寡雄腐霉对禾谷镰刀菌防效及其产孢诱导剂筛选
2023-10-19王子洋熊雨洁冯发运余向阳朱耀华张雷刚
王子洋, 熊雨洁, 冯发运, 余向阳, 朱耀华, 张雷刚
(1.江苏大学食品与生物工程学院,江苏镇江 212013; 2.江苏省农业科学院农业资源与环境研究所,江苏南京 210014;3.河南省漯河市源汇区农业技术推广站,河南漯河 462000)
禾谷镰刀菌(Fusariumgraminearum)是引起小麦赤霉病(Fusariumhead blight,FHB)的主要致病菌,可在小麦多个生长时期和部位侵染致病,不但造成产量损失,还会在麦粒中积累多种真菌毒素,危害食品安全[1]。2000—2018年期间,我国每年有超过5.4万km2、约占总产量23%的小麦受该病影响。由于缺乏有效的抗病品种和生防产品,目前仍以化学药剂防治为主[2]。
生防真菌因具有广谱的生防作用、货架期长等特性,是目前应用较为广泛的一类生防制剂,如酵母菌(Saccharomycesspp.)、木霉菌(Trichodermaspp.)、绿僵菌(Metarhizumspp.)等[3]。寡雄腐霉(Pythiumoligandrum)属真菌门卵菌纲霜霉目腐霉科腐霉属,是一种强攻击性重寄生微生物,广泛存在于植物根际与土壤中[4]。作为具有优良拮抗效果的生防真菌,寡雄腐霉可抵抗多种病原菌对植物的侵害,包括马铃薯炭疽病[5]、番茄根腐病[6]、草莓根腐病[7]、辣椒炭疽病[8]、葡萄霜霉病[9]、水稻立枯病[10]等,是一种具有应用潜力与价值的生防因子。目前,针对寡雄腐霉生防机制的研究众多,主要发现其能够:(1)通过重寄生作用直接攻击病原菌;(2)诱导植物产生系统抗性;(3)分泌抗生物质;(4)促进植物生长,间接抵抗病原菌;(5)与病原菌竞争营养与空间等多种手段来抑制病原菌[11]。重寄生作用是寡雄腐霉的主要生防手段,通过直接攻击其他病原菌、产生细胞壁裂解酶并分泌抗生物质来杀死病原菌。Benhamou等发现,寡雄腐霉可与土壤中多种植物病原菌发生互作,并通过扫描电镜发现在寡雄腐霉与其他病原菌接触后会导致病原菌菌丝结构发生改变,病原菌菌丝被寡雄腐霉入侵,细胞膜潜在渗透位点发生改变[12]。
