福建漳江口红树林湿地溶解无机碳横向输送的研究
2023-11-15王继纲高敏学贺万阳刘真松
张 娜,王继纲,高敏学,贺万阳,刘真松,张 劲,刘 茜
(1. 中国海洋大学 深海圈层与地球系统前沿科学中心和海洋化学理论与工程技术教育部重点实验室,山东 青岛 266100;2. 中国海洋大学 化学化工学院,山东 青岛 266100;3. 中国海洋大学 海洋高等研究院,山东 青岛 266100;4. 日本富山大学 理学部,日本 富山 9308555)
红树林湿地作为全球生产力最高的海洋生态系统之一,是有效的蓝色碳汇,在全球碳循环中发挥着重要作用。除了垂向上与大气气体交换和沉积物碳埋藏外,红树林湿地还通过潮汐作用与邻近河口和海洋中的溶解无机碳(Dissolved Inorganic Carbon, DIC)、溶解有机碳(Dissolved Organic Carbon,DOC)和颗粒有机碳进行水平方向上的交换(Alongi, 2020; 王法明等, 2021)。红树林的横向碳交换在全球碳循环中具有重要的科学意义,研究表明红树林是近海总碱度(Total Alkalinity,TA)、DIC、DOC和温室气体的净来源(Lu et al, 2023),且DIC是溶解碳输出的主要形式(Cabral et al, 2021)。这种横向交换主要通过孔隙水交换或海底地下水排放(Submarine Groundwater Discharge, SGD)来实现(Maher et al, 2013; Call et al, 2019b; Chen et al, 2021b)。SGD指从海底排放进入近岸海域的所有水流(Burnett et al, 2003),包括陆源SGD和咸化SGD(包括再循环海水和孔隙水)(Garcia-Orellana et al,2021)。尽管SGD或孔隙水交换的流量较小,但其输送的物质通量不容忽视。红树林湿地沉积物中存在广泛的动物洞穴,这些洞穴创造了大的孔隙空间,增加了沉积物的渗透能力(Guimond et al,2020),为孔隙水交换提供了绝佳的条件(Tait et al, 2016)。
镭存在4种天然同位素,包括短寿命的224Ra(半衰期3.66 d)、223Ra(半衰期11.4 d)以及长寿命的228Ra(半衰期5.7年)和226Ra(半衰期1600年)。放射性镭同位素是常用的示踪剂,可被用于评估河流-大型水库中水体运移及生物地球化学过程总体影响(Xu et al, 2018),也可用于示踪河口和近岸等短停留时间尺度下的孔隙水交换(Sadat-Noori et al, 2017),因此被广泛应用于河口、红树林湿地等环境中SGD/孔隙水的交换及其携带的物质通量(Sadat-Noori et al, 2015; Tait et al, 2017; Wadnerkar et al, 2021;Wang et al, 2021a; Wang et al, 2023),并探究了SGD输送的营养物质对沿海生态系统产生的影响(Wang et al, 2018; Xiao et al, 2019)。
SGD/孔隙水交换被认为是河口、红树林湿地系统中碳动力学的主要驱动力,评估其输送的碳通量对于准确评估红树林湿地的固碳能力和沿海碳收支具有重要意义。新修订的世界红树林蓝碳收支预算表明孔隙水通过细菌分解有机质横向输出DIC、DOC和TA,解决了“碳缺失”问题(Alongi,2022),中国九龙江河口的研究也揭示了SGD净输出DIC和TA的重要性(Wang et al, 2015)。在澳大利亚亨特河口进行的研究中发现,红树林通过孔隙水输出的DOC通量至少比通过当地河流输送到沿海海洋的通量高2个数量级,揭示了孔隙水交换的重要性(Sadat-Noori et al, 2019)。在中国广东漠阳江河口和江苏大丰麋鹿自然保护区的盐沼区进行的研究发现地下水/孔隙水是DIC输送的重要贡献者,DIC是溶解碳输出的主要形式(Chen et al, 2022; Wang et al, 2023)。其他研究表明,孔隙水/SGD输入的溶解碳通量远超河流输入通量,是红树林湿地碳收支的主要组成部分(Chen et al, 2018;王亚丽等, 2020)。然而,对于受人为活动影响的红树林湿地地下水输送碳的认识仍然有限(Wang et al, 2021b)。
