烟台海岸带地区狗尾草内稳性缓解P限制
2023-10-26钟小瑛任奕炜衣华鹏
钟小瑛,任奕炜,衣华鹏*,祝 贺,高 猛
烟台海岸带地区狗尾草内稳性缓解P限制
钟小瑛1,2,任奕炜1,衣华鹏1*,祝 贺2,高 猛3
(1.鲁东大学资源与环境工程学院,山东 烟台 264025;2.中国科学院水利部成都山地灾害与环境研究所,四川 成都 610299;3.烟台大学,山东 烟台 264005)
为探究不同海岸梯度上狗尾草((L.) Beauv.)养分利用及对环境的适应策略,以烟台砂质海岸防护林下的狗尾草为研究对象,对狗尾草叶片和土壤养分含量进行分析,并探讨叶片化学计量特征和内稳性特征与土壤之间的关系.结果表明,不同海岸梯度上狗尾草叶片和土壤养分含量及化学计量特征存在一定梯度响应;研究区土壤P含量极低,狗尾草叶片P-N:P(叶片氮磷比)>16,狗尾草生长发育更多受到P元素的限制;狗尾草叶片P-C(叶片有机碳),P-P(叶片全磷),P-C:N(叶片碳氮比),P-C:P(叶片碳磷比)和P-N:P(叶片氮磷比)为绝对稳态,1/H(P-N)(叶片全氮的内稳性系数)为0.26,属于弱稳态型,与N和P元素相比,N:P更稳定,狗尾草在生长过程中按一定比例调控自身营养元素;研究区内狗尾草主要受P元素限制,但整体上内稳性较高,说明狗尾草适应性极强,可以在多变的环境中维持体内化学计量的动态平衡,缓解海岸带地区P限制的影响,在贫瘠的环境中缓慢生长.
狗尾草;内稳性;养分限制;化学计量特征;砂质土壤
生态化学计量内稳性是指植物在不同外部营养条件下维持自身养分组成相对稳定的能力[1],是生态化学计量学的理论基础和研究前提[2].当土壤养分等外界环境因子发生改变时,植物可以通过动态平衡维持自身化学组成的相对稳定,使其内部环境的变化保持在较小的范围[3].因此,内稳性能够反映植物对环境变化的生理和生化适应[4],内稳性越高,物种优势度和稳定性也越高[5].植物对土壤养分的选择性吸收和凋落物的输入影响土壤养分含量和化学计量特征[6].叶片碳(C),氮(N),磷(P)含量及其生态化学计量特征可以反映土壤养分的供给能力和植物对营养元素的需求[7-8].研究植物的内稳性特征有助于深入了解植物对环境的适应策略,深入探讨植物内稳性与生态系统功能的关系[9],其结果对评价狗尾草对环境的适应性响应以及生物多样性与初级生产力之间的相关性具有重要意义.然而目前关于植物内稳性的研究较少,研究成果也非常有限.
前人已对植物养分限制状况与内稳性之间的关系开展了研究[10-11],认为植物养分元素内稳性较弱时,该养分即为限制性养分[10],也有研究表明,限制性养分元素内稳性较强,对环境变化的响应也较为稳定[11].狗尾草((L.) Beauv.)作为C4植物适应性极强,具有极强的耐贫瘠,耐干旱和耐盐碱等抗逆特性[12].狗尾草是烟台砂质海岸防护林下草本层的优势种,不仅减缓了土壤盐碱化进程,提高了防护林生态系统的初级生产力,还丰富了防护植被的生物多样性,改善了生态环境[13].目前,国内外关于植物内稳性的研究多集中于乔木[14-15]和草地中的草本植物[16-17],关于森林生态系统中草本植物的研究相对较少,有关狗尾草在沿海应激环境(如干旱,养分限制和海风胁迫)下的养分平衡策略和生态适应性的研究未见报道.不同海岸梯度(从海岸线开始,在垂直于海岸线方向由沿海向内陆延伸形成的梯度)上,土壤盐分,土壤养分含量和土壤质地等因子具有一定的空间异质性[18-19].环境因子不同,植物养分状况与内稳性特征也会相应地发生变化.如果忽略环境变化对植物生长发育的影响,则无法深入研究植物对环境的适应性.
本文以烟台砂质海岸防护林下的狗尾草为研究对象,阐述不同海岸梯度上狗尾草叶片和土壤元素含量及化学计量特征和内稳性特征,探讨狗尾草生长特性与土壤因子之间的关系.本文主要研究以下问题:(1)不同海岸梯度上狗尾草叶片和土壤养分是否存在梯度变化?(2)狗尾草生长受N还是P限制?(3)叶片内稳性与限制元素之间存在什么关系?这将为研究狗尾草在环境变化背景下的适应策略和养分平衡机制提供新的思路,同时揭示元素相互作用与制约变化规律及稳定性特征,丰富了化学计量学理论在狗尾草管理与经营方面的应用,为更好地利用植物特性促进烟台砂质海岸恢复与建设提供理论参考.
1 研究地区与研究方法
1.1 研究区概况
研究区位于山东省烟台沿海防护林省级自然保护区牟平区段内(37°26′N~37°28′N,121°49′E~ 121°55′ E)(图1),是保护区内面积最大的核心区,物种丰富,集中,地域连片,生态系统较完整,干扰因素较少,是保持水土,抵御海浪和海风的袭击,防御海洋自然灾害的一道有效防线,生态区位十分重要.研究区濒临黄海,受季风和海洋影响显著,属暖温带东亚季风型大陆性气候,热量充足,雨量丰沛[20].研究区地势低平,土壤为弱酸性滨海砂土,其质地为疏松的中,粗砂,土壤含水量低,有机质含量极低,土壤贫瘠.研究区植被简单,草本层植物主要有狗尾草((L.) Beauv.),狗牙根((L.) Pers.),砂引草(L.),马齿苋(L.)和滨藜((Litv.) Iljin.)等.