寡雄腐霉卵孢子是其能够抑制病原菌侵染和促进植物生长的主要作用成分,常见的制备方法是直接将寡雄腐霉菌碟接入液体培养基中以获得菌丝体,然后通过搅打或振摇的方式使孢子脱离菌丝并游离至水中[8],此方法获得的寡雄腐霉卵孢子数量和效率还有待提高。陈晨等发现,在深绿木霉(Trichodermaatroviride)液体发酵过程中添加 127.4 mmol/L 钙离子,其产孢量显著提升[13]。梁玎玎等采用控制变量法筛选出了适合草茎点霉SYAU-06菌株产孢的培养基和诱导方法,鸭跖草汁的添加缩短了孢子器的形成时间[14]。旷文丰等的研究表明,通过优化光照、营养成分等发酵和后处理工艺是提高木霉菌生防制剂产业化生产能力的重要策略[15]。本研究旨在通过离体和活体验证寡雄腐霉卵孢子对禾谷镰刀菌的防效,并利用正交试验获得产孢诱导剂的最优配方,从而增加寡雄腐霉菌丝体数量并诱导其高效产孢,以期为寡雄腐霉生物防治产品研发提供技术支撑。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株 植物病原菌禾谷镰刀菌(Fusariumgraminearum)标准菌株PH1、生防真菌寡雄腐霉(Pythiumoligandrum)PO-1及瓜果腐霉(Pythiumaphanidermatum)PA-1保藏于江苏省农业科学院农产品产地环境研究中心。PH1与2种腐霉菌分别在PDA和V8培养基上保存,使用前 20 ℃ 活化2次。
1.1.2 培养基 PDA培养基:200 g马铃薯煮沸 20 min 后用4层纱布滤出浸提液,加入20 g葡萄糖、15 g琼脂粉,蒸馏水定容至1 L即为PDA液体培养基;YEPD培养基:10 g蛋白胨、3 g酵母提取物、20 g葡萄糖,蒸馏水定容至1 L;绿豆汤培养基:20 g绿豆煮沸30 min后用3层纱布滤出浸提液,蒸馏水定容至1 L;V8培养基:100 mL V8果汁中加入1 g CaCl2,7 000 r/min离心5 min收集上清液,蒸馏水定容至1 L即为V8液体培养基,再加入15 g琼脂粉即为V8固体培养基。以上培养基均在121 ℃高压灭菌15 min。
1.1.3 供试植物 小麦种子为市场购买的感病品种,4 ℃保存备用。试验于2021年在江苏省农业科学院开展。
1.2 P. oligandrum对F. graminearum菌丝生长的影响