已有研究报道了漳江口红树林溶解碳的地表水横向输送(Lu et al, 2023),结果表明红树林是漳江口TA和DIC的净来源,但关于漳江口红树林地下水输送碳通量的研究仍未见报道。为了探讨SGD对于碳横向输送的重要性,本文利用Ra质量平衡模型和潮动力模型,定量估算漳江口红树林孔隙水速率及其携带的碳通量,并对比利用FVCOM水动力模型计算的水流量和每小时碳浓度估算的该区域地表水横向输送的碳通量,以期为红树林区域蓝碳收支的准确评估提供数据支撑。
1 材料与方法
1.1 区域概况
漳江口红树林自然保护区(117°24′07″~117°30′00″E,23°53′45″~23°56′00″N)位于福建省东南部的云霄县漳江入海口,红树林中分布着许多潮沟,最终与漳江干流汇合,流入东山湾,再流入南海。保护区总面积为2360 hm2,是我国北回归线以北面积最大、种类最多、生长最好的红树林湿地保护区(周在明等, 2021)。漳江口红树林湿地属亚热带海洋性季风气候,年平均气温21.2 ℃,年平均降水量为1714.5 mm,主要分布在4—9月(Gao et al, 2021b)。该区域为半日潮,潮差0.43~4.67 m(平均2.32 m)(Wang et al, 2022)。在低潮时,上游的水产养殖废水和生活污水会通过一些闸门流入红树林潮沟。
1.2 样品采集、处理和测定
本研究选取红树林潮沟TS点(117°25′19.2″E,23°55′28.5″N)作为时间序列观测站,连续观测时间为2017年4月29日14:00至30日14:00,每个小时采集表层水(水深约0~0.5 m层)镭同位素、DIC、TA和pH样品。此外将4月29日17:48(AP1)和30日13:25(AP2)左右采集的水样利用预称重和预燃烧的GF/F膜(孔径0.7 μm,直径142 mm)过滤,收集颗粒态224Ra和228Th样品。使用多参数水质测定仪(WTW 3430, Germany)每小时原位测定水文参数(温度、盐度);使用Sea Sun Tech CTD 48 M探头连续记录时间序列观测站(TS)的水深;使用原位声学多普勒剖面流速仪(Acoustic Doppler Current Profiler, ADCP)连续测量流速;在潮沟位置附近选取5个孔隙水采样站,并用蠕动泵连接孔隙水采样器(pushpoint)从沉积物中采集孔隙水样品(PW1~PW5)。此外,分别于4月28日、29日和30日采集了外海水端元(SW)、漳江(ZJ)和闸门生活污水端元(ZM)的各参数样品,并测量了闸门水的水深和横截面积。采样站位见图1,端元点站位各参数数据见表1。
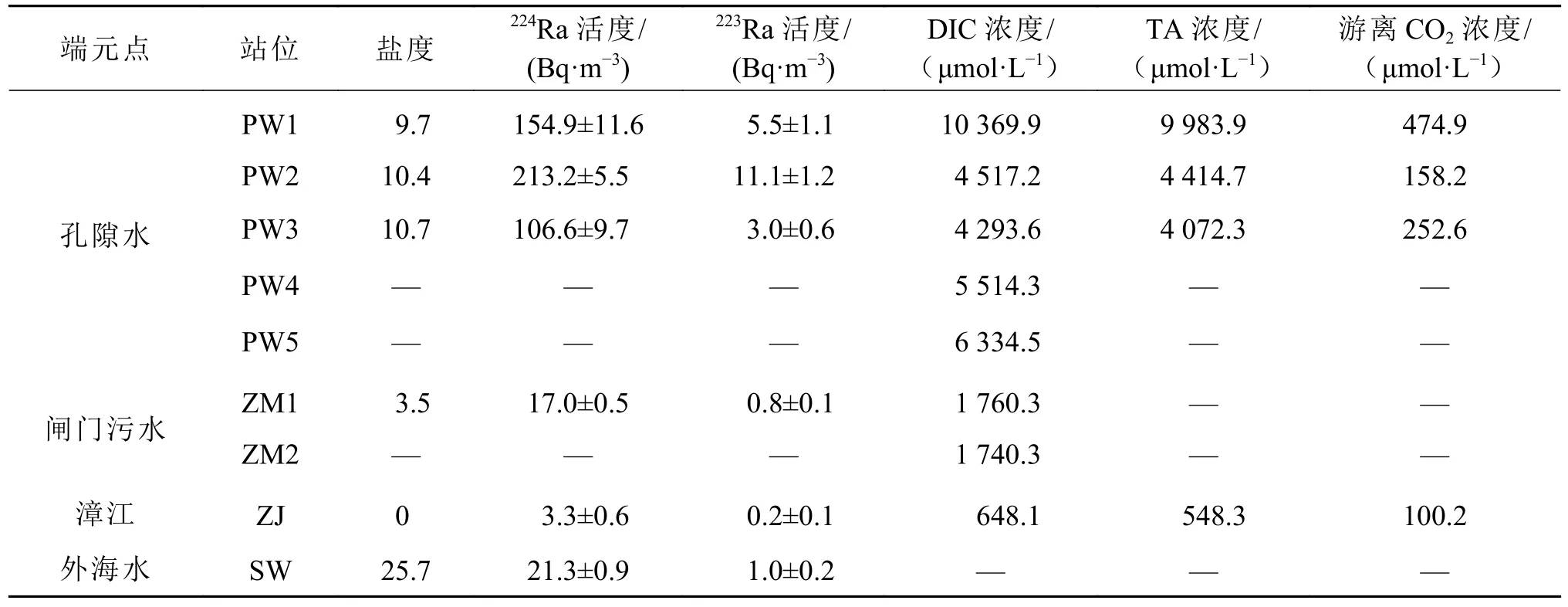
表1 红树林孔隙水、闸门、河水和海水端元的盐度、Ra活度和碳酸盐浓度Table 1 Salinity, Ra activity and carbonate system concentrations in porewater, sewage, river, and offshore seawater samples
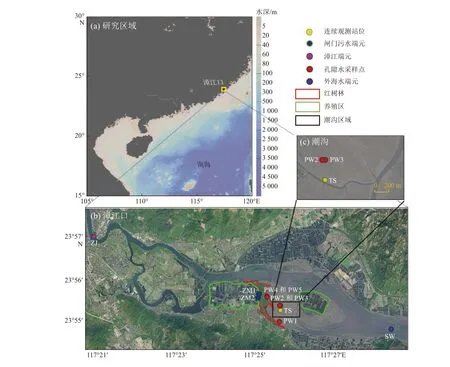
图1 研究区域采样站位分布Fig. 1 Map of sampling stations in the study area
采集的镭同位素样品(约30 L)分别经10 μm和1 μm的滤芯过滤后,以约1 L/min的流速通过预先称重的20 g左右的镀锰(MnO2)纤维,然后用蒸馏水清洗锰纤维以去除盐分,并控制水与纤维的质量比在0.4~1.1(Moore, 2008)。DIC、pH和TA样品利用Tygon管分别采集于125 mL硼硅酸盐玻璃瓶和125 mL高密度聚乙烯(HDPE)瓶中,样品充分溢出保证没有气泡。DIC、pH和TA样品收集后均加入0.1%饱和HgCl2阻止微生物活动,pH样品采样当天测定完毕,DIC和TA样品带回实验室,一周内进行测定。
