单行区,每个家系一行,每行15 株,行距和株距分别为0.50 m 和0.35 m,田间管理及水肥施用与当地大田玉米相同。 待果穗成熟收获晒干后,用直尺测量穗基部到穗顶端的长度,即穗长。F2∶3家系穗长通过测量株行中间正常8 个果穗的平均值获得。 表型数据录入Microsoft Excel 表中保存,频数分布图及方差分析由SPSS 20.0 完成。
1.2 遗传连锁图谱构建及QTL 定位
在玉米抽雄吐丝前,对两个亲本以及F2采取单株取样。 剪取大小合适的叶片分别装入带有标记的离心管中,置于液氮罐后转至冰箱-80 ℃保存待用。 DNA 提取、基因组测序、SNP 分型及高密度遗传连锁图谱构建的具体方法见前期研究[28]。 应用QTL Cartographer 2.5 的复合区间作图法(CIM)的向前回归模型进行穗长表型数据的全基因组扫描,步长1 cM,为避免遗传信息遗漏,LOD 值设置为2.5;QTL 加性或显性效应值为正表明QTL 的增加效应来自母本SG5,QTL 加性或显性效应值为负则表明QTL 的增加效应来自父本SG7。 应用MapChart 2.32 软件进行QTL 在染色体上的位置描述。 将多环境F2和F2∶3群体QTL定位结果进行比较,多环境重复表达的QTL 位点被认定为稳定的QTL 位点。
2 结果与分析
2.1 穗长性状的统计分析
将2018、2019 年种植的F2∶3分离群体分别记为F2∶3-2018 和F2∶3-2019。 对F2、F2∶3-2018 和F2∶3-2019 三个环境穗长表型数据进行统计分析,结果(表1 和图1)显示,三个环境下,亲本穗长表现稳定,SG5 穗长为14.38 cm,SG7 穗长为12.72 cm;F2家系间穗长变异范围为12.8 ~21.0 cm, 平均15.42 cm;F2∶3-2018 家系间穗长变异范围为12.1~20.2 cm, 平均15.37 cm; F2∶3-2019 家系间穗长变异范围为10.8 ~21.7 cm, 平均15.40 cm。可见,群体呈超亲混合分布形式,家系间差异明显,存在遗传变异,该分离数据可以进行QTL 定位分析。

图1 F2、F2∶3-2018、F2∶3-2019 群体穗长性状频次分布
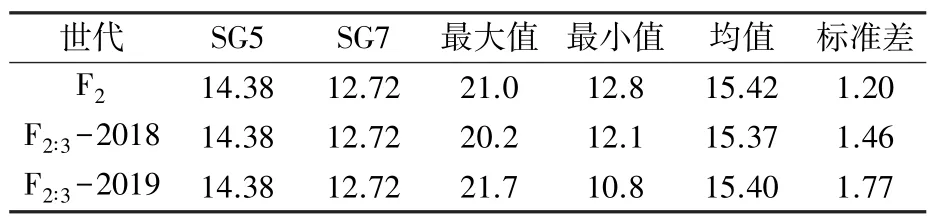
表1 F2、F2∶3-2018 和F2∶3-2019 分离群体的玉米穗长性状的统计分析 cm
2.2 QTL 定位结果
基于前期研究中构建的高密度遗传连锁图谱[28],利用CIM 法,对多环境穗长表型数据进行QTL 定位。 从检测结果(表2)看,在F2群体中共检测到4 个QTL 位点qEL-1、qEL-2、qEL-3和qEL-4,分别位于第1、2、5、6 染色体上,其中,位于第1 染色体上的qEL-1具有最大的表型贡献率,为24.0%,LOD 值为15.3;qEL-2具有最小的表型贡献率,为3.0%,LOD 值7.2。 在F2∶3-2018中共检测到4 个QTL 位点(qEL-1、qEL-2、qEL-3和qEL-5),分别位于第1、2、3、5 染色体上,其中,位于第1 染色体上的qEL-1具有最大的表型贡献率,为23.5%,LOD 值为16.6;qEL-2位于第2染色体,表型贡献率最小,为4.2%。 在F2∶3-2019中共检测到3 个QTL 位点(qEL-1、qEL-2、qEL-5),分别位于第1、2、3 染色体上,其中,位于第1染色体上的qEL-1具有最大的表型贡献率,为25.6%,LOD 值为15.3;qEL-2位于第2 染色体,其表型贡献率最小,为3.9%。