平板对峙试验:从平板边缘划取相同大小的新鲜菌饼,在距PDA平板边缘约2 cm的相对应的2点上分别接种禾谷镰刀菌PH1菌饼和寡雄腐霉 PO-1 菌饼。以接种PH1和瓜果腐霉PA-1的处理为阳性对照,以接种PH1和PDA菌碟的处理为阴性对照。置于25 ℃培养箱中对峙培养2、4、6 d后分别观察菌落形态,使用Image J软件测量PH1菌落面积。
1.3 P. oligandrum卵孢子液对F. graminearum孢子萌发的影响
PH1分生孢子悬浮液的制备:菌株在PDA上培养2~3 d,从新鲜培养的PH1平板边缘划取5~10个菌碟接种于绿豆汤培养液,25 ℃、170 r/min摇培 5~7 d,用无菌擦镜纸过滤,滤液经5 000 r/min离心10 min,弃上清,用灭菌水洗涤2次,显微镜下调整孢子悬浮液至适宜浓度。
PO-1卵孢子悬浮液的制备:在直径为90 mm的圆形培养皿中倒入30 mL V8液体培养基,从新鲜的PO-1平板边缘割取直径为7.5 mm的菌丝块倒置在培养基液面上,20 ℃避光培养,5 d后取出菌毯并弃置固体培养基部位,加入无菌水用小型豆浆机匀浆,显微镜下调整PO-1卵孢子悬浮液至适宜浓度。
在50 mL锥形瓶中添加8 mL YEPD培养基和 1 mL 浓度为5×105个/mL的PH1孢子悬浮液,以无菌水为空白对照,加入1 mL浓度为5×105个/mL的PO-1卵孢子悬浮液。在25 ℃、180 r/min恒温摇床中培养8 h后观察孢子萌发率,并使用Image J软件测量萌发芽管长度(计算每个孢子最长芽管的长度),每个处理最少测量150个孢子。
1.4 P. oligandrum卵孢子液对F. graminearum活体侵染的影响
小麦幼苗培养:小麦种子采用3%次氯酸钠溶液浸泡5 min,无菌水冲洗干净后,采用75%乙醇溶液浸泡5 min,再次用无菌水冲洗干净。将完成灭菌的小麦种子放置于干净的烧杯中,加入无菌水浸泡。24 h后挑选露白的种子转移至铺设8层纱布和1层滤纸的培养皿中,加入无菌水至滤纸微微湿润后避光催芽36~48 h,期间用喷壶保持皿内水分。