锰纤维上富集的镭同位素活度用镭延迟符合计数器(RaDeCC,美国Scientific Computer Instrument)测定。基于3次测量确定223Ra和224Ra的活度,第一次测定在采样当天,第二次测定在采样7~10 d后,为了减少224Ra(半衰期3.66 d)对223Ra(半衰期11.4 d)产生的干扰。第三次测定在采样25~30 d,此时224Ra的活度由样品中的228Th产生,过剩的224Ra活度指第一次测量的总224Ra与第三次测量的228Th支持的224Ra之间的差值(Moore, 2008)。采集的颗粒态224Ra和228Th样品在采样后8 h内用改进的RaDeCC进行测量,详细内容见Cai等(2012)。DIC样品经酸化转化为二氧化碳,由氮气定量载入非色散红外二氧化碳检测器(LICOR-Model: AS-C3)测定;TA样品利用恒温槽使样品温度控制在25 ℃,利用Gran滴定法测定;DIC和TA的分析精度均为±2 μmol/L。TA和DIC测定均使用美国斯克里普斯海洋研究所(Scripps Institution of Oceanography)提供的海水参考物质进行校正。pH样品置于(25±0.2)℃的恒温浴中,待样品温度恒定后使用Orion 3 Star pH台式分析仪和Orion Ross组合pH电极(8102BN型)测定。pH测定的精密度和准确度为±0.005。根据测定的TA、DIC、盐度和温度,采用CO2SYS软件 (https://www.nodc.noaa.gov/ocads/oceans/CO2SYS/co2rprt.html.)计算pCO2和游离CO2浓度。
1.3 方 法
本文基于2种模型估算孔隙水交换通量,并结合孔隙水中的溶解碳浓度估算孔隙水输送的溶解碳通量,通过对比地表水横向输送的溶解碳通量揭示孔隙水输送的重要性。
1.3.1 Ra质量平衡模型
在假定稳态的系统中,近岸水体中镭通量的添加和损失相平衡,基于此,223Ra和224Ra在水体中的质量平衡方程(Liu et al, 2021)如下(式中所有源汇项的单位均为Bq/d):
式中:Friv和FPW分别表示河流和孔隙水输入的Ra通量;Fsed和Fdes分别表示沉积物扩散和悬浮颗粒物解吸的Ra通量;Fdec表示自身放射性衰变损失;Fmix表示与近海水域潮汐交换引起的混合损失;FZM表示闸门污水输入的Ra通量。
我们可以计算式(1)中除FPW之外的其他项:通过源汇项相等确定孔隙水输入的Ra通量;将FPW除以孔隙水端元中Ra活度(RaPW)和研究区域面积A,计算得到孔隙水速率(PWex,单位:m/d):
1.3.2 潮动力模型
另一种估算孔隙水交换的方法基于时间序列观测中的Ra同位素,计算式(Liu et al, 2021)如下:
式中:Rabkgd为背景值,为了规避潮汐外的其他影响因素,以潮周期内活度的最低值作为背景值;d为水深;τ为水体停留时间;Raobs为基于连续站观测的每一个时间点的Ra活度。
1.3.3 水体停留时间估算-镭同位素模型
该模型假设系统处于稳定状态,Ra的添加和损失相平衡,则河口中224Ra和223Ra的平衡可以表示为(Moore et al, 2006):
式中:F224Ra和F223Ra分别为进入研究系统的总的224Ra和223Ra通量;I224Ra和I223Ra分别为研究系统中224Ra和223Ra的储量;λ224和λ223分别为224Ra和223Ra衰变常数;数值分别取0.189 d−1和0.0608 d−1;τ代表系统的水体表观年龄,用式(4)除以式(5)可以得到:
式中:F(224Ra/223Ra)代表输入到研究系统内的224Ra和223Ra通量的活度比;I(224Ra/223Ra)代表系统内224Ra和223Ra的活度比。假设系统中Ra同位素主要来源于孔隙水排放的贡献,则F(224Ra/223Ra)可用孔隙水中224Ra和223Ra的活度比表示,I(224Ra/223Ra) 可用潮沟水体中224Ra和223Ra的比值表示。
1.3.4 水体停留时间估算-纳潮量模型
基于纳潮量模型,冲刷时间可以通过以下计算式估算(Moore et al, 2006):
式中:Tf为水体停留时间;V为水体体积;T为一个潮周期的时长(该区域为半日潮,取潮周期为0.5 d);P为涨潮时进入的潮水量,即纳潮量;R为漳江输入至红树林区域的净流量;b为落潮时返回外海的水量的占比,假设这个比例是海水端元的贡献比例,可以由每个被测样品的盐度除以海水盐度得到(Liu et al, 2018)。因为涨落潮时的水体面积未知,用以下计算式估算该区域的纳潮量(Moore et al, 2006):
式中:A为研究区域面积(802212 m2);z为一个潮周期内水深(H)的变化。
1.3.5 地表水横向输送
