糖尿病与宿主肠道菌群的关系及饮食介导的菌群调控作用研究进展
2023-09-12王丹萍徐珒昭张晓航徐远志解庆刚许晓曦
王丹萍,徐珒昭,张晓航,宁 可,刘 叶,徐远志,解庆刚,许晓曦,*
(1.东北农业大学食品学院,黑龙江 哈尔滨 150030;2.东北农业大学 乳品科学教育部重点实验室,黑龙江 哈尔滨 150030;3.黑龙江飞鹤乳业有限公司,黑龙江 齐齐哈尔 164800)
糖尿病是胰岛素分泌不足(和/或利用障碍)引发的代谢性疾病,典型的糖尿病临床表现为“三多一少”症状,即多饮、多尿、多食、体质量下降,以及高血糖、高尿糖症状[1-2]。近年来,随着生活节奏的加快、超加工食品的泛滥及生活、工作压力的日益加剧,糖尿病发病人数激增且呈现年轻化趋势。国际糖尿病联盟在2021年调查数据显示,全球糖尿病患者人数已达5.37 亿,其中中国患病人数约为1.36 亿,较2020年增长了2.8%[3]。糖尿病患者的快速增长给各个国家均带来巨大的经济负担,2007—2017年美国用于糖尿病的支出从2 320亿 美元增加到7 270亿 美元,且预计到2045年突破7 600亿 美元[4]。
饮食在生命体过程中发挥着至关重要的作用。近期有研究指出[5],与物种的体型和寿命相比,引起动物死亡风险的更关键因素是饮食。糖尿病的发生、发展与饮食结构和方式密切相关,高脂高糖饮食被机体代谢后会促进肠道致病菌生长,产生毒力因子脂多糖(lipopolysaccharide,LPS)、氧化三甲胺(trimethylamineN-oxide,TMAO)以及吲哚等,也可使血浆中游离脂肪酸升高,损伤胰岛素分泌功能,进而导致肥胖及糖尿病[6-10]。肠道微生物作为吸收饮食成分产生相应代谢物的转换载体,其组成结构的动态平衡与糖尿病的发生、发展密切相关,一旦失衡可直接导致糖、脂、氨基酸代谢异常,出现非感染性炎症,肠屏障通透性增加,使肠道致病菌产生的毒素直接入血。近年来,一些研究结果表明肠道菌群是饮食与疾病之间的主要介导因素。科学饮食可增加菌群的多样性、建立良好的肠道环境并加固肠道屏障,激发机体自身保护机制降低糖尿病发生率及其恶化;与其相反的是,不良饮食(高脂、高糖、高盐、少膳食纤维等)会促进有害菌增殖,破坏肠稳态及屏障作用,降低肠道免疫功能。已有不少临床研究结果证实,不同类型糖尿病的肠道菌群结构特征以及与其他检测指标具有直接的相关性。归纳、整理糖尿病患者的肠道微生态特征,充分阐述饮食介导肠道菌群及其代谢产物通过调节肠免疫达到预防和调控糖尿病的原理及方法至关重要,同时可以指导患者通过科学饮食有效缓解并延缓糖尿病并发症,也为糖尿病的及早发现和干预指明方向。
1 肠道微生物与糖尿病的关系
糖尿病患者往往会出现肠道微生物群系稳态失衡、屏障受损、免疫功能下降等现象,严重者会诱发如中风、冠状动脉阻塞、视网膜病变,以及肝脏、肾脏等组织器官功能减退、衰竭等并发症[11-13]。是否能够通过其肠稳态改变及一些菌群代谢产物的变化进行糖尿病的预判还需深入研究。
1.1 不同类型糖尿病及其患病个体肠道菌群特征
不同类型糖尿病患者的基因、生活环境、饮食习惯各异,致使其肠道微生物特征也存在明显差异。肠道微生物标志性特征可作为糖尿病诊断的表征之一。
1.1.1 1型糖尿病