侵染与拮抗:挑选生长情况相对一致的幼苗转移至新的放有8层纱布和1层滤纸的培养皿中,在 25 ℃ 养苗室内12 h—12 h光暗交替培养。待叶尖稍露出胚芽鞘1~2 mm,剪去顶端3 mm制造伤口。在伤口处滴加2 μL浓度为1×105个/mL的PH1孢子悬浮液,2 h后喷洒10 mL浓度为1×105个/mL的PO-1卵孢子悬浮液,以喷洒10 mL无菌水为对照。后续每天喷洒2次,每次5 mL,7 d后观察小麦发病情况并记录病斑长度。每个处理设置3个平行,每个平行为25株小麦,试验重复2次。
1.5 P. oligandrum产孢诱导剂的筛选
初筛:在V8培养基中分别添加硫酸镁、硫酸铜、硫酸锌、碘化钾、氯化钴、钼酸钠、硫酸铵和乙二胺二乙酸铁钠(EDTA铁钠)作为诱导剂,根据前期预试验结果分别设置质量分数梯度:硫酸镁、硫酸铜、硫酸锌、碘化钾、氯化钴、钼酸钠设置6组质量分数梯度,分别为5、10、50、100、150、200 mg/L;硫酸铵设置6组质量分数梯度分别为10、100、200、300、400、500 mg/L;EDTA铁钠设置6组质量分数梯度分别为100、500、1 000、1 500、2 000、2 500 mg/L。与“1.3”节中的方法相同,20 ℃避光培养5 d后取出菌毯并弃置固体培养基部位,加入30 mL无菌水用小型豆浆机匀浆,显微镜下统计PO-1卵孢子数量。
正交试验:在初筛试验的基础上,挑选对PO-1产孢具有促进效果的诱导剂与其合适的梯度范围,在梯度范围内选取合适的水平进行正交试验,并根据最优组合对试验结果进行验证。
1.6 数据处理与分析
试验数据利用SPSS Statistics 26软件进行方差分析(α=0.05),计算所有均值的标准差。
2 结果与分析
2.1 P. oligandrum对F. graminearum菌丝扩展的离体抑制效果
由图1可知,通过对峙平板试验发现,与对照及PA-1处理相比,PO-1能够显著抑制病原菌PH1的菌丝扩展。4 d时PO-1菌丝和PA-1菌丝分别与PH1菌丝接触,此后PO-1明显抑制了PH1菌丝生长,其扩展速率几乎停止,边缘接触位置的菌丝呈现出不正常的黄色,而PA-1则无明显抑制作用,PH1菌丝覆盖PA-1并继续生长,边缘位置菌丝生长情况与对照组无明显差异(图1-a)。根据菌丝面积图像采集结果,2 d后PO-1处理的病原菌菌丝扩展非常缓慢,4 d和6 d时菌落面积仅为空白对照的60.13%和53.60%,而PA-1处理的病原菌菌落面积与空白对照相当(图1-b)。