红树林潮沟地表水的横向输送通量通过每个小时的水流量乘以碳浓度,并在整个潮汐周期中积分来计算(Cabral et al, 2021)。每小时水流量利用Finite-Volume Community Ocean Model(FVCOM)水动力模型估算,正值代表从潮沟输出进入河口,模型覆盖漳江口和东山湾等海域(Wang et al, 2020)。
1.3.6 海-气CO2交换通量
海-气界面CO2交换通量(FCO2)取决于水体和大气中pCO2的差值以及水温、盐度和风速等环境因素。计算式(Sweeney, 2007)为:
式中:FCO2为海-气交换通量,单位为mmol/(m2∙h);Kh为特定温度和盐度下温室气体的溶解度系数,计算式(Weiss, 1974)为:
式中:T为热力学温度,单位为K;S为盐度;A和B为常数,单位为mol/(kg∙atm)(1atm=101325 Pa),A1=−60.2409,A2=93.4517,A3=23.3585,B1=0.023517,B2=−0.023656,B3=0.004736。
∆pCO2为潮沟水体和大气中pCO2的差值。假定大气中的CO2值为412 μatm(Reithmaier et al, 2020)。k2为气体传递速度(m/h),计算式如下:
式中:u为风速,使用010C风速传感器测量,本文所用数据引自Zhu等(2019),为同时期实时测定的风速;X为指数,当u<3.6 m/s时,X= 0.6667,当u>3.6 m/s时,X= 0.5;SC为CO2的施密特数(Wanninkhof et al, 1992),计算式如下:
式中t为温度,单位是℃。
2 结果与讨论
2.1 Ra活度及碳相关参数
在连续观测期间,潮沟表层水体的水深、盐度、Ra(224Ra、223Ra)活度、DIC浓度、TA浓度、pH、游离CO2浓度和pCO2的变化如图2所示,显示出明显的潮汐趋势。水深变化范围为0.6~3.3 m,pH变化范围为7.27~7.98,盐度变化范围为5.0~12.1,盐度与潮汐趋势相似,涨潮时随着海水的入侵盐度增加,落潮时盐度降低。潮沟水体224Ra活度为21.3~53.1 Bq/m3,平均值为(30.0±8.2)Bq/m3,223Ra活度为0.8~2.4 Bq/m3,平均值为(1.2±0.4)Bq/m3。Ra活度遵循类似的潮汐趋势,低潮时活度高,高潮时活度低(图2b和图2c)。

图2 2017年4月29日—30日连续观测期间各参数的潮汐变化趋势Fig. 2 The variations of various parameters with the tidal change during time series observation period from April 29 to 30, 2017
孔隙水盐度范围为9.7~10.7,平均为10.3。孔隙水中的224Ra和223Ra活度分别为106.6~213.2 Bq/m3和3.0~11.1 Bq/m3,平均活度分别为(158.2±43.6)和(6.5±3.4) Bq/m3,明显高于地表水中的Ra活度,表明孔隙水可能是地表水中镭同位素的主要来源。
地表水中DIC浓度、TA浓度、游离CO2浓度、pCO2分别为1156.8~1915.9 μmol/L、1150.8~1859.6 μmol/L、28.9~72.8 μmol/L、519.9~3122.1 μatm,平均值分别为(1434.3±220.3)、(1415.0±206.4)、(52.6±26.2) μmol/L、(1540.0±798.3) μatm。与镭同位素类似,DIC、TA、游离CO2浓度、pCO2在连续观测期间呈现明显的潮汐趋势,即低潮时浓度高、高潮时浓度低(图2e~图2h)。孔隙水中,DIC、TA、游离CO2浓度、pCO2分别为(6205.9±2206.5)μmol/L、(6156.9 ±2709.7)μmol/L、(295.2±132.7)μmol/L、(8854.6±4067.1) μatm,明显高于地表水中的浓度,这可能是由孔隙水中有机物在微生物活动的影响下降解所致(Alongi, 2022)。
但是第一个低潮时(21:00至23:00)(图2中阴影部分)最高的DIC浓度对应低的Ra活度。此时间段是污水闸门开放的时间,闸门附近采集的污水Ra活度(17.0 Bq/m3)远低于地下水和潮沟水体;但闸门污水的DIC浓度(1750.3 μmol/L)相对于潮沟水体较高(表1)。据此,推测第一个低潮时观测到的低Ra活度和高DIC浓度,可能是受污水排放的影响。另外排放的闸门污水也可能含有较高浓度的有机物,造成潮沟水体有机物耗氧呼吸产生高的DIC,这些需要今后的进一步研究。pH值在低潮时闸门污水排放的影响下没有遵循严格的潮汐趋势(图2d)。由于污水排放的影响,pCO2峰值未出现在最低潮,但在最高潮出现最小值519.9 μatm。