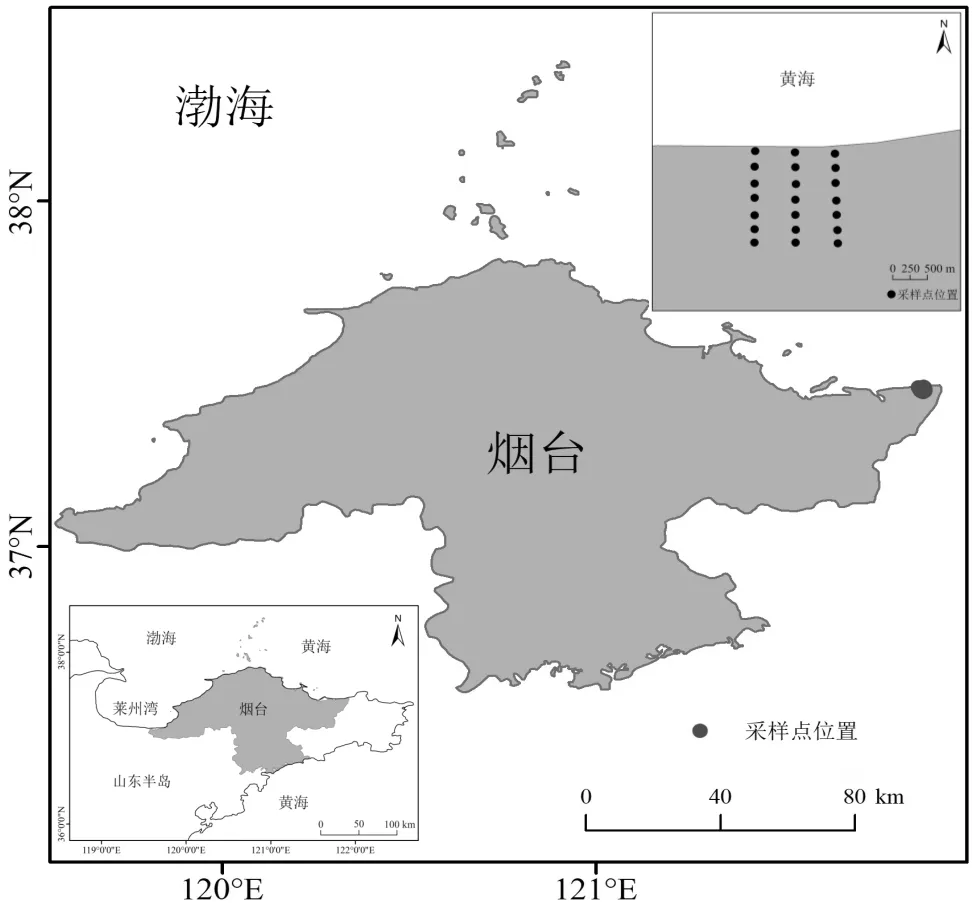
图1 狗尾草采样点分布示意
1.2 样品采集
于2021年9月在研究区进行野外样品采集工作,根据典型地段防护林面积,设置样线数和样方数.该防护林区段垂直海岸线宽度最大处约1500m,在这个空间范围内,沿垂直于海岸线方向,由沿海向内陆每隔200m设置1个采样点,左右间隔300m各设置1条平行样带,共设3条样带,各样带设7个样点,共计21个样点.各样带中海岸距离相同的采样点设为同一个海岸梯度,起始设为200m,依次为400,600, 800,1000,1200和1400m,共计7个海岸梯度,分别记为梯度1~梯度7.每个样点设置一个20m×20m的样方,在其东北,西南对角线上各设置1个1m×1m的草本样方[3].各草本样方内选取10株长势良好的狗尾草为取样对象.狗尾草叶片经去离子水冲洗后,于105℃烘箱中杀青10min,75℃烘干至恒重,冷却后采用多功能粉碎机磨碎后过0.25mm筛,然后将样品分别编号测定植物有机碳(P-C),全氮(P-N)和全磷(P-P)含量.
在每个样方内根据狗尾草根系生长状况分别使用5点取样法采集距地表0~20cm的土壤样品,并除去其中可能包含的生物残体,砾石等杂质,装入干净无杂质的密封袋中,土样原始重量不少于500g.采集的土壤样品放置在室内通风处风干,根据不同的实验要求过不同孔径筛,分别测定土壤pH值,电导率(EC),有机碳(S-C),硝酸盐氮(NO3--N),铵态氮(NH4+-N),速效磷AP),全氮(S-N)和全磷(S-P)含量.
1.3 试验方法
土壤经水土比5:1浸提后,分别用酸度计测定pH值,电导率仪测定电导率.植物和土壤样品有机碳含量均采用重铬酸钾(K2Cr2O7)—氧化外加热法测定.10.00g土样用50mL 2mol/L KCl浸提后,分别用双波长法和靛酚蓝比色法测定土壤NO3--N和NH4+-N.1.00g土样用7mL 0.03mol/L NH4F- 0.025mol/L HCl浸提后测定土壤AP.植物样品经H2SO4-H2O2消解后,定容至100mL,采用靛酚蓝比色法测定P-N含量,钼锑抗比色法测定P-P含量.1.00g土样经H2SO4-混合催化剂消解后,定容至100mL,采用靛酚蓝比色法测定S-N.1.00g土样经HClO4- H2SO4法消解后,定容至100mL,采用钼锑抗比色法测定S-P.
1.4 数据分析
Persson等[21]认为,化学计量内稳性可以用内稳性指数(H)进行量化.Sterner等[11]提出了内稳性特征模型,具体表示为:
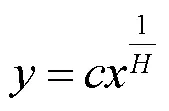
即
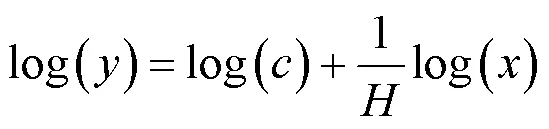
内稳性指数可以表示为:
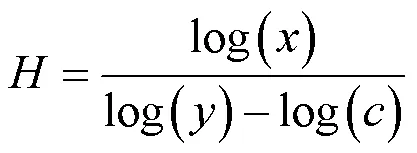
式中:为土壤C,N,P元素含量或化学计量比;为植物C,N,P元素含量或化学计量比;1/为内稳性系数,为常数.
为方便统计,多用内稳性系数即斜率1/来衡量内稳性的强弱[22].Persson等[21]经过大量研究认为,回归关系不显著(>0.05),则认为该植物为绝对稳态;回归分析显著(£0.05),则将植物按内稳性系数1/划分为4种类型:0<1/<0.25,稳态型;0.25<1/<0.5,弱稳态型;0.5<1/<0.75,弱敏感型;1/>0.75,敏感型.1/越接近0,植物养分含量越稳定,物种越趋于稳态[23].
运用SPSS 19进行数据分析和整理,利用Origin 2021进行绘图,狗尾草叶片与土壤化学计量比均采用百分比表示.采用SPSS 19中单因素ANOVA检验中的Duncan(D)进行显著性检验(<0.05),采用Origin 2021中的Pearson相关分析确定狗尾草叶片与土壤养分含量及化学计量比之间的相关性.图中数据为(平均值±标准误).
2 结果与分析
2.1 不同海岸梯度上土壤养分含量及化学计量特征
研究区土壤pH值,EC,NO3--N,NH4+-N和AP含量分别为5.74~6.10,28.90~51.84µS/cm,2.16~ 3.28mg/kg,2.57~6.69mg/kg和8.61~25.77mg/kg,平均值分别为(5.90±0.04), (38.57±3.25)µS/cm, (2.76± 0.15)mg/kg, (4.79±0.54)mg/kg和(16.26±2.44)mg/kg (图2). EC,NO3--N,NH4+-N和AP含量在不同梯度间差异显著(<0.05),pH值在不同梯度间无显著差异.从海岸梯度变化规律来看,EC和NH4+-N含量大致随海岸梯度的增加而递增,NO3--N和AP含量随海岸梯度的增加而递减,pH值在不同梯度上较为稳定.
研究区土壤S-C,S-N和S-P含量分别为6.51~ 10.21,0.20~0.40,0.04~0.08g/kg,平均值分别为(8.20± 0.43),(0.29±0.03),(0.06±0.00)g/kg(图3). S-C,S-N和S-P含量均在不同海岸梯度之间差异显著(<0.05),且均大致随海岸梯度的递增而递增,S-N和S-P含量在不同海岸梯度下变化趋势具有同步性.S-C, S-N和S-P含量均在梯度6最高,梯度1最低.S-C: N,S-C:P和S-N:P分别为26.09~35.43, 130.89~ 152.24和4.26~5.59,平均值分别为(29.58±1.28), (141.23±3.44)和(4.90±0.16).S-C: N,S-C:P和S-N:P在7个梯度间均无显著差异,S-C: N和S-C:P在不同海岸梯度下变化趋势具有同步性.