2.2 P. oligandrum卵孢子对F. graminearum孢子萌发的抑制效果
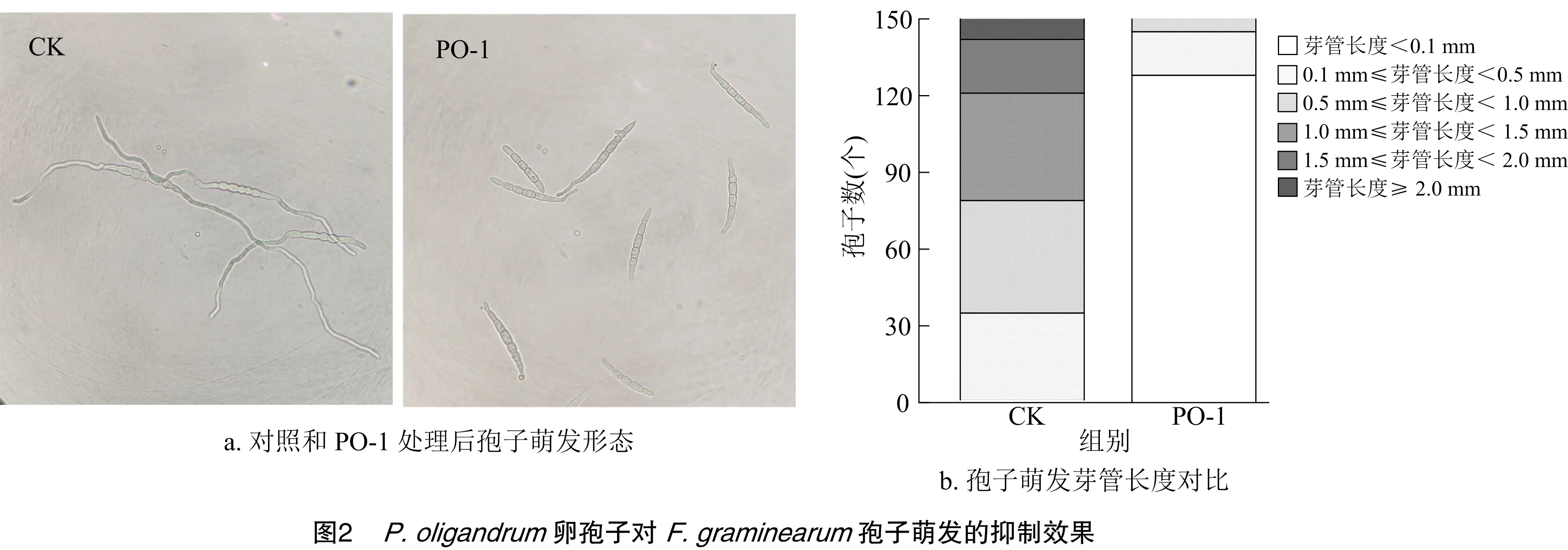
分生孢子萌发是F.graminearum侵染致病的重要方式。由图2-a可知,正常芽管一般从两端长出,长度达孢子长度的3~5倍,而PO-1卵孢子悬浮液处理的PH1分生孢子芽管发育不正常,萌发的位置随机分布。通过显微镜观察测定,由图2-b可知,自然萌发的PH1孢子8 h后萌发率为99.9%,平均芽管长度为178.12 μm,而PO-1卵孢子悬浮液显著抑制PH1孢子的萌发和芽管伸长(P<0.05),孢子萌发率仅为14.6%,平均芽管长度为 6.59 μm;表明P.oligandrum卵孢子对病原菌无性生殖能力具有显著抑制作用。
2.3 P. oligandrum卵孢子对F. graminearum侵染小麦的活体抑制效果
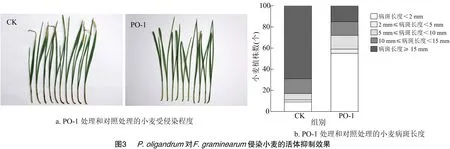
通过活体接种的方式研究P.oligandrum对F.graminearum侵染小麦的防治效果。由图3可知,在小麦伤口处接种PH1孢子悬浮液后,小麦茎基部出现黑斑,叶部尖端发黄枯萎,植物组织逐渐坏死,而喷洒PO-1卵孢子悬浮液后,小麦茎基部不发病或发病减缓,叶部尖端健康未枯萎;7 d后PO-1处理的茎基部被完全侵染的小麦植株(病斑长度≥15 mm)占14%,显著低于对照组(68%),茎基部几乎未发病的小麦植株(病斑长度<2 mm)占57%,显著优于对照组(8%)。
2.4 P. oligandrum产孢诱导剂配方的筛选
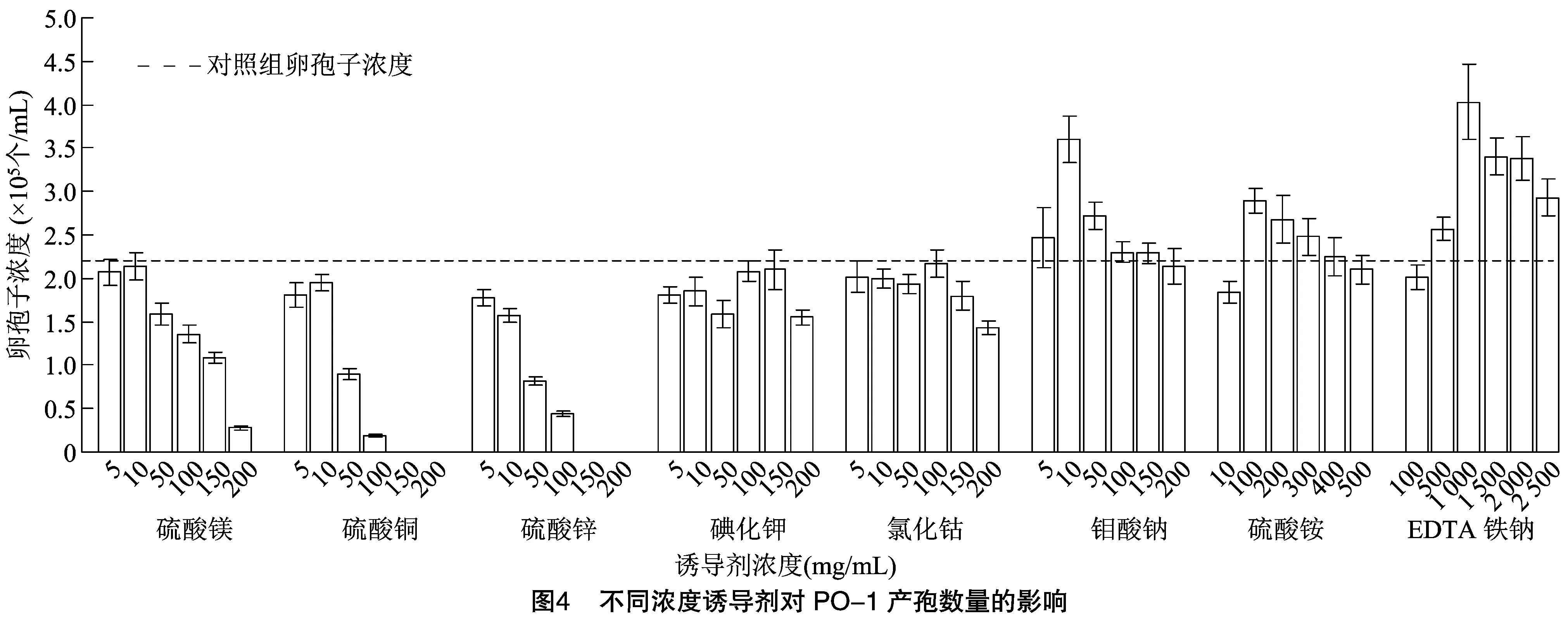
由图4可知,不添加诱导剂的对照组5 d内产孢数量为2.20×105个/mL,不同浓度的硫酸镁、硫酸铜、硫酸锌、碘化钾和氯化钴均无法促进PO-1产孢,而5~150 mg/L 的钼酸钠、100~400 mg/L的硫酸铵、500~2 500 mg/L的EDTA铁钠均能不同程度地提高PO-1的产孢数量。