连续观测期间224Ra活度、DIC浓度与盐度之间的关系如图3所示。224Ra活度在落潮时存在显著的添加量(图3a空心圆采样点),涨潮时添加量不明显(图3a实心圆采样点)。落潮时224Ra的添加可能受悬浮颗粒物解吸和孔隙水排放的影响。DIC浓度与盐度的分布特征与224Ra活度和盐度的关系类似,在落潮时出现添加,可能受孔隙水或闸门污水排放的影响,涨潮时基本保守(图3b实心圆采样点),主要受河流和海水混合(图3b黑色线)影响,这与图2的规律相对应。

图3 2017年4月29日—30日连续观测期间224Ra活度和DIC浓度与盐度的关系Fig. 3 Plots of 224Ra activity and DIC concentration versus salinity during time series observations period from April 29 to 30, 2017
2.2 红树林潮沟Ra的源汇
河流输入的Ra通量包括溶解的Ra和颗粒物释放的Ra。溶解的Ra通量用河流径流量与河流端元点的Ra活度的乘积(表1)来估算。颗粒物释放的Ra通量由河流径流量乘以河流中颗粒物浓度(14 mg/L)、再乘以Ra的解吸分数求得。河流径流量选择的是漳江2020年4月的径流量,值为13.6 m3/s(Zhu et al, 2019),由此计算出2017年4月河流输入的224Ra 和223Ra通量Friv分别为4.1×106和1.8×105Bq/d。
近岸水体另一个潜在的Ra来源是底部沉积物扩散,特别是对于短寿命的Ra同位素,底部沉积物的贡献不容忽视。漳江河口沉积物主要为粉砂质,颗粒较细(Gao et al, 2021a),采用与该区域沉积物类型相同的红树林潮滩湿地最大Ra释放量(F224Ra=20 Bq/(m2·d),F223Ra=0.4 Bq/(m2·d))(谷河泉, 2015),然后乘以该区域底部沉积物面积(802212 m2),得到该区域沉积物扩散的224Ra和223Ra通量Fsed分别为1.6×107和3.3×105Bq/d。
在近岸水域,潮汐和波浪是沉积物重新进入水体的主要驱动因素。因此,悬浮颗粒物解吸也是镭同位素的来源。2017年4月,盐度为8.5(AP2)和11(AP1)时,颗粒态224Ra分别为0.25和0.02 Bq/g,228Th分别为0.35和0.32 Bq/g。假设224Ra和228Th在悬浮颗粒上处于平衡状态,颗粒物228Th和224Ra之间的差额代表河口中解吸的224Ra,然后除以228Th得到解吸的224Ra的分数(Wang et al, 2018)。计算得到的224Ra解吸平均分数为(43±18)%,与九龙江口东南河口(43%)报道的值相同(Wang et al,2015)。将平均颗粒态228Th((0.34±0.02) Bq/g)与224Ra解吸比例(43±18)%相乘,得到平均解吸224Ra为(0.15±0.06) Bq/g。223Ra的解吸量选用与该区域类型相同的红树林湿地沉积物的最大可交换223Ra量(7.5 ×10−3Bq/g)(谷河泉, 2015)。连续观测期间红树林潮沟的悬浮颗粒物浓度平均值为(186±116)mg/L。假设所有的悬浮颗粒物均来源于表层沉积物的再悬浮,利用悬浮颗粒物浓度乘以水量和镭解吸率估算悬浮颗粒物解吸产生的镭通量。将所得结果除以水体表观年龄,估算沉积物再悬浮解吸的224Ra 和223Ra通量Fdes分别为6.3×106和3.2×105Bq/d。
上游的水产养殖废水和生活污水在落潮时会经过一些闸门流入红树林潮沟,在采样期间(2017年4月29日21:00至23:00)观测到闸门污水的排放,因此,污水输入也是镭同位素的一个来源。通过测量的闸门水的水深(0.8 m)、闸门宽度(2.5 m)以及流速仪测定的水体流速(2.5 m/s)求得闸门污水的流量为3.4×104m3/d,然后将流量乘以相应闸门端元点Ra的活度(表1),得到闸门水输送的224Ra和223Ra通量FZM分别为5.7×105和2.5×104Bq/d。