肠道微生物组与糖尿病在病理生理学方面有着密切联系,菌群结构的改变可以影响1型糖尿病(type 1 diabetes,T1D)患者的糖代谢与脂肪代谢。T1D主要由遗传因素引起,是由易感个体细胞免疫介导的胰腺细胞自我破坏引起胰岛素缺乏导致的自身免疫性疾病。大部分T1D患者是先天性,即从出生开始肠道有益菌就低于正常水平,其肠道微生物群的短链脂肪酸(short-chain fatty acids,SCFAs)生物合成、支链氨基酸分解代谢以及细菌发酵能力相对较弱[14]。T1D患者肠道菌群特征表现出α多样性显著降低,粪杆菌属(Faecalibacterium)、考拉杆菌属(Phascolarctobacterium)、芽孢杆菌属(Bacillus)、双歧杆菌属(Bifidobacterium)等相对丰度降低,拟杆菌属(Bacteroides)、布劳特氏菌属(Blautia)、链球菌属(Streptococcusgenera)等相对丰度显著升高[15],瘤胃球菌(Ruminococcus)、韦荣球菌属(Veillonella)等在不同文献中结果存在差异性,普雷沃菌属(Prevotella copri)、惰性真杆菌及Alistipes inops富集。在门水平上T1D患者和健康人群也存在差异,两者优势菌门分别是拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)[16]。T1D患者肠道内细菌代谢活性减弱,相应的外分泌胰腺蛋白质水平较低,在T1D患者体内发现其血糖控制性指标糖化血红蛋白水平与普雷沃氏菌科丰度呈负相关,肠杆菌目丰度与平均血糖指数呈正相关[17]。
1.1.2 2型糖尿病
2型糖尿病(diabetes mellitus type 2,T2DM)患者类型占总糖尿病患者的90%,目前已发现瘤胃球菌丰度与T2DM呈正相关,而产生丁酸盐细菌(如毛螺菌属、布劳特氏菌属)的丰度与T2DM呈负相关[18]。肠道内拟杆菌属(Bacteroides)和双歧杆菌属(Bifidobacterium)是糖尿病研究中最常见的有益菌属,较多研究结果表明双歧杆菌在糖尿病患者体内比例显著减少[19-20],拟杆菌属丰度则与T2DM无明显相关性。阿克曼菌属(Akkermansia)作为一种新发现的对宿主代谢有益的共生菌群成员,其丰度与T2DM呈显著负相关,也因此被作为糖尿病患者肠道标志菌[21-22]。小鼠肠道菌群失调,如疣微菌在门、纲、目等水平增加,拟杆菌在科、属、种水平减少,普氏菌在科、属水平减少会促进其T2DM的发展[23]。研究进一步发现,一些肠道菌群超出相对稳定的阈值范围会增加血清LPS、连蛋白水平,使肠道屏障功能受损,促进T2DM的恶化[24-25],而在对T2DM小鼠进行粪菌移植后,其肠道屏障功能增强,血清LPS、LPS结合蛋白和促炎细胞因子水平均呈降低趋势。
1.1.3 妊娠糖尿病
妊娠期孕妇生理变化明显,是糖尿病易感人群,诱发妊娠糖尿病(gestational diabetes mellitus,GDM)内部原因是遗传因素,其决定个体易感性,外部原因则是饮食不规律引起机体炎症,GDM是一种异质性代谢紊乱疾病,患者容易发生高血压和产后出血等隐患,对后代健康也会有一定影响[26]。糖尿病易致使肠道稳态发生较大变化,大多数患者GDM肠道菌群α多样性降低,放线菌门(Actinobacteria)、变形菌门(Proteobacteria)丰度呈上升、拟杆菌门丰度普遍呈下降趋势。水痘克雷伯菌、产碱杆菌属(Alcaligenes)等丰度增加,而甲烷短杆菌(Methanobacilerium)、另枝菌属(Alistipes)丰度相对降低[27-28]。其中条件致病菌产碱杆菌属作为潜在的病原体可能会诱发肺炎、腹膜炎甚至结直肠癌等,不同时期GDM患者肠道菌群变化差异很大,但是仍能看出GDM的发生与一些菌群存在相关性。