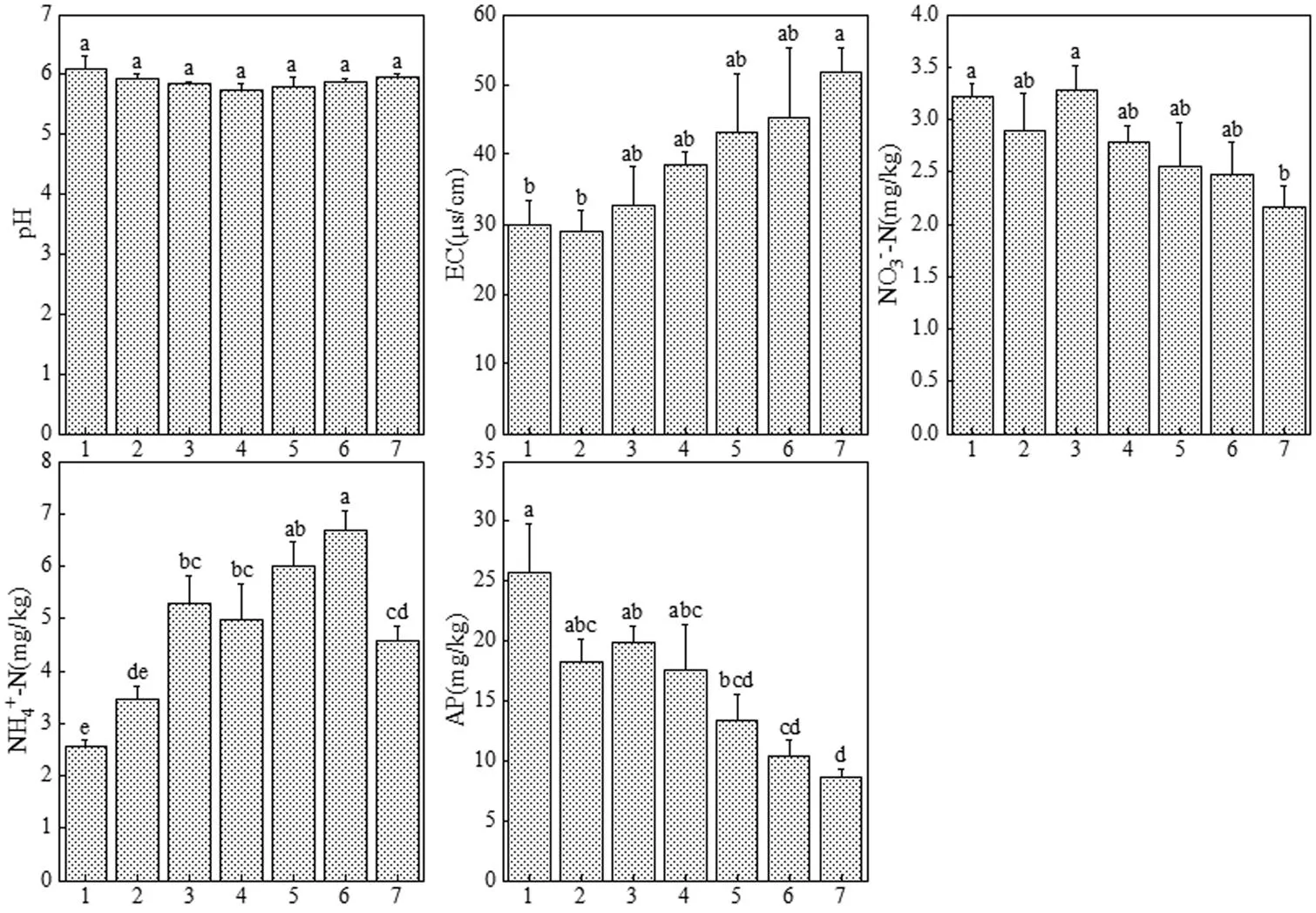
图2 不同海岸梯度上土壤pH值,EC,NO3--N,NH4+-N和AP含量
不同字母表示不同梯度间差异显著(<0.05).横轴1-7分别代表梯度1~梯度7.下同

图3 不同海岸梯度上土壤C,N,P元素含量及化学计量比
土壤C,N和P元素及化学计量比分别用S-C,S-N,S-P,S-C:N,S-C:P和S-N:P表示
2.2 不同海岸梯度上狗尾草叶片养分含量及化学计量特征
狗尾草叶片P-C,P-N和P-P含量分别为443.43~542.20,17.11~20.22和0.77~1.90g/kg,平均含量分别为(516.24±13.28),(19.06±0.46)和(1.22±0.16) g/kg(图4).P-C和P-P含量在不同海岸梯度之间差异显著(<0.05),P-N含量无显著差异.从梯度变化规律来看,P-P含量呈“V”字型变化趋势,在梯度5达到最小值.P-C:N,P-C:P和P-N:P分别为23.67~31.05, 266.24~703.35和8.93~26.46,平均值分别为(27.44± 0.91), (542.60 ±57.95)和(19.58±2.13). P-C:P,P-N:P在不同梯度间差异显著(<0.05), P-C:N不同梯度间无显著差异.P-C:P和P-N:P在不同海岸梯度上变化趋势具有一致性.
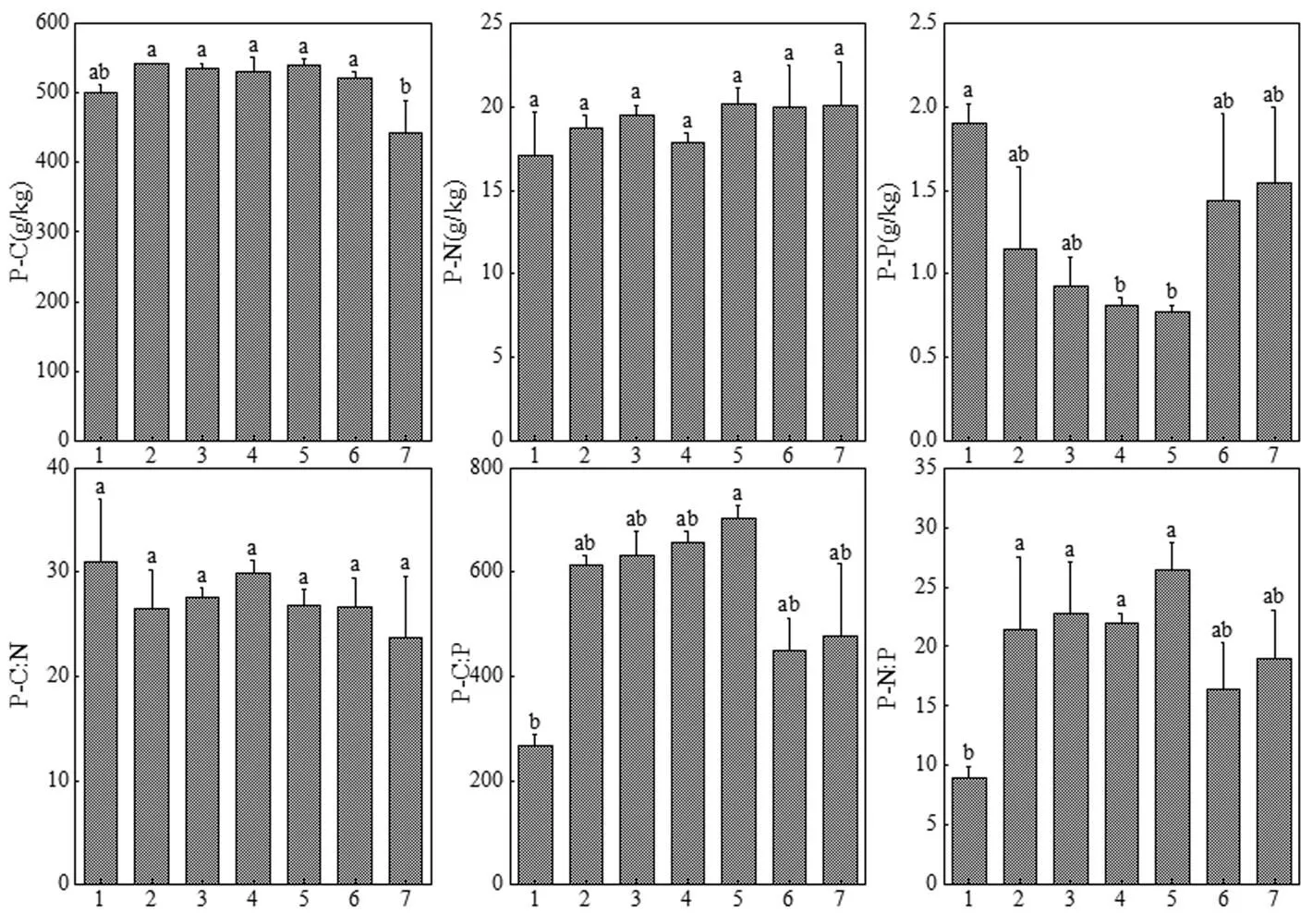
图4 不同海岸梯度上狗尾草叶片C,N,P元素含量及化学计量比
狗尾草叶片C,N和P元素及化学计量比分别以P-C,P-N,P-P,P-C:N,P-C:P和P-N:P表示
2.3 狗尾草叶片内稳性特征
由图5可知,log(P-C)与log(S-C),log(P-P)与log(S-P), log(P-C:N)与log(S-C:N),log(P-C:P)与log(S-C:P)以及log(P-N:P)与log(S-N:P)之间利用内稳性模型模拟结果不显著(>0.05),因此认为狗尾草叶片P-C,P-P,P-C:N,P-C:P和P-N:P在不同海岸梯度上为绝对稳态;而log(P-N)与log(S-N)之间的模拟结果显著(<0.05),内稳性系数为0.26,0.25<0.26<0.5,狗尾草叶片N元素属于弱稳态型.