以钼酸钠、硫酸铵、EDTA铁钠3个诱导剂作为研究变量,在所得梯度浓度范围内分别选取4个水平进行正交试验,采用L16(43)正交试验确定PO-1产孢诱导剂的最优配方,所用正交试验因素水平见表1,试验结果见表2。

表1 PO-1产孢诱导剂筛选正交试验因素水平
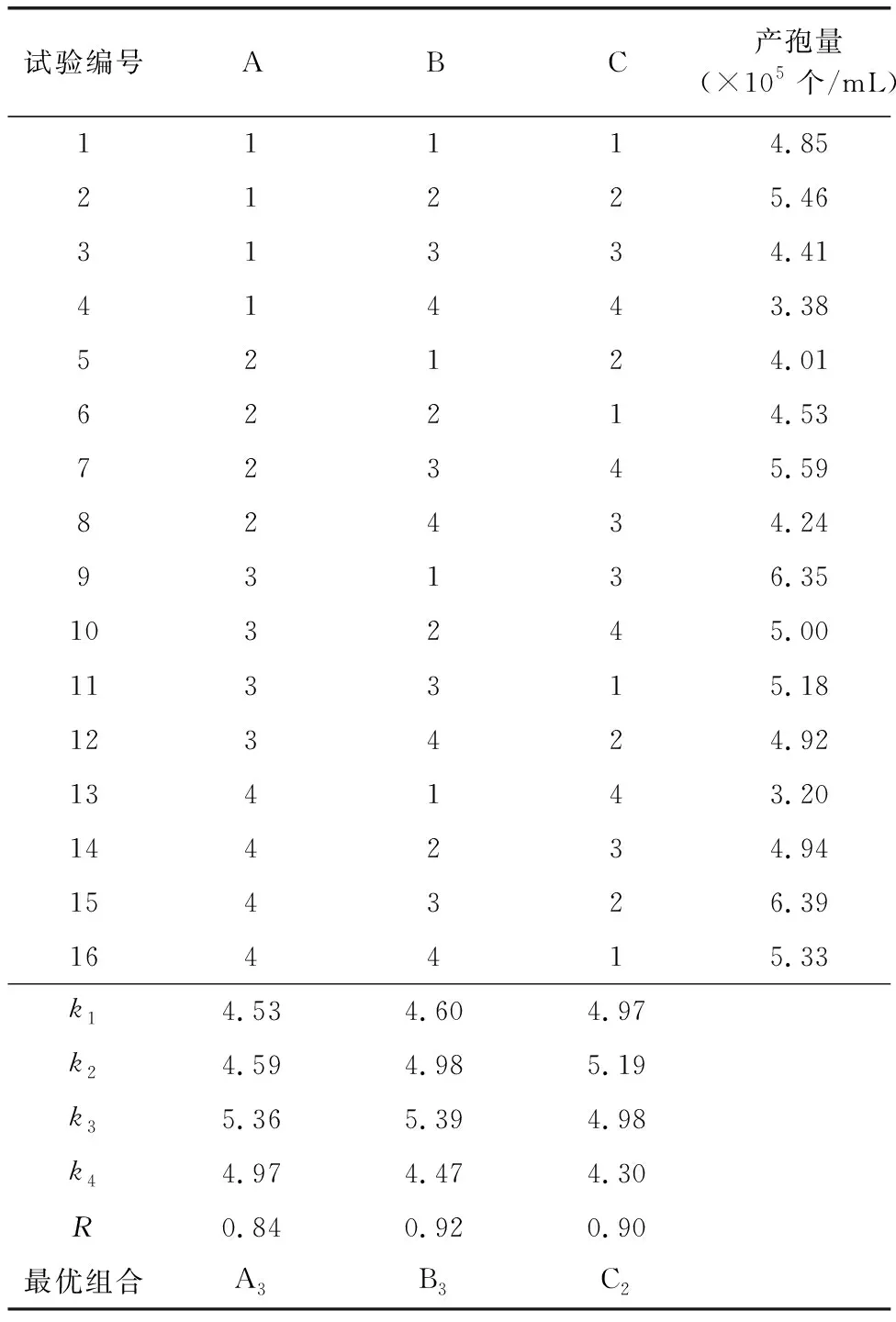
表2 产孢诱导剂筛选正交试验结果
由表2可知,3个诱导剂对PO-1产孢数量的影响能力顺序为 B>C>A,最优组合为A3B3C2,即硫酸铵对产孢数量的影响最大,EDTA铁钠次之, 钼酸钠对产孢的影响最小,当钼酸铵、硫酸铵、EDTA铁钠浓度分别为30、150、1 000 mg/L时,PO-1产孢数量最多。由于正交试验的组合中不包含最优组合,故按照最优组合的参数进行验证试验,结果表明该组合所得卵孢子数为7.04×105个/mL,是对照组产孢量的3倍以上。
3 讨论
研究表明,生防真菌的抑菌机制以重寄生作用和分泌抗生物质为主。Zhang等从海洋环境中分离得到1株木霉菌HN082102.1,能够包围并分解尖孢镰刀菌菌丝从而抑制黄瓜枯萎病[16];Yassin等在离体试验中发现,木霉菌属能够通过分泌多种细胞壁分解酶来分解真菌细胞壁,对小麦黑点病的多种致病菌均有抑制作用[17];Ferraz等发现,酿酒酵母和异常威克汉姆酵母能够通过死体营养寄生的方式控制可可植株的主要致病菌Moniliophthoraperniciosa,其中,异常威克汉姆酵母可通过菌毛相互连接形成伞状网络,从而集体杀死病原菌[18];Horner等采用绿色荧光蛋白(GFP)标记寡雄腐霉,发现寡雄腐霉菌丝可围绕致病疫霉,并将其作为营养来源[19]。本研究通过离体对峙试验发现,禾谷镰刀菌PH1与寡雄腐霉PO-1的菌丝接触后,PH1菌丝生长明显受阻,形态发生变化,直接接触部位菌丝呈黄色。瓜果霉腐PA-1与PO-1同属,两者所需营养与空间相似,但PA-1与PH1菌丝接触后不能阻碍其生长,也未使其菌丝形态发生变化,菌落形态与对照组一致。这与Ribeiro等的研究结果[20]相似,他们发现寡雄腐霉能够抑制P.periplocum侵染甜菜根,而对同属的棘腐霉(P.acanthicum)则不具有这种抑制作用。