1.1.4 其他类型糖尿病
某些糖尿病的症状介于T1D和T2DM之间且区别于两者,包括莫迪型糖尿病(Mody diabetes,MODY2)、成人隐匿自身免疫性糖尿病(latent autoimmune diabetes in adults,LADA),以及胰腺外分泌疾病、矮妖综合征A型胰岛素抵抗、药物引起的皮质激素异常、感染青少年一型、免疫介导性一型等糖尿病。研究发现,MODY2患者肠道内普雷沃菌丰度显著升高,瘤胃球菌和拟杆菌丰度显著降低[12]。LADA患者则严重缺乏SCFAs产生细菌,肠道微生物群结构与T1D患者更相似[29]。
1.2 微生物对糖尿病的调控作用
微生物群代谢产物及衍生物通过肠道上皮细胞进入机体,直接或间接影响炎症调节、肠道通透性变化以及糖脂代谢而发挥重要调控作用,调控路径汇总如图1所示。肠道通透性决定了某些菌群代谢产物通过特定分子通道而被吸收。例如,肠道菌群代谢的SCFAs等刺激小肠L细胞释放胰高血糖素样肽-1(glucagon-like peptide-1,GLP-1)和肽YY,影响胰岛素合成、饱腹感和脂肪组织的活跃程度。同样,黑皮质素样肽通过黑皮质素-1受体发挥作用,抑制细胞因子释放以响应促炎刺激反应[30-32]。三甲胺、LPS、咪唑丙酸、吲哚等促进促炎细胞因子、趋化因子释放,引发炎症。脂肪细胞是脂肪酸代谢和内分泌器官,非酯化脂肪酸释放的增加抑制了骨骼肌中胰岛素刺激的葡萄糖代谢,从而引发胰岛素敏感性以及T2DM[33-34]。
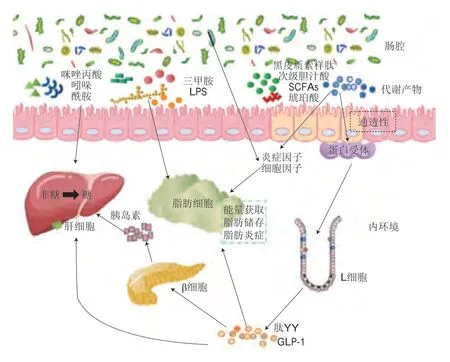
图1 微生物菌群参与T2DM调控的路径Fig.1 Pathways by which the intestinal microbiota is involved in diabetes mellitus type 2 (T2DM) regulation
1.2.1 参与脂肪代谢
肠道微生物参与机体的脂肪代谢过程,结肠中微生物可将未完全消化的碳水化合物转变为S C FA s。S C FA s 中丁酸的益菌效应尤为突出,产丁酸菌主要包括梭菌科(Clostridiaceae)、消化链球菌科(Peptostreptococcaceae)、巴氏杆菌科(Pasteurellaceae)等[35]。普雷沃菌、双歧杆菌、粪杆菌属、罗斯氏菌属等[36]也可以增加肠道中SCFAs水平。SCFAs可以通过激活G蛋白偶联受体(G protein-coupled receptor,GPR)43和GPR109A[37],降低炎症水平[38]并作为脂质和葡萄糖的合成原料[39]而达到预防和稳定糖尿病的目的[40]。空气污染及不良饮食对机体健康造成的不利影响体现在肠道菌群失稳,产黑普氏菌(Prevotella melaninogenica)等产生的LPS可抑制小鼠微粒体甘油三酯转移蛋白的表达[41],使脂代谢异常及SCFAs含量降低,同时肠道益生菌的减少也会影响机体对脂类的代谢,如介导炎症因子、脂肪因子和IgA免疫应答发生变化,诱导肥胖、胰岛素抵抗和糖尿病的发生[42-44]。