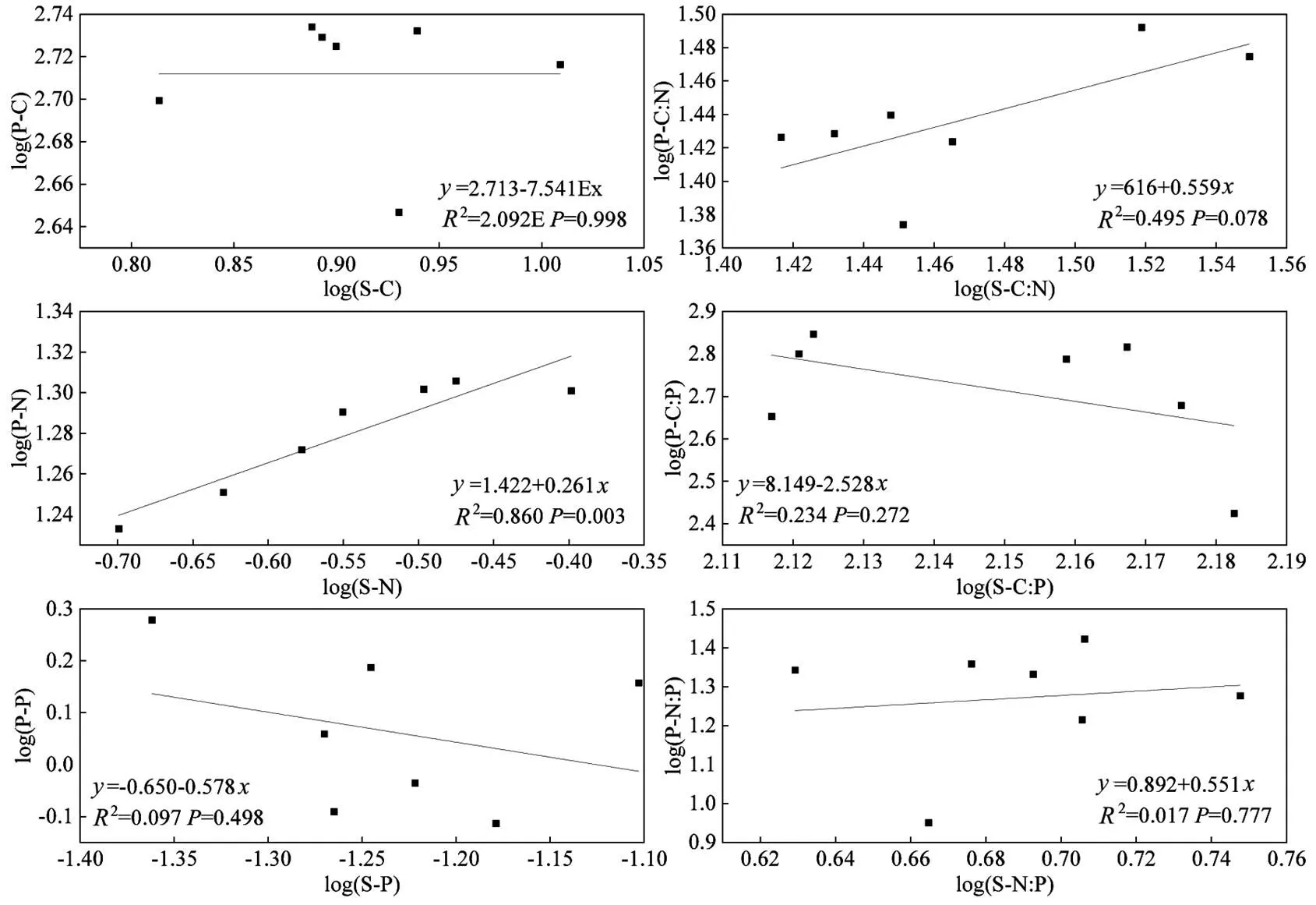
图5 狗尾草叶片内稳性特征
狗尾草叶片P-C,P-N和P-P元素及化学计量比的对数分别用log(P-C), log(P-N),log(P-P),log(P-C:N),log(P-C:P)和log(P-N:P)表示.土壤S-C,S-N和S-P元素及化学计量比的对数分别用log(S-C),log(S-N),log(S-P),log(S-C:N),log(S-C:P)和log(S-N:P)表示
2.4 狗尾草叶片元素含量及化学计量比与土壤的关系
由相关性分析可看出,狗尾草叶片元素含量及化学计量比和土壤元素含量及化学计量比具有一定的相关性.S-C与P-N呈显著正相关关系(<0.05); S-N与P-N呈极显著正相关关系(<0.01);S-P与P-N呈显著正相关关系(<0.05);S-C:N与P-N呈极显著负相关关系(<0.01);S-N:P与P-N和P-C:N分别呈显著正相关关系(P<0.05)和极显著负相关关系(<0.01);pH与P-P呈极显著正相关关系(<0.01),与P-C:P和P-N:P均呈显著负相关关系(<0.05); NH4+-N与P-N呈显著正相关关系(<0.05);AP与P-N呈极显著负相关关系(<0.01),与P-C:N呈显著正相关关系(<0.05).