禾谷镰刀菌引起的小麦赤霉病是全球谷物生产面临的关键问题之一,在全球范围内主要种植区均有报道[21]。大量研究证明,生物防治手段对禾谷镰刀菌具有抑制作用。Chen等从小麦穗部微生物菌群中获得1株高效生防细菌PseudomonaspisciumZJU60,它通过大量分泌抑菌活性物质(吩嗪-1-甲酰胺)抑制赤霉病菌生长、致病和毒素合成[22]。Xu等分离出1株解淀粉芽孢杆菌,能够抑制禾谷镰刀菌并降解其产生的玉米赤霉烯酮(ZEN)毒素[23];Diabankana等发现,芽孢杆菌PS17能够在对峙试验中抑制包括禾谷镰刀菌在内的多种镰刀菌属病原菌的生长[24]。目前,针对禾谷镰刀菌的生物防治菌株以生防细菌为主,对生防真菌的研究较少。
分生孢子是高等真菌繁殖和病原菌传播的主要方式和关键途径,不但具有长期生存能力,还能在逆境中保护自身基因组免受伤害,孢子萌发是病原菌突破防线侵染植株的第一步[25];Ngolong等认为孢子会广泛分布在环境中,一旦感应到外界条件适宜就会开始萌发并侵染植物[26]。抑制病原菌孢子萌发是防治植物病害的有效举措之一,张莹莹等筛选出1株多黏类芽孢杆菌P1,可抑制71.3%的芸薹根肿菌孢子萌发,从而防治菜心根肿病[27];Liu等发现,采用2 mg/L臭氧处理硫色镰刀菌(F.sulphureum) 2 min 可损伤其菌丝和孢子结构,从而将马铃薯干腐病病斑直径降低27.20%、蛇形菌素含量降低39.44%[28]。在离体试验中,本研究发现PO-1能够抑制PH1孢子萌发,阻碍其芽管伸长,采用PO-1处理PH1侵染过的小麦植株,可显著降低发病率,减缓病斑扩展,提示PO-1可作为防治小麦赤霉病病害流行的有效手段。
孢子是生防真菌抑制病原菌控制病害的主要功能成分,但与细菌相比,生防真菌孢子产量较少,难以满足大规模生产的要求,探究如何提高真菌孢子产量,是推动生防真菌实际应用的必要保障。研究人员常通过改变培养基中碳氮等营养物质、调节铵离子浓度或加入其他人工添加剂等方式来提高孢子产量[14-15]。本试验对几种常见的产孢诱导剂进行了初步筛选,发现钼酸钠、硫酸铵和EDTA铁钠对PO-1产孢数量具有明显影响,在V8培养基中加入30 mg/L钼酸钠、150 mg/L硫酸铵及 1 000 mg/L EDTA铁钠,20 ℃培养5 d后,产孢数量为7.04×105个/mL,是对照组产孢量的3倍以上,这为提高寡雄腐霉生防制剂的生产效率提供了重要的技术支撑。