膳食对肠道菌群结构产生一定影响,而肠道菌群变化也影响宿主对食物及能量的摄取,乳杆菌属、双歧杆菌属、链球菌属等益生菌可显著降低糖尿病患者的空腹血糖、甘油三酯及胰岛素水平[45-46]。此外,阿克曼菌属可增加脂肪组织中2-油酰基甘油、2-棕榈酰甘油、2-酰基甘油的水平,从而促进脂肪酸氧化和脂肪细胞分化[47]。多种益生菌尤其是产丁酸菌会参与脂肪代谢过程,而通过膳食增加产丁酸菌数量可有效改善肠道微生态紊乱,调控炎症,缓解糖尿病。
1.2.2 调控炎症因子
糖尿病可诱发机体促炎细胞因子、趋化因子和炎症蛋白水平升高。LPS可促进抗炎细胞因子和趋化因子表达引起代谢性内毒素血症和低级别炎症,肠道菌群衍生物琥珀酸也可促进解偶联蛋白1表达,增加脂肪能量消耗[48-49];肠黏膜发生炎症和肠屏障损伤时,菌群产生的内毒素、细菌成分和代谢物易位进入循环系统,削弱肝脏清除力,促进代谢炎症发生[50];白细胞介素(interleukin,IL)-22是能恢复胰岛素敏感性和缓解糖尿病的抗炎细胞因子,研究证实T1D模型小鼠中肠黏膜受损,肠道屏障功能改变,诱导IL-22表达的分节丝状菌丰度减少,使IL-22表达显著降低[51-52]。目前的研究证明,肠道益生菌群的摄入可以降低人体炎症,如罗氏菌属(Roseburia)、拟杆菌、阿克曼菌以及乳酸菌属(Lactobacillus)等微生物诱导IL-10过度表达并抑制肿瘤坏死因子α表达从而降低胰岛素抵抗[53-55]。同样,多种乳酸菌可降低IL-1β、单核细胞趋化蛋白-1、细胞间黏附分子-1、IL-8、CD36蛋白和C反应蛋白等炎症蛋白水平表达[56-57]。罗氏菌属和普拉梭菌均为产丁酸菌,通过抑制核因子κB的活性而降低炎症[58-59]。干酪乳杆菌(Lactobacillus casei)会降低促炎细胞因子II型干扰素的产生[60]。由此表明,肠道菌群的调节可从抵抗炎症、维持肠道菌群稳态以及恢复肠黏膜功能方面显著改善糖尿病。
1.2.3 改变肠道通透性
肠道屏障通透性增加是T2DM的一个特征,肠道通透性增加导致肠道微生物代谢物易位进入血液容易引起代谢性内毒素血症[61]。糖尿病患者肠黏膜屏障功能破坏以及血糖升高导致胞间连接完整性降低,使肠道上皮细胞渗透性增加,影响抗原的吸收,最终诱发全身病理性免疫反应[62-63]。以往研究证明了肠道微生物对肠道通透性的重要作用,拟杆菌属可上调结肠中紧密连接蛋白基因的表达,降低肠道通透性,从而减少LPS的产生,改善内毒素血症[64]。阿克曼菌可通过激活上皮中的腺苷酸活化蛋白激酶改善肠道紧密连接,该菌的外膜蛋白Amuc-1100可增强紧密连接蛋白的表达,还能抑制肠道中的大麻素1型受体表达,降低肠道通透性和全身LPS水平[65]。由普拉梭菌和罗氏菌属产生的丁酸盐可以通过血清素转运体降低肠道通透性。肠道微生物作为生物屏障的重要部分,可阻止糖尿病的恶化和并发症的产生。
1.2.4 影响肠促胰素效应