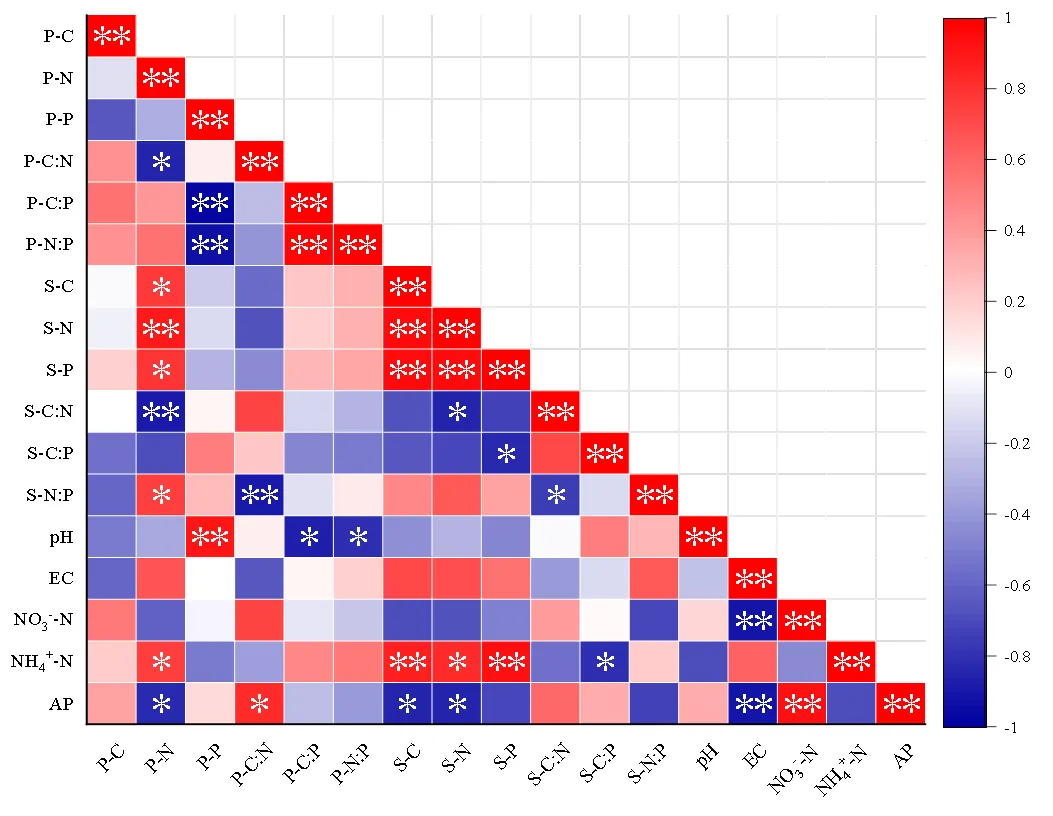
图6 狗尾草叶片与土壤的相关性
红色表示正相关,蓝色表示负相关,颜色强度与相关系数成正比;*表示显著相关(<0.05),**表示极显著相关(<0.01)
3 讨论
3.1 不同海岸梯度上土壤养分含量变化特征
土壤中营养元素的持续供给是植物生长发育的重要前提[24],其在植物生长和生理调节中起着重要作用[2].但是实际上全球范围内存在一定程度上土壤养分供给的N限制或者P限制[14,25],尤其是在海岸带地区往往由于植被类型单一或者群落演替不完整导致植物凋落物较少,减少了土壤C和N等营养元素的来源[26];此外,土壤颗粒组成也是影响土壤营养元素的重要因素,海岸带区域砂质土壤中矿质颗粒表面的离子结合位点极其有限,对有机质和养分的固持能力极低,导致土壤养分难以大量积累[27-28].因此,研究区不同海岸梯度上土壤S-C (8.20±0.43)g/kg,S-N(0.29±0.03)g/kg,S-P(0.06±0.00)g/kg含量均低于全球陆地土壤平均C(25.71g/kg), N(2.10g/kg),P(0.36g/kg)含量[27]和中国陆地土壤C(24.46g/kg),N(1.88g/kg),P(0.23g/kg)含量[29],养分含量整体偏低.所以仅仅关注S-C,S-N和S-P等元素的含量并不能真正反应土壤中营养元素与植物生长发育之间的关系.
土壤C:N:P是反映土壤养分有效性,预测土壤养分饱和状态的重要指标[7].研究区土壤S-C:N (29.58±1.28),S-C:P(141.23±3.44)远高于中国陆地土壤平均C:N(11.90),C:P(61.00)[29].C:N可以预测有机质的分解速率,反映土壤质量[30].当土壤C:N<25时表明土壤有机质腐殖化程度高,易于有机质矿化[31].研究区土壤S-C:N>25(图3),这可能是因为研究区内主要凋落物为黑松(Parl.)针叶,黑松针叶含有较多木质素和纤维素导致凋落物难以分解,从而导致土壤有机质腐质化程度低[28].土壤C:N越高肥力越低,C:P越高磷有效性越低[32].较高的S-C:N,S-C:P表明研究区土壤肥力较低,土壤贫瘠.土壤N:P反映了N元素的饱和状态,可用于判断土壤养分限制阈值[26].研究区土壤S-N:P(4.90±0.16)均低于全球森林土壤(6.60)[25]和中国陆地土壤N:P平均值(5.20)[29].主要原因在于生物的固N量与土壤N:P呈负相关,N:P越高则能被植物吸收利用的有效N越低[33].研究区土壤较低的S-N:P说明土壤中N元素相对缺乏,但生物固N量较高,能被狗尾草吸收利用的有效性N相对充足,狗尾草生长未受N限制.
不同海岸梯度上S-C,S-N和S-P含量均存在空间异质性,且大致随海岸梯度的递增而递增(图3),这可能是因为随海岸梯度增加,人为扰动减少,群落演替逐渐完整,地表凋落物增多,分解归还给土壤的养分增加.对同一海岸环境的变化S-C:N和S-C:P总是同步响应的,表现出几乎相同的变化规律,这是因为S-N和S-P含量在不同海岸梯度下变化趋势具有同步性(图3),S-N和S-P含量的变化决定了S-C:N和S-C:P水平.S-C:N,S-C:P和S-N:P在不同海岸梯度上无显著差异,说明与单一元素相比,元素计量比受海岸梯度影响较小相对比较稳定.
3.2 狗尾草生长限制性养分分析
C,N和P元素是植物的基本组成成分,在植物生长和生理调节中起着重要作用[2].本研究中狗尾草叶片P-C含量(516.24±13.28)g/kg高于Elser观测的全球植物平均C含量(464.00)g/kg[34],狗尾草作为C4植物具有极高的光合速率,有机物积累能力高,能够储存更多的C来抵抗环境胁迫[35].叶片P-N(19.06± 0.46)g/kg,P-P(1.22±0.16)g/kg含量均低于全球植物叶片平均N和P含量(分别为20.60g/kg,1.49g/kg)[34],也低于Han等研究得出的中国陆地植物叶片平均N,P含量(分别为20.20g/kg,1.45g/kg)[36],这可能是因为草本植物所需的营养元素主要来源于土壤[37],而研究区土壤S-N和S-P含量偏低,植物和土壤的N,P含量通常在生态系统尺度上耦合[36],导致狗尾草叶片N,P含量偏低.从海岸梯度变化规律来看,C含量对不同海岸梯度的响应较为明显,这是因为不同海岸梯度上植被覆盖率不同,黑松林下狗尾草获得的光照强度不同,叶片合成的有机物变化,从而导致不同海岸梯度之间差异显著;N含量大致随海岸梯度的增加而递增,与土壤的递变规律相似,这可能是由于植物体内的N元素主要通过根系从土壤中吸收[38];P含量呈“V”字型变化趋势,变化规律具有独特性.