镭的储量可由潮沟区域镭的活度与水量相乘来估算。由镭衰变常数估算的224Ra和223Ra放射性衰变通量分别为6.8×106和8.8×104Bq/d。由于潮汐活动,近岸水域与外海水域发生水体交换,为了估算镭损失,我们使用了Liu等 (2018) 的方法,它基于过剩镭(过剩的Ra活度指表层水体Ra的实测值减去外海水和河流贡献的Ra活度)的储量乘以1/τ的混合率,选择的外海端元点(用最靠近东山湾的站位SW在4月28日14:47采集的数据代替)盐度为25.7,224Ra和223Ra活度分别为21.3 和1.0 Bq/m3。在高盐度时镭的活度随着盐度的增加而降低,真正的海水端元盐度更高,镭更低。因此,计算的过剩的镭储量是低估,最终计算的SGD流量也是保守的低估。本研究应用224Ra/223Ra活度比和纳潮量模型估算水体停留时间。孔隙水中平均224Ra/223Ra活度比为(27.8±6.9),地表水中224Ra/223Ra活度比为(26.6±5.5),由式(6)计算出平均水体停留时间为(0.5±0.3)d。利用纳潮量模型(式7)估算的水体停留时间为(0.3±0.02) d。2种方法计算的水体停留时间在误差范围内基本一致。由此与外海水混合损失的224Ra和223Ra通量Fmix分别为4.7×107和1.7×106Bq/d。
最后,利用式(1)和已知的镭的源汇项计算孔隙水输入的224Ra和223Ra通量FPW,分别为2.7×107和9.4×105Bq/d。孔隙水交换是Ra的主要来源,占总来源项的50%~52%,其次是沉积物的贡献,沉积物贡献的224Ra占总来源项的30%。河流输入仅占总来源项的7%,对224Ra通量的贡献不大。研究区域大部分镭的损失是由于与外海水混合造成的,占总损失项的87%~95%(表2)。

表2 2017年4月红树林潮沟Ra的源汇收支Table 2 The sources and sinks of Ra in the mangrove tidal creek during April 2017
2.3 孔隙水速率的估算及不确定性分析
孔隙水速率通过孔隙水贡献的Ra通量除以地下水端元中的Ra活度得到。本研究中选择潮间带孔隙水的平均Ra活度作为地下水端元的Ra活度。基于224Ra和223Ra质量平衡模型估算的孔隙水交换率均为(0.2±0.1)m/d。潮动力模型中选择时间序列观测中测量值的最小活度作为近岸水的背景值,以计算保守的孔隙水交换率,基于式(3),通过224Ra估算的孔隙水速率为0.004~0.300 m/d,平均为(0.1±0.1)m/d,通过223Ra估算的孔隙水速率为0.05~0.40 m/d,平均为(0.2±0.1) m/d。应用2种方法(即质量平衡模型和潮动力模型)估算的红树林孔隙水速率范围为0.004~0.4 m/d,平均为(0.2±0.04)m/d,相比全球红树林而言,处于中间水平(表3)。可能是由于该研究区域沉积物主要由粉砂(69%~72%)和黏土(27%~31%)构成(邱勇等, 2019),且该区域红树林沉积物存在蟹洞,在沉积物中创造了大的孔隙空间,增加了沉积物的有效渗透率,增强了孔隙水的交换。
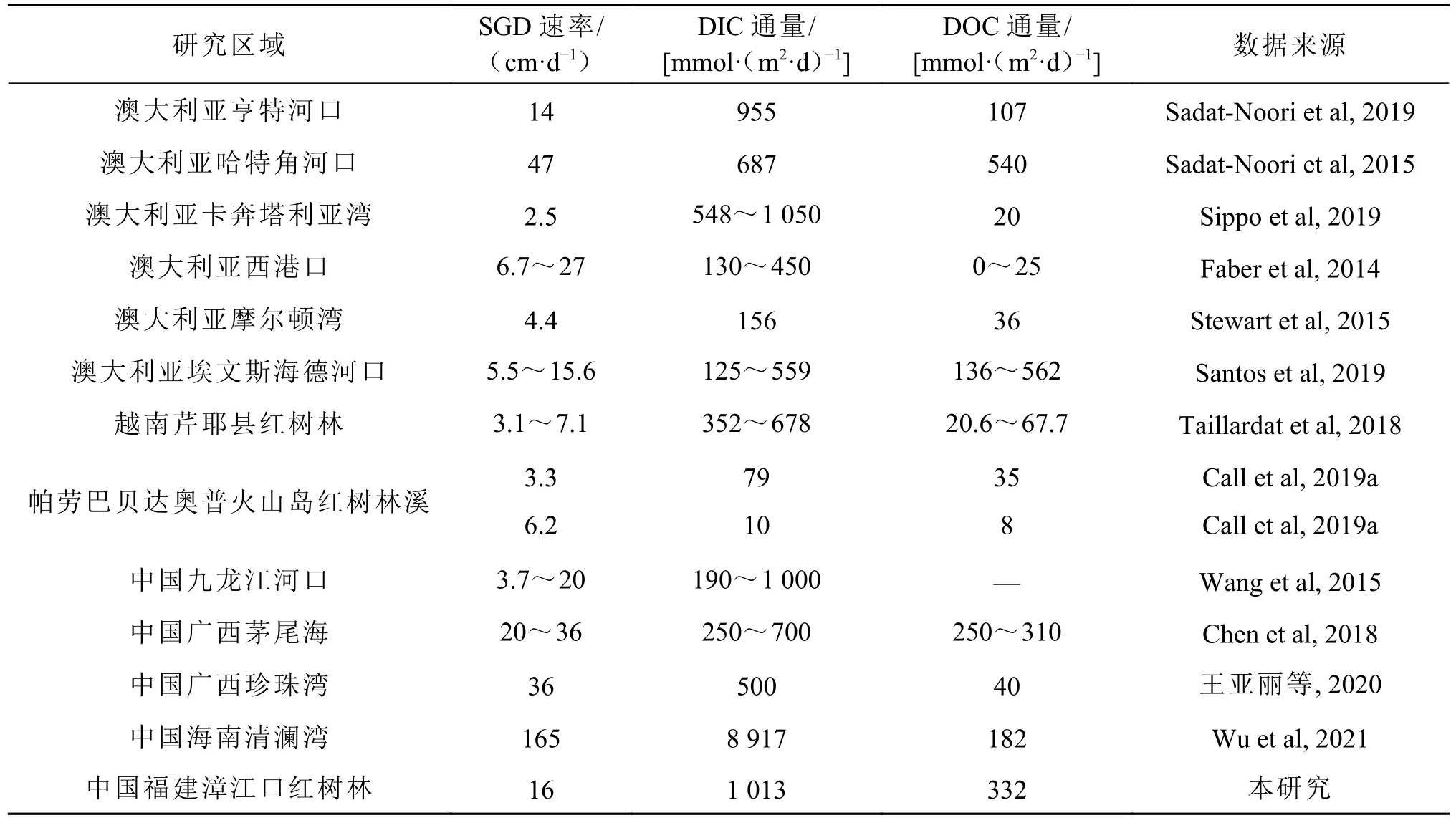
表3 河口和海湾等典型红树林生态系统SGD/孔隙水速率及其携带的DIC和DOC通量汇总Table 3 Summary of SGD rates and SGD/porewater derived DIC and DOC fluxes in typical mangrove ecosystems such as estuaries and bays
孔隙水交换率的不确定性是由式(1)中Ra的各个源汇项不确定性或误差引起的,包括Ra的测定误差和模型计算孔隙水交换率所用到的误差传递,具体计算原理和计算式见Taylor (1997)和Wang 等 (2015)。河流和污水输入通量的不确定性由Ra活度的测量误差导致,通过误差传递公式计算得到河流和污水输入通量的不确定性分别为16%~34%和3%~20%。由水体停留时间的不确定性导致的悬浮颗粒物解吸通量的不确定性为20%。由224Ra和223Ra活度测量误差导致的自身衰变损失的不确定性分别为27%和35%。孔隙水交换率的总的不确定性主要来自与外海水的混合损失和孔隙水端元中224Ra和223Ra的不确定度。对于224Ra来说,底部沉积物扩散的不确定性也是孔隙水交换率估算的不确定性的一个重要来源,其改变20%导致孔隙水速率变化38%。