糖尿病患者肠促胰素效应受损,进餐后GLP-1浓度的升高幅度减小。作为机体代谢的关键调节因子,GLP-1可刺激胰岛素分泌[66],提高心率、保护心脏、降低中风风险,还可作用于大脑以抑制进食和保护神经[67],GLP-1受体激动剂可治疗T2DM和肥胖,Tirzepatide是一种包括GLP-1受体的双重激动剂,可显著降低糖化血红蛋白、空腹血糖浓度及体质量,并提升糖化血红蛋白达标率,且安全性良好[68]。肠道是GLP-1的主要分泌器官,胰岛和胃也可通过分泌GLP-1发挥调控血糖的作用[69],但远端肠道是调控GLP-1分泌的关键位点。碳水化合物、脂肪和蛋白质的摄入可刺激小肠L细胞分泌GLP-1,抑制胰高血糖素分泌、胃排空和食欲,增加饱腹感和参与心血管活动等[70]。除此之外,肠道菌群的代谢产物可影响GLP-1的产生,阿克曼菌产生的P9蛋白通过与细胞间黏附分子2结合而诱导GLP-1的分泌,从而改善小鼠的糖稳态及肥胖[71]。该菌还可通过降低β-鹅脱氧胆酸、促进胰岛素分泌和糖原合成从而抑制糖异生作用[72]。
1.3 基于肠道微生态介导的糖尿病治疗方法
迄今为止糖尿病仍无法根治,现有的主要治疗方法为饮食调整、加强运动、口服药物以及注射胰岛素。而一些关于利用益生菌干预(口服益生菌制剂及粪菌移植术)对糖尿病治疗的临床试验及动物实验研究都证明了基于微生态治疗糖尿病具有良好的效果。
1.3.1 糖尿病传统治疗方法
糖尿病传统治疗方法主要为药物治疗,二甲双胍具有明显的降糖效果,是治疗T2DM的重要药物,可显著降低糖尿病患者体质量和低密度脂蛋白胆固醇[73]。近年来,研究还发现其具有抗肿瘤等其他功效。此外,还有一些具有控糖或减少糖摄入的药物,如罗格列酮和阿卡波糖[74]、胰岛素注射类的诺和灵3R(笔芯)[75]等。这些药物可使糖尿病患者血糖水平恢复正常,并有效缩短尿糖达标时间,但摄入药物过多会引起皮质激素异常等新问题[76]。目前一些具有改善葡萄糖稳态等糖尿病症状的中草药成分被发现,如黄连素可抑制NOD样受体热蛋白结构域相关蛋白3的活化并减少促炎因子释放、降低胰岛素抵抗,姜黄素可维持肠道屏障功能,人参皂苷对改善胆碱能及抗氧化系统并降低血糖也具有积极作用[77-78]。与西药相比,中草药成分对人体的副作用相对较小,但是见效相对较慢且需要多重成分共同作用。此外,更多研究显示中药成分发挥作用与肠道菌群存在重要关联。
1.3.2 基于微生态治疗糖尿病方法
健康机体中各项成分均保持着动态平衡,人体内环境动态平衡的变化表现为疾病的发生发展,肠道菌群的动态变化对应机体系统组件的改变,因此通过肠道菌群预测、预防和调控疾病成为未来医学研究领域的新趋势。分析肠道菌群的高维分子数据(如转录组、表观基因组、糖组、微生物组)可以对发病系统组织成分进行描述和监测[79]。相比传统的按照直线因果关系原则治疗,用系统论的原理和方法解决医学问题效果更为有效。研究发现,与定期服用降血糖药物常规治疗方法相比,规范化治疗对糖尿病患者的疗效更显著[80]。人为的菌群干预,如粪菌移植、服用益生菌制剂或者使用益生元介导肠道菌群逐渐成为调控糖尿病的新方式。
粪菌移植是一种将健康人体粪便中的功能菌群移植到患者胃肠道内,实现肠道疾病治疗的新技术。将健康小鼠的肠道菌群转移到T2DM小鼠可以显著提高糖尿病小鼠的肠道菌群多样性以及降低受体小鼠的胰岛素敏感性[81],粪菌移植改变了糖尿病小鼠的肠道菌群结构,并改善糖脂代谢、增加胰岛面积和β细胞比例。目前,粪菌移植技术以其标准化、人性化特点在相关领域得到广泛的运用。粪菌移植可显著促进T2DM患者菌群的定植,且粪菌移植结合生活方式干预还可减少总胆固醇及低密度脂蛋白胆固醇水平,降低肝脏硬度[82-83],多酚结合粪菌移植也可产生降血脂作用,黄连中提取的植物生物碱小檗碱协同益生菌可以提高肝脏低密度脂蛋白摄取和脂肪褐变[84]。