植物C:N:P化学计量可以反映植物的养分利用效率及其对环境变化的响应和适应[39].植物C:N和C:P可以反映植物的固碳能力和生长状况[40].狗尾草叶片P-C:N(27.44±0.91),P-C:P(542.60±57.95)均高于全球植物平均叶片C:N (22.50),C:P(469.16)[34],较高的P-C:N和P-C:P反映了狗尾草具有较高的N和P元素利用效率和固碳能力[41],表明狗尾草可以通过调节自身化学计量比来适应贫瘠的环境.一般研究表明,当植物N:P>16时其生长主要受P限制;当植物N:P<14时其生长受N限制;当植物N:P在14和16之间时,植物生长受N和P共同限制[42].本研究狗尾草叶片P-N:P(19.58±2.13)>16(图4),表明狗尾草生长可能受到P元素限制,这与Han[36]研究得出的中国大多数土壤中植物可能受P限制一致.不同海岸梯度上P-C:P,P-N:P存在空间异质性,这可能是因为P-C,P-P含量在不同海岸梯度之间差异显著(P <0.05),在P-N含量基本保持不变情况下,P-C,P-P含量的变化决定了P-C:P,P-N:P水平.P-C:N在不同海岸梯度上无显著差异,说明P-C:N受海岸梯度影响较小,相对比较稳定,这可能是因为P-N含量在不同海岸梯度上无显著差异.
3.3 叶片内稳性与限制元素的关系
如前文所述,研究区内狗尾草生长可能存在P限制,但其可以通过多种生理生化机制调节限制元素的有效性和利用效率[43],以在贫瘠环境下缓解P的限制以保持缓慢生长.主要原因在于内稳性高的植物采用消极的养分利用策略应对营养下降,养分利用方式较为保守,使其在贫瘠的环境中也能维持植物的缓慢生长[9],从而具有较高的生产力和群落稳定性.本研究中狗尾草叶片P-C,P-P,P-C:N,P-C:P和P-N:P为绝对稳态(图5),说明狗尾草对环境的变化有较强的适应能力,能够调节自身养分分配策略维持其正常生长,并使其在养分贫瘠的环境中具有较高的稳定性和优势度,最终成为适宜生长的优势种.叶片1/H(P-N)为0.26均低于闽江河口湿地3种植物(互花米草,短叶茳芏和芦苇1/H(N)分别为0.64,0.71,0.50)[44],这可能是由于研究区位于海岸带防护林地区,较河口湿地环境较为稳定.叶片1/H (P-N)属于弱稳态型(图5),狗尾草叶片中的N含量对环境变化的响应比P更为敏感.一般情况下,N元素比P元素更为稳定[45],但狗尾草叶片对P元素比对N元素具有更强的调控能力,P元素在狗尾草体内的含量具有相对稳定性,对环境变化的响应也较为稳定,在一定程度上证实了限制性养分元素稳定性假说[11].但是在判断植物内稳性强弱时,用1/H(N:P)表示植物的内稳性比1/H(N)或1/H(P)更为接近植物维持其自身动态平衡的真实能力,因为化学计量动态平衡反映了生长期间植物对养分的消耗和储存能力[46].狗尾草叶片内稳性指数H(P-N:P)>H(P-N) (图5),元素计量比的内稳性高于元素本身,这说明与N,P元素相比,N:P更稳定,受外界环境等因素的影响小,这是因为植物体中N含量远高于P含量,但N含量的累积通常伴随着P含量的增加,其变化具有协同性[27],说明狗尾草对其自身内稳性的调节主要体现在N:P而不是每种元素的含量上[47],狗尾草在生长过程中按一定比例调控自身营养元素,与闽江河口湿地互花米草一致[44].因此,用H(N:P)表示植物的内稳性更为接近植物维持其自身内稳性的真实能力[9],虽然研究区内狗尾草生长可能受到P限制,但狗尾草自身生长策略的调控缓解了这一矛盾.
植物体内养分含量体现了植物对环境的适应策略,土壤有效养分状况则反应了植物养分的供给需求[6].狗尾草叶片P-C含量与土壤养分含量均不存在显著相关关系(图6),表明狗尾草体内C元素的主要来源不是土壤,而是其自身的光合作用[48].本研究中狗尾草叶片P-N含量与土壤NH4+-N呈显著正相关,虽然NO3--N和NH4+-N是植物吸收氮素的主要形态[49],但是在酸性土壤中NH4+-N是其主要的无机氮形式[50],C4禾草偏好NH4+-N的吸收[51],所以土壤中NH4+-N可以满足狗尾草的生长需求.叶片P-N含量与土壤S-N呈极显著正相关(图6),与土壤S-P,S-N:P呈显著正相关,与S-C:N呈极显著负相关,与AP呈显著负相关的相关关系,这意味着植物对N的需求影响了土壤有效N和有效P的含量.与N不同,研究区内可能存在P限制,但研究结果表明狗尾草叶片P-P含量与土壤S-P含量并未表现出显著相关性,表明狗尾草叶片P含量并非由土壤P直接决定,这可能与狗尾草在P限制下的特殊调节机制有关.植物元素含量及化学计量比不仅受土壤养分有效性的影响,还与植物维持不同功能所需的养分需求有关[52].狗尾草自身具有较强的内稳性,即使在土壤可能存在P限制条件下,也可以通过养分调控机制形成独特的养分特征和生理生态习性,降低P限制对其生长发育的不利影响,增强对贫瘠环境的适应能力,并逐渐演化成物种的基本属性.
4 结论
4.1 狗尾草叶片和土壤养分存在一定梯度变化.土壤S-N和S-P,S-C:N和S-C:P,植物P-C:P和P-N:P在不同海岸梯度下变化趋势具有同步性.植物P-P呈“V”字型,变化趋势具有独特性.
4.2 研究区土壤P含量极低,狗尾草叶片P-N:P>16,狗尾草生长发育可能受到P元素限制.
4.3 狗尾草叶片P-C,P-P,P-C:N,P-C:P和P-N:P为绝对稳态,1/H(P-N)为0.26,属于弱稳态型,狗尾草限制性养分内稳性较强,对环境变化的响应较为稳定.
[1] Wang J, Wang J, Wang L, et al. Does stoichiometric homeostasis differ among tree organs and with tree age? [J]. Forest Ecology and Management, 2019,453(C):117637-117637.
[2] Li Y F, Li Q, Liu H, et al. Ecological stoichiometry-based study of the influence of soil saline-alkali stress on nutrient homeostasis in L. chinensis [J]. Ecotoxicology and Environmental Safety, 2018,165: 243-249.
[3] 海旭莹,董凌勃,汪晓珍,等.黄土高原退耕还草地C,N,P生态化学计量特征对植物多样性的影响[J]. 生态学报, 2020,40(23):8570- 8581. Hai X Y, Dong L B, Wang X Z, et al. Effects of carbon nitrogen and phosphorus ecological stoichiometry characteristics on plant diversity since returning farmland to grassland on the Loess Plateau. [J]. Acta Ecologica Sinica, 2020,40(23):8570-8581.
[4] Hessen D O, Agren G I, Anderson T R, et al. Carbon sequestration in ecosystems: The role of stoichiometry [J]. Ecology, 2004,85(5): 1179-1192.
[5] 张仁懿,史小明,李文金,等.亚高寒草甸物种内稳性与生物量变化模式[J]. 草业科学, 2015,32(10):1539-1547. Zhang R Y, Shi X M, Li W J, et al. Response of species homeostasis and biomass on a sub-alpine grassland [J]. Pratacultural Science, 2015,32(10):1539-1547.