表2 F2、F2∶3-2018 和F2∶3-2019 分离群体的玉米穗长性状QTLs 定位结果
经遗传位置比对(图2),在F2、F2∶3-2018 和F2∶3-2019 三个环境下共定位到5 个穗长QTLs,其中,qEL-1和qEL-2在三个环境中重复被检测到,qEL-3在F2和F2∶3-2018 两个环境中被检测到,qEL-5在F2∶3-2018 和F2∶3-2019 两个环境中被检测到,而qEL-4只在F2环境中被检测到。

图2 基于F2、F2∶3-2018 和F2∶3-2019 分离群体的玉米穗长QTLs 位置
可见,qEL-1和qEL-2为三环境稳定表达QTLs。 从QTL 作用方式看,qEL-1表现为加性效应,其增效等位基因来自父本SG7;qEL-2表现为显性效应,其增效基因来自父本SG7。
3 讨论
遗传图谱的分子标记密度对QTL 准确定位及候选基因预测具有重大的理论价值。 分子标记密度低,分布不均匀,分子标记间的遗传距离大,则定位到的QTL 之间距离过大,对QTL 定位准确性影响较大。 随着测序技术的不断发展,全基因组测序逐渐被应用到QTL 定位中,SNP 分子标记借助于高通量基因分型技术,已在遗传分析中得到广泛应用。 同低密度遗传连锁图谱相比,高密度遗传连锁图谱具有众多优势:首先,可以更准确地定位QTL 位置,缩小候选区间,提高QTL 定位的精度,同时,高密度遗传连锁图谱还能够检测到更多的QTLs,提高检测的灵敏性;其次,高密度遗传连锁图谱可以提供更多的分子标记和候选基因信息,有助于筛选与目标性状相关的候选基因,进一步了解基因功能和分子机制;此外,高密度遗传连锁图谱可以加速分子育种进程,通过分子标记辅助育种,实现目标性状的精准选育。 可见,高密度遗传连锁图谱不仅能提高QTL 的检测效率,还能提供与QTL 紧密连锁的分子标记,用于分子标记辅助选择。 前期研究中基于F2分离群体构建的高密度遗传连锁图谱[28],为本研究QTL 定位提供了重组的分子标记数量,获得了与目标QTL紧密连锁的分子标记,为后期玉米穗长性状精细定位奠定了坚实基础。 应用构建的高密度遗传连锁图谱,我们分别在1、2、3、5、6 号染色体上检测到玉米穗长QTLs,其最大表型贡献率达到25.6%,位于第1 染色体上。 王帮太[7]、胡利宗[8]、Li[9]、霍冬敖[16]、李庭锋[17]、吕学高[29]、周广飞[30]等均在第1 染色体检测到玉米穗长QTLs;王帮太[7]、Li[9]、汤继华[11]、代国丽[14]、霍冬敖[16]等均在第2 染色体检测到玉米穗长QTLs。 可见,1、2 号染色体是挖掘玉米穗长性状的重要染色体,这与本研究在第1、2 染色体获得三环境稳定表达穗长QTLs 的结果一致。
多环境检测能够减少环境误差的影响,提高QTL 检测的准确性和可靠性,同时还能发现遗传效应较小的QTL,提高检测的灵敏性。 不同环境对同一性状的影响不同,在不同环境中检测QTL能够验证其效应是否一致和稳定,排除偶然或环境干扰的影响,有助于进一步研究QTL 的表达规律和遗传机制。 玉米大部分产量性状QTL 为复杂的数量性状,受环境影响较大,环境条件不同,产量性状QTL 表达也不同,因此在多环境中被重复检测到的QTL 可视为稳定QTL,对于作物遗传改良和分子育种具有重要的理论和实践价值。
4 结论
本研究通过多环境QTL 检测方法,以F2、F2∶3-2018 及F2∶3-2019 三环境穗长表型数据,结合高密度遗传连锁图谱,从SG5 和SG7 的F2∶3群体中检测到5 个控制玉米穗长QTLs,其中,qEL-1和qEL-2在三个环境中稳定表达,且qEL-1具有超过20%的稳定表型贡献率;qEL-3和qEL-5均在两个环境中被检测到。 这些QTLs 可视为多环境稳定表达位点,不仅可为玉米穗长性状主效位点的精细定位、图位克隆提供依据,同时可为玉米高产分子育种奠定理论基础。