补充益生菌、益生元或合生制剂或可改善糖尿病患者的肠道菌群失调、促进胰岛素分泌、减少胰岛素抵抗从而调控糖尿病进程[85-86]。双歧杆菌可通过与胆固醇结合、阻断宿主肠道吸收或改变宿主胆汁酸信号转导改善血脂异常,实现T2DM风险控制[87]。菊粉可通过改善T1D小鼠肠道菌群组成变化而恢复胰腺正常功能,并显著提高胰岛素敏感性,恢复机体本身对血糖的控制并减少食物和水分摄入[88]。磷脂酰肌醇3-激酶(phosphatidylinositol-3-kinase,PI3K)-蛋白激酶B(protein kinase B,AKT)-哺乳动物雷帕霉素靶蛋白信号通路是经典的胰岛素信号响应通路[89]。有研究发现,紫苏油可通过PI3K/AKT信号通路调控T2DM小鼠肠道菌群,恢复肠道菌群多样性并缓解胰岛素抵抗[90],药食同源性的紫苏油可能是一种潜在的治疗糖尿病的功能性食品。
2 饮食通过肠道微生物介导调控糖尿病
现代医学及营养学逐渐发现并验证了通过改善肠道菌群、靶向菌群调节、精准营养等手段可有效对抗糖尿病。科学的个性化饮食可预防及纠正肠道炎症引发的机体糖代谢紊乱、免疫力下降、长期高血糖状态、胰岛素抵抗等症状(图2)。

图2 饮食通过肠道菌群干预糖尿病Fig.2 Diet affects diabetes by regulating the intestinal microbiota
2.1 科学配伍饮食对糖尿病预防与调控
科学配伍饮食可以维持肠道微生物稳态、建立良好的肠道环境以及加固肠道屏障,利用机体自身保护机制降低糖尿病发生几率和发展。植物膳食中含有丰富的天然功能性化合物及膳食纤维,有利于促进肠道益生菌群的定植并发挥其功效,影响菌群结构并提高SCFAs含量从而驱动机体的免疫代谢。合理的进食时间也可通过肠道微生态的改变发挥预防和改善T2DM血糖调控的作用[91-92]。
2.1.1 素食饮食和地中海饮食
以往素食饮食作为宗教传统以及可缓解肥胖、改善代谢被大众所接受和认知。其中植物性食物成分如VC、类胡萝卜素及其复合生物标志物可以降低血糖和尿糖排泄量、血浆总胆固醇、甘油三酯和极低密度脂蛋白胆固醇浓度等[93-94],且摄入量与T2DM发病率呈负相关[95-96],可见素食饮食可以降低疾病风险,是对糖尿病患者适用性较高的一种饮食模式。研究发现,素食饮食对肠道微生物的作用效果同样明显。植物性食物中多酚、多糖、聚糖等成分具有肠道益生作用。多酚包括黄酮和酚酸,黄酮类化合物具有广泛的生物学作用,如调节酶活性、抑制细胞增殖、抗氧化和抗炎特性。素食饮食干预显著改变肠道菌群的组成,增加碳水化合物降解菌,如双歧杆菌属、乳杆菌属[97],改变SCFAs水平[98]。目前研究较多的DASH(dietary approaches to stop hypertension)饮食和地中海饮食均属于半素食饮食,DASH饮食可显著降低代谢综合征患者收缩压、降压药需求及身体质量指数,并改变肠道菌群组成及功能,如影响SCFAs产生等。大量研究结果显示,长期坚持DASH饮食对降低炎症高血压[99]、糖尿病的降压[100]、降糖[101]效果明显。