[6] Liu R S, Wang D M. Soil C, N, P and K stoichiometry affected by vegetation restoration patterns in the alpine region of the Loess Plateau, Northwest China [J]. PloS one, 2020,15(11):e0241859-e0241859.
[7] Ding D D, Arif M, Liu M H, et al. Plant-soil interactions and C:N:P stoichiometric homeostasis of plant organs in riparian plantation [J]. Frontiers in Plant Science, 2022,13(13):979023-979023.
[8] 王维奇,王 纯,刘白贵.盐度对湿地枯落物分解过程中碳氮磷化学计量比的影响 [J]. 中国环境科学, 2012,32(9):1683-1687. Wang W Q, Wang C, Liu B G. Effect of salinity on carbon, nitrogen and phosphorus stoichiometry during the decomposition of wetland litter. [J]. China Environmental Science, 2012,32(9):1683-1687.
[9] 张婷婷,刘文耀,黄俊彪,等.植物生态化学计量内稳性特征[J]. 广西植物, 2019,39(5):701-712. Zhang T T, Liu W Y, Huang J B, et al. Characteristics of plant ecological stoichiometry homeostasis [J]. Guihaia, 2019,39(5):701- 712.
[10] Garnier E. Interspecific variation in plasticity of grasses in response to nitrogen supply [M]. Cambridge University Press, 1998,14:15582.
[11] SternerR W, ElserJ J. Ecological Stoichiometry: the Biology of Elements from Molecules to the Biosphere [M]. Princeton University Press, 2002.
[12] Valeriano F R, de Moura S M, Travassos Lins J, et al. Evaluation of Setaria viridis responses to salt treatment and potassium supply: A characterization of three contrasting accessions [J]. Brazilian Journal of Botany, 2021,44:821-836.
[13] Chen C, Hong T, Lin Y, M et al. Spatial variation of soil respiration in a coastal protection forest in southeastern China [J]. Journal of Soil and Sediments, 2017,17(9):2284-2295.
[14] Bai X J, Wang B R, An S S, et al. Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China [J]. Catena, 2019,183(C):104186-104186.
[15] Wang J Y, Wang J N, Guo W H, et al. Stoichiometric homeostasis, physiology, and growth responses of three tree species to nitrogen and phosphorus addition [J]. Trees, 2018,32(5):1377-1386.
[16] Yu Q, Chen Q S, Elser J J, et al. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability [J]. Ecology Letters, 2010,13(11):1390-1399.
[17] Yu Q, Wilcox K, Pierre K L, et al. Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change [J]. Ecology, 2015,96(9):2328-2335.
[18] 吴锡麟,叶功富,张尚炬,等.不同海岸梯度上短枝木麻黄小枝金属元素含量及其再吸收率动态[J]. 应用与环境生物学报, 2011,17(5): 645-650. Wu X L, Ye G F, Zhang S J, et al. Contents of Some Mineral Elements and Their Resorption Effi ciencies in Casuarina equisetifolia Branchlets Across a Coastal Gradient [J]. Chinese Journal of Applied and Environmental Biology, 2011,17(5):645-650.
[19] Zhang L H, Lin Y M, Ye G F, et al. Changes in the N and P concentrations, N:P ratios, and tannin content in Casuarina equisetifolia branchlets during development and senescence [J]. Journal of Forest Research, 2008,13(5):302-311.
[20] Zhou D, Shi P, Wu X Q, et al. Effects of urbanization expansion on landscape pattern and region ecological risk in Chinese coastal city: A case study of Yantai City [J]. The Scientific World Journal, 2014,2014:821781.
[21] Persson J, Fink P, Goto A, et al. To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs [J]. Oikos, 2010,119(5):741-751.
[22] Hood J M, Sterner R W, Donald L D, et al. Diet mixing: Do animals integrate growth or resources across temporal heterogeneity? [J]. The American naturalist, 2010,176(5):651-663.
[23] Song H X, Huang J J, Ge L M, et al. Interspecific difference in N:P stoichiometric homeostasis drives nutrient release and soil microbial community composition during decomposition [J]. Plant and Soil, 2020,452(1/2):29-42.
[24] Hu C, Li F, Xie Y H, et al. Soil carbon, nitrogen, and phosphorus stoichiometry of three dominant plant communities distributed along a small-scale elevation gradient in the East Dongting Lake [J]. Physics and Chemistry of the Earth, 2016,103:28-34.
[25] Cleveland C C, Liptzin D. C:N:P Stoichiometry in Soil: Is There a "Redfield Ratio" for the Microbial Biomass? [J]. Biogeochemistry, 2007,85(3):235-252.
[26] Wu X, Niu Y B, Xun M Y, et al. Soil carbon, nitrogen, and phosphorus storages and their stoichiometry due to mixed afforestation with hippophae rhamnoides in the Loess Hilly Region, China [J]. Forests, 2021,12(12):1718-1718.
[27] Wei Y J, Dang X H, Wang J, et al. Response of C:N:P in the plant-soil system and stoichiometric homeostasis of Nitraria tangutorum leaves in the oasis-desert ecotone, Northwest China [J]. Journal of Arid Land, 2021,13(9):934-946.
[28] 任奕炜,衣华鹏,钟小瑛.沙质海岸防护林下土壤养分特征研究[J]. 防护林科技, 2022,(5):1-6,28. Ren Y W, Yi H P, Zhong X Y. Research on soil nutrient characteristics under sandy coastal shelterbelts [J]. Protection Forest Science and Technology, 2022,(5):1-6,28.
[29] Tian H Q, Chen G S, Zhang C, et al. Pattern and variation of C:N:P ratios in China's soils: A synthesis of observational data[J]. Biogeochemistry, 2010,98(1/3):139-151.
[30] Liu X, Li L H, Wang Q, et al. Land-use change affects stocks and stoichiometric ratios of soil carbon, nitrogen, and phosphorus in a typical agro-pastoral region of northwest China [J]. Journal of Soils and Sediments, 2018,18(11):3167-3176.
[31] Paul E A, Clark F E. Soil microbiology and biochemistry [M]. Academic Press, 1996.
[32] Jiang Y F, Guo X. Stoichiometric patterns of soil carbon, nitrogen, and phosphorus in farmland of the Poyang Lake region in Southern China [J]. Journal of Soils and Sediments, 2019,19(10):3476-3488.
[33] 杨 慧,涂春艳,李青芳,等.区次生林地不同地貌部位土壤C,N,P化学计量特征[J]. 南方农业学报, 2015,46(5):777-781. Yang H, Tu C Y, Li Q F, et al. Analysis of C, N and P stoichiometry of secondary forest indifferent landforms in karst area [J]. Journal of Southern Agriculture, 2015,46(5):777-781.
[34] Elser J J, Fagan W F, Denno R F, et al. Nutritional constraints in terrestrial and freshwater food webs [J]. Nature, 2000,408(6812): 578-580.
[35] 张 晶,左小安,吕 朋,等.科尔沁沙地典型草地植物功能性状及其相互关系[J]. 干旱区研究, 2018, 35(1):13743. Zhang J, Zuo X A, Lv P, et al. Functional Traits and Interrelations of Dominant Plant Species on Typical Grassland in the Horqin Sandy Land, China [J]. Arid Zone Research, 2018,35(1):13743.
[36] Han W X, Fang J Y, Guo D L, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China [J]. New Phytologist, 2005,168(2):377-385.