混合损失的不确定性源于Ra测量值和水体停留时间的不确定度,根据误差传递方程得到224Ra和223Ra混合损失的不确定性分别为31%和51%。孔隙水端元的不确定性源于孔隙水端元中Ra活度的空间变化。地下水端元224Ra和223Ra活度的相对误差分别为28%和64%,利用误差传递公式估算224Ra和223Ra示踪的孔隙水的不确定性分别为78%和66%。同时,本研究利用潮沟的一个时间序列观测点反推整个红树林通过孔隙水横向输送碳通量,也存在不确定性,在未来的研究中,需要在红树林地区的不同时间和空间尺度上进行一系列采样点,以更好地了解和评估红树林生态系统中孔隙水的重要性。
2.4 通过孔隙水和地表水输送的碳通量
孔隙水碳通量是指红树林通过孔隙水往潮沟横向输送的碳通量,地表水输送的碳通量是指在潮汐的作用下,潮沟水体横向交换输入至漳江河口的碳通量。本研究基于孔隙水速率与地下水端元和潮沟水体物质浓度的差值相乘,然后除以红树林的面积(802212 m2)来估算单位红树林面积孔隙水输送的物质通量。DIC、TA、DOC(孔隙水中的DOC浓度数据引自Wang等 (2019),地表水中的DOC浓度数据引自凌楠 (2018);DOC浓度数据均为同航次数据)和游离CO2浓度分别为4771.6、4751.6、1564、243.2 μmol/L,孔隙水输入的DIC通量为1013 mmol/(m2·d),TA通量为1008 mmol/(m2·d),DOC通量为332 mmol/(m2·d),游离CO2通量为52 mmol/(m2·d)。孔隙水流入潮沟的游离CO2通量仅占孔隙水输送DIC通量的约5%,孔隙水主要以碳酸氢根的形式向潮沟输送DIC。此外,潮沟地表水中DIC浓度与224Ra活度、TA浓度与224Ra活度呈现显著的正相关(P<0.05)(图4a、图4b),这意味着孔隙水交换可能是红树林潮沟水体DIC和TA的重要来源。在其他地方也观察到类似的现象。例如在崇明东滩盐沼近岸水域和澳大利亚亨特河口红树林湿地观察到DIC浓度与224Ra活度之间存在显著正相关(Sadat-Noori et al, 2019; Liu et al, 2021),这可能是由于湿地沉积物矿化产生的无机碳有一部分以DIC形式存在于孔隙水中(Lu et al, 2023),红树林孔隙水富含DIC,在潮汐泵的作用下携带大量的DIC排入到潮沟中。而河流DIC浓度低,对潮沟DIC影响不大。此外潮沟停留时间短,推测来不及发生生物地球过程显著影响潮沟的DIC浓度。因此孔隙水是潮沟DIC的重要来源。pCO2与224Ra活度之间未观察到明显的相关性(图4c),说明pCO2不是由孔隙水主控,可能受海气交换、温度和生物活动影响。潮沟水体海-气界面二氧化碳交换通量为1.5~55.4 mmol/(m2·d),平均为(17.3±14.9)mmol/(m2·d),正值代表潮沟水体是大气CO2的源,说明孔隙水输送至潮沟水体的游离CO2有34%(17.3/52)经过海-气CO2交换释放到大气中。
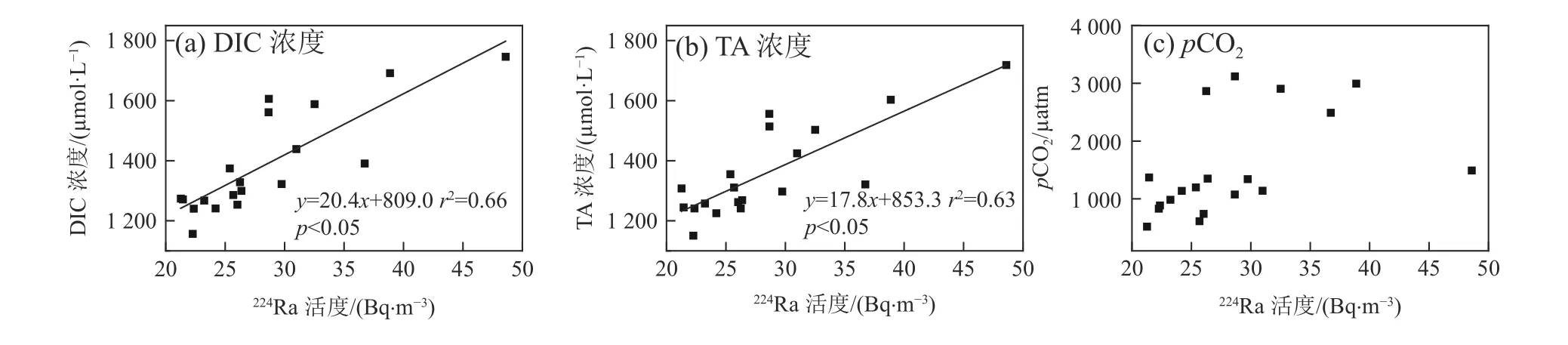
图4 DIC浓度、TA浓度、pCO2与224Ra活度之间的关系Fig. 4 Relationship between DIC concentration, TA concentration, pCO2 and 224Ra activity
与之前研究结果相比,本研究估算的孔隙水输入的DIC通量与澳大利亚亨特河口、卡奔塔利亚湾和中国九龙江河口估算的通量相差不大,相对于中国海南清澜湾的通量较小,高于帕劳巴贝达奥普火山岛红树林溪。孔隙水输入的DOC通量处于较高水平,与中国广西茅尾海估算的通量相差不大,远高于澳大利亚西港口和帕劳巴贝达奥普火山岛红树林溪(表3),源于孔隙水中高的DOC浓度。孔隙水输送的DIC通量大于DOC通量,可能是因为漳江口红树林沉积物中有丰富的有机质(Gao et al, 2021b),微生物活动促进了有机物中有机质的降解,并将大量DIC释放到含水层中(Chen et al, 2018)。也可能是由于红树林沉积物中大量有机碳快速耗氧,导致沉积物-水界面形成还原条件,大部分有机碳通过各种电子受体厌氧氧化,例如NO3−等,并转化为DIC在孔隙水中累积(Wu et al,2021)。