研究发现,地中海饮食干预可显著改善患者的血糖、胰岛素水平及胰岛素抵抗,并诱导毛螺菌科及瘤胃球菌科的富集,通过影响胆汁酸代谢而调控胰岛素水平[102-103],同时具有调节新陈代谢、降低体质量、提高运动能力的效果[104]。研究表明,地中海饮食的老年人步行速度更快、膝关节肌肉力量更大、整体认知水平更高以及记忆力更好,且与低脂饮食相比,地中海饮食老年人的颈动脉内膜中层厚度和颈动脉斑块高度降低更多[105-106],说明地中海饮食改善动脉粥样硬化效果更明显。因此对于糖尿病患者,运动结合地中海饮食是相对较好的选择。
与治疗糖尿病常采用的生酮饮食相比,素食饮食的可持续性更好[107],生酮饮食干预使患者缺乏纤维、VC、叶酸及镁等多种营养元素。因此生酮饮食方法不利于心脑血管疾病患者。
2.1.2 发酵食物或益生菌干预
近年来益生菌产业爆炸式发展,且很多发酵类食品产业也呈快速增长趋势。以往诸多研究已证实,某些益生菌可显著降低患者的糖化血红蛋白、空腹血糖和空腹胰岛素的水平以及胰岛素抵抗指数,且多菌株和高剂量的益生菌补充剂对血糖稳态更有益[108]。而GDM患者补充含有嗜酸乳杆菌、干酪乳杆菌、两歧双歧杆菌、发酵乳杆菌的益生菌产品可改善其过氧化物酶体增殖物激活受体γ、转化生长因子β和血管内皮生长因子的表达,以及血糖控制、血脂状态、炎症标志物和氧化应激[109]。研究表明,T2DM患者服用含有菊粉和产丁酸细菌拜氏梭菌(Clostridium beijerinckii)、丁酸梭菌(Clostridium butyricum)、厌氧丁酸菌(Anaerobutyricum hallii),复杂低聚糖降解细菌阿克曼菌、婴儿双歧杆菌的益生菌制剂可显著降低患者的餐后血糖、糖化血红蛋白及血糖增量,且安全性及耐受性良好[110]。另一项研究结果表明,含乳杆菌属、双歧杆菌属及链球菌属活菌株的益生菌补充剂可使T2DM患者的空腹血糖水平显著下降,高密度脂蛋白胆固醇水平显著上升[111]。对前驱糖尿病成年患者分别补充包含冻干的乳杆菌属及双歧杆菌属菌株的多菌种益生菌、基于菊粉的合生制剂,发现其可显著改善糖代谢相关参数,稳定血糖稳态。进一步研究发现,益生菌还可改善高密度脂蛋白胆固醇水平,但疗效存在一定的个体差异[112]。目前文献中关于益生菌的种类、剂量以及功效等级不一而同,但是基本认为乳酸菌和双歧杆菌是用于降低临床前和人群试验的空腹血糖和胰岛素水平最广泛应用的益生菌[113]。
2.1.3 间歇性禁食
除饮食结构和成分对糖尿病具有一定调节功效外,近期的研究发现禁食也可通过能量与糖分限制调控肠道微生态而改善机体代谢。间歇性禁食可引起细菌比例适应性变化,从而完成细胞和组织器官在进食和禁食期间的源物质在葡萄糖和脂肪酸/酮体间转换。常见的间歇性禁食模式有隔日禁食和限时进食。限时进食是通过调节昼夜节律系统影响体质量控制和生化参数,并通过影响瘦素分泌改善饱腹感调控系统;而隔日禁食在促进体质量控制激素调节的同时,还需要在禁食日减少能量摄入。二者均可显著减轻体质量、改善代谢并调节肠道菌群,通过改变厚壁菌门/拟杆菌门比例和增加有益菌属(如乳杆菌属和阿克曼菌)丰度而抑制体质量增长[114]。间歇性禁食是一种相对接受度更高的饮食干预措施,可实质性重塑肠道微生物组,产丁酸的毛螺菌科丰度明显上调[115]。改变能量摄入的时间对成人T2DM肥胖患者不仅具有降低体质量、血糖,改善胰岛素抵抗等作用,改善心血管方面的问题,如显著降低甘油三脂、总胆固醇、低密度脂蛋白含量[116-117]。目前又发现一种新型的间歇性禁食方法,即早限时进食(只在8:00—16:00期间进食),结果显示,与对照进食相比,早限时进食可改善全身胰岛素敏感性并增加餐后骨骼肌对葡萄糖和支链氨基酸营养素的摄取[118]。