[37] 刘艳艳.武夷山自然保护区不同海拔甜槠林细根结构与养分的关系研究 [D]. 福建农林大学, 2014. Liu Y Y. Relationship between fine roots structure and soil nutrient ofForest among different altitudes in Mt.Wuyi Nature Reserve [D]. Fujian Agriculture and Forestry University, 2014.
[38] 何亚婷,刘文治,党高弟,等.秦岭亚高山草甸30种草本植物的碳,氮分布研究 [J]. 草业科学, 2008,25(10):1-5. He Y T, Liu W Z, Dang G D, et al. Qinling Mountains; subalpine meadow; carbon content; Nitrogen content [J]. Pratacultural Science, 2008,25(10):1-5.
[39] Jiang J, Lu Y C, Chen B B, et al. Nutrient resorption and C:N:P stoichiometry responses of a pinus massoniana plantation to various thinning intensities in Southern China [J]. Forests, 2022,13(10):1699.
[40] 杨路寒,王 训,王定勇.基于碳氮化学计量探究植被格局变化对土壤汞累积的影响 [J]. 中国环境科学, 2023,43(6):2993-3001. Yang L H, Wang X, Wang D Y. Effect of vegetation pattern change on soil mercury accumulation based on carbon and nitrogen stoichiometry [J]. China Environmental Science, 2023,43(6):2993-3001.
[41] Viciedo D O, Prado R D M, Martínez C A, et al. Short-term warming and water stress affect. stoichiometric homeostasis and biomass production [J]. Science of the Total Environment, 2019,681:267-274.
[42] Koerselman W, Meuleman A F M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation [J]. Journal of Applied Ecology, 1996,33(6):1441-1450.
[43] 刘明辉,谢婷婷,袁中勋,等.三峡水库消落带适生树种落羽杉(Taxodium distichum)叶片-细根碳/氮/磷生态化学计量特征[J]. 湖泊科学, 2020,32(6):1806-1816. Liu M H, Xie T T, Yuan Z X, et al. Carbon, nitrogen, and phosphorus ecological stoichiometric characteristics in leaves andfine roots of Taxodium distichum in the water-level fluctuation zone of the Three Gorges Reservoir region [J]. Journal of Lake Sciences, 2020,32(6): 1806-1816.
[44] 蒋利玲,何 诗,吴丽凤,等.闽江河口湿地3种植物化学计量内稳性特征 [J]. 湿地科学, 2014,12(3):293-298. Jiang L L, He S, Wu L F, et al. Characteristics of stoichiometric homeostasis of three plant species in wetlands in Minjiang Estuary [J]. Wetland Science, 2014,12(3):293-298.
[45] 原雅楠,李正才,王 斌,等.不同林龄榧树根,枝,叶的C,N,P化学计量及内稳性特征[J]. 南京林业大学学报(自然科学版), 2021,45(6): 135-142. Yuan Y N, Li Z C, Wang B, et al. Ecological stoichiometry in leaves, branches and roots ofwith different forest ages and its stoichiometric homoeostasis [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2021,45(6):135-142.
[46] Blouin M, Mathieu J, Leadley P W. Plant homeostasis, growth and development in natural and artificial soils [J]. Ecological Complexity, 2012,9(Feb.):10-15.
[47] 王雪梅,闫帮国,史亮涛,等.车桑子幼苗生物量分配与叶性状对氮磷浓度的响应差异[J]. 植物生态学报, 2020,44(12):1247-1261. Wang X M, Yan B G, Shi L T, et al. Different responses of biomass allocation and leaf traits of Dodonaea viscosa to concentrations of nitrogen and phosphorus [J]. Chinese Journal of Plant Ecology, 2020,44(12):1247-1261.
[48] 钟小瑛,任奕炜,衣华鹏.狗尾草叶片N、P、K养分重吸收与化学计量比的偶联关系 [J]. 植物科学学报, 2023,41(2):193-203. Zhong X Y, Ren Y W, Yi H P. Coupling relationship between nutrient resorption and stoichiometric ratios of N, P, and K in(L.) Beauv. leaves [J]. Plant Science Journal, 2023,41(2):193-203.
[49] Kirkby E A, Knight A H. Influence of the level of nitrate nutrition on ion uptake and assimilation, organic Acid accumulation, and cation-anion balance in whole tomato plants [J]. Plant Physiology, 1977,60(3):349-353.
[50] Zhang J B, Cai Z C, Zhu T B, et al. Mechanisms for the retention of inorganic N in acidic forest soils of southern China [J]. Scientific Reports, 2013,3(1):2342.
[51] Taylor S H, Hulme S P, Rees M, et al. Ecophysiological traits in C3 and C4 grasses: a phylogenetically controlled screening experiment [J]. New Phytologist, 2010,185(3):780-791.
[52] Tian D S, Reich P B, Chen H Y H, et al. Global changes alter plant multi‐element stoichiometric copling [J]. New Phytologist, 2018, 221(2):807-817.
Homeostasis of(L.) Beauv. in Yantai coastal zone alleviated P limitation.
ZHONG Xiao-ying1,2, REN Yi-wei1, YI Hua-peng1*, ZHU He2, GAO Meng3
(1.College of Resources and Environmental Engineering, Ludong University, Shandong Yantai 264025, China;2.Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Sichuan Chengdu 610299, China;3.Yantai University, Shandong Yantai 264005, China)., 2023,43(10):5498~5507
In order to explore the nutrient utilization and environmental adaptation strategies of(L.) Beauv. on different coastal gradients, this paper analyzed the nutrient content ofleaves and soil under the sandy coastal shelter forest in Yantai, and discussed the relationship between leaf stoichiometry characteristics and endostabilizing characteristics and soil. The results showed that there was a certain response in the leaf and soil nutrient content and stoichiometric characteristics ofon different coastal gradients. The soil P content in the study area was extremely low, and more specifically the leaves ofgrass P-N: P(leaf nitrogen-phosphorus ratio)>16. The growth and development ofwere more limited by the P element. Moreover, P-C (leaf organic carbon), P-P (leaf total phosphorus), P-C:N (leaf carbon-nitrogen ratio), P-C:P (leaf carbon-phosphorus ratio) and P-N:P(the leaf nitrogen-phosphorus ratio) were all in absolute steady state. 1/H(P-N) (the internal stability coefficient of leaf total nitrogen) was 0.26, which belonged to the weak steady state type, but N:P was more stable than N and P elements. It is concluded thatregulate its nutrients in a certain proportion during the growth process. In the study area,was mainly limited by the P element, but the overall internal stability was high indicating thatwas highly adaptable in the coastal environment. It could maintain the dynamic balance of stoichiometry in vivo in a changeable environment, and alleviate the influence of P restriction in coastal areas with a low growing rate.
;homeostasis;nutrient restriction;stoichiometry characteristics;sandy soil
X173
A
1000-6923(2023)10-5498-10
2023-04-03
国家自然科学基金资助项目(31570423)
* 责任作者, 教授, huapengyi@sina.com
钟小瑛(1998-),女,山东潍坊人,鲁东大学硕士研究生,主要从事河口海岸环境演变与整治.zxy736284@163.com.
钟小瑛,任奕炜,衣华鹏,等.烟台海岸带地区狗尾草内稳性缓解P限制 [J]. 中国环境科学, 2023,43(10):5498-5507.
Zhong X Y, Ren Y W, Yi H P, et al. Homeostasis of(L.) Beauv. in Yantai coastal zone alleviated P limitation [J]. China Environmental Science, 2023,43(10):5498-5507.