为了对比河流的DIC、TA和游离CO2通量,利用河流的流量与河流端元的DIC(648.1 μmol/L)、TA(548.3 μmol/L)和游离CO2浓度(100.2 μmol/L)的乘积来估算,河流输入的DIC、TA和游离CO2通量分别为7.6×105、6.4×105和1.2×105mol/d。整个红树林区域通过孔隙水输送的DIC(8.1×105mol/d)和TA(8.0×105mol/d)通量均超过河流输送通量,污水输入的DIC通量为5.9×104mol/d,远小于孔隙水排放和河流输送的DIC通量。说明在天的时间尺度上,污水排放等人为活动对潮沟DIC的影响不显著。综上,孔隙水输送是红树林潮沟水体中DIC的最主要来源。如果用孔隙水交换的单位红树林面积的碳通量乘以河道面积(28700 m2),得到的通过孔隙水从红树林向潮沟输送的DIC和DOC通量分别为2.9×104和9.5×103mol/d,红树林植被固定的碳通量(1.8×105mol/d)通过净初级生产(0.23 mol/( m2·d))(Zhu et al, 2019)乘以红树林面积(802212 m2)求得,红树林沉积物的碳埋藏量(3.2×104mol/d)由红树林面积乘以碳埋藏速率(约0.04 mol/(m2·d))(Chen et al, 2021a) 求得。因此,红树林潮沟通过孔隙水交换的溶解碳(DIC和DOC)通量(3.9×104mol/d),占红树林植被固碳量的23%,是土壤碳埋藏量的1.2倍。
通过对碳浓度和水流量在一个日循环中进行积分,估算了潮沟地表水的横向碳输送通量。涨潮期间的平均流量为26.0 m3/s,落潮期间的平均流量为16.0 m3/s,涨潮期间的水流量高于落潮期间,且涨潮时的最值(65.4 m3/s)大于落潮时的最值(38.1 m3/s)。落潮时水流量变化趋势相对于涨潮时平缓。溶解碳通量和碱度通量的变化趋势与水流量一致,表明通量由水流量控制(图5)。计算结果表明,在红树林为主的潮沟,DIC(1136 mmol/(m2·d))、TA(1349 mmol/(m2·d))、DOC(131 mmol/(m2·d))和游离CO2通量(170 mmol/(m2·d))均向下游河口净输出,面积为河道面积。DIC是地表水溶解碳横向输出的主要形式,约占溶解碳输出的90%,这与中国海南清澜湾的结果类似(以DIC形式出口的碳约占溶解碳的98%)(Wu et al, 2021)。地表水横向输送的游离CO2通量占DIC通量的18%,说明地表水主要向邻近河口输出碳酸盐碱度,与地下水输送的结果类似。通过孔隙水从红树林向潮沟输送的DIC通量为2.9×104mol/d,贡献了地表水横向输送的DIC通量(3.3×104mol/d)的89%。虽然这些碳输出通量是根据一个潮汐周期的数据进行估算,今后需要更长期的观测来准确量化输送通量,但以上结果表明孔隙水排放是红树林碳横向输送至河口的重要组成部分,应在碳收支估算中予以考虑。
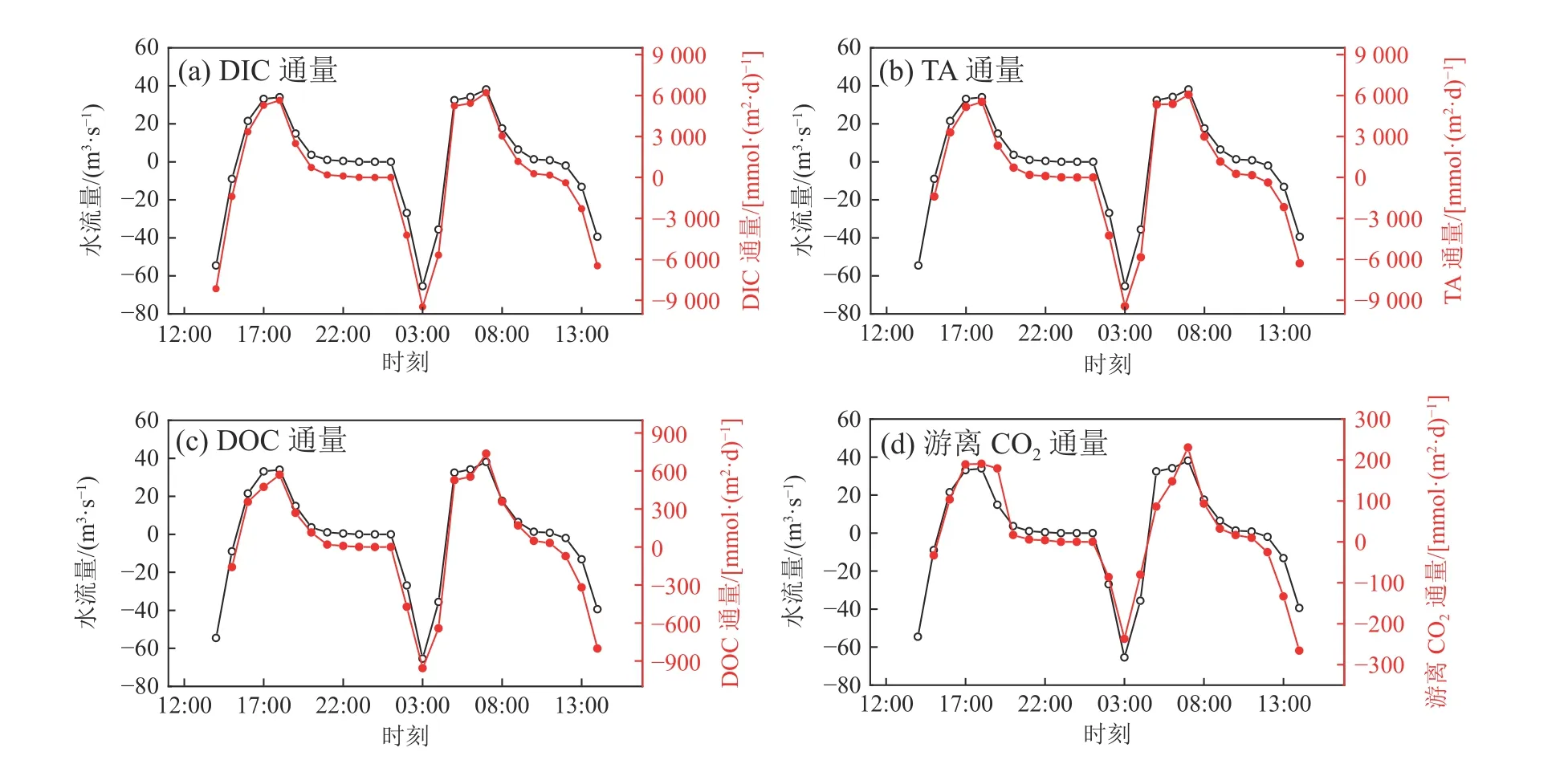
图5 时间序列观测期间碳酸盐通量与水流量之间的关系Fig. 5 Relationship between fluxes of the carbonate system and water flow during the time series study
3 结论
本研究在漳江口红树林潮沟处进行24 h 的时间序列观测,通过2种模型估算了孔隙水交换速率,量化了红树林横向输送的溶解碳和碱度通量。研究表明:①红树林通过孔隙水输入潮沟的DIC和TA通量比输送DOC和游离CO2的通量高1~2个数量级,孔隙水是红树林DIC和TA横向输出的主要途径,DIC是地表水溶解碳横向输出的主要形式。②潮沟水体是大气CO2的源,孔隙水输送至潮沟水体的游离CO2有34%经过海-气CO2交换释放到大气中。③孔隙水交换是不容忽视的重要碳汇,红树林潮沟通过孔隙水交换的溶解碳通量占红树林植被固定的碳通量的23%,是土壤碳埋藏量的1.2倍。④孔隙水排放是红树林DIC横向输送至河口的重要组成部分,红树林潮沟通过孔隙水交换的DIC通量贡献了潮沟地表水横向输送溶解DIC通量的89%。对于红树林蓝碳收支评估时应考虑孔隙水/SGD的贡献,沿海湿地系统通过孔隙水/SGD横向输送的碳可能对近海区域碳循环产生重要影响。
致谢:感谢厦门大学王桂芝教授和陈能汪教授在样品采集和测定方面提供的帮助,感谢中国海洋大学马莉、曲金燕和李方茹在测样工作中的帮助。