2.2 不健康饮食影响肠道微生物诱发糖尿病
目前,西方饮食、快餐饮食(外卖)等不健康的饮食方式通过介导肠道微生物结构改变成为糖尿病发病的高风险诱因。高动物脂肪和蛋白质及低植物纤维饮食诱导宿主蛋白酶发生蛋白水解,生成含硫氨基酸和芳香族氨基酸,这些氨基酸与代谢性疾病有关的异常菌群发生硫酸盐还原反应、脱氨基反应、脱羧反应等多种复杂反应,生成硫化氢、氨、胺、氢气、二氧化碳、甲烷、有机酸、苯乙酸、对甲酚、苯丙酸、吲哚丙酸、LPS等代谢物,使肠上皮细胞黏液分泌减少,乙醛及肠腔内pH值增加,支链脂肪酸和TMAO等有害物质增加,引起血糖上升、胰岛素分泌下降。西式饮食使机体有益细菌(如约氏乳杆菌和格氏乳杆菌)减少和致病菌(如Romboutsia ilealis)增多,增加疾病风险。补充这两种乳杆菌可提高血液中还原型谷胱甘肽水平,改善肝脏的线粒体形态和功能,减少肝脏脂质、改善系统性糖代谢[119]。动物实验研究显示,某些肠道菌发酵膳食纤维产生的SCFAs(尤其是乙酸盐)使鼠柠檬酸杆菌的初期定植受到抑制,而鼠柠檬酸杆菌滞留感染促进了小鼠肠道低度炎症和胰岛素抵抗[120]。高脂饮食会引起小鼠肠道黏膜炎症,并导致上皮细胞线粒体活性下降,从而造成呼吸链电子受体(氧气和硝酸盐)的增加[121]。
现有研究已证实,饮食成分越单一,糖尿病患病概率越高,超加工食物的摄入会增加25%的T2DM患病风险[122]。肥胖者体内脂肪越多,其胰岛素抵抗和糖代谢异常更明显,T2DM风险越大。高脂肪食物的摄入会改变菌群结构,富含植物多糖的饮食与普雷沃氏菌的主导肠型相关,而高脂高糖饮食与拟杆菌属的主导肠型相关[123]。微生物群改变还会增加对食物过敏的易感性,有研究对小鼠进行12 周的高脂饮食喂饲诱导小鼠肥胖,证明过敏反应与肠道效应细胞反应失调、肠道通透性增加和肠道生态失调有关[124-125],肠乳酸杆菌的丰度与大鼠脂肪质量和体质量呈负相关,高脂饮食使肠乳酸杆菌大幅减少、致病菌丰度增加,其代谢产物导致内毒素血症发生并诱导产生炎症[126-128]。
食物的摄入种类、摄入量、摄入方式、摄入时间等均会引起机体包括肠道菌群稳态发生变化,不健康的生活方式(熬夜、吸烟、饮酒等)、精神状态(焦虑、抑郁等)及不良饮食(高脂、高糖、高盐、低膳食纤维、少新鲜绿叶蔬菜等)是糖尿病产生的主因。
3 结 语
以系统医学为代表的现代医学及营养学秉承了中国传统医学“食为药”的理念,将天然食物(或药食同源食材)作为预防及治疗疾病的一种特殊个性化方式,正逐渐被医护工作者和普通大众所认知和接受。可根据糖尿病患者的肠道微生态和健康人群之间的显著差异及早对糖尿病进行筛查,进而根据个体自身特点(菌群特征、遗传基因等),科学地、因人而异地制定健康饮食模式(如DASH饮食、间歇性饮食、地中海饮食和益生菌饮食),并且保持规律的生活方式更利于提高机体自愈能力,抵抗糖尿病风险,而采用粪菌移植或服用益生菌及益生元制剂可更为快速地调节肠道微生态,减少胰岛素抵抗并有效预防糖尿病并发症的发生。
