西藏色季拉山区风毛菊属植物花粉形态特征及其分类学意义
2023-09-09张淑钧杨欣欣
张淑钧 杨欣欣 罗 建*
(1.西藏农牧学院高原生态研究所,林芝 860000;2.西藏高原森林生态教育部重点实验室,林芝 860000;3.西藏林芝高山森林生态系统国家野外科学观测研究站,林芝 860000;4.西藏自治区高寒植被生态安全重点实验室,林芝 860000)
菊 科(Asteraceae)风 毛 菊 属(Saussurea)于1810 年 由 植 物 学 家De Candolle[1]建 立。1881 年Hooker[2]首 次 将 风 毛 菊 属 划 分 为2 个 亚 属。到1979 年前苏联学者Lipschitz[3]详细调查了全世界范围内的风毛菊类群,根据植株被毛、茎明显程度、苞片颜色、总苞片是否被附属物与膜质附属物是否明显等形态特征,将该属划分为6 个亚属20个组。该系统成为后期大部分风毛菊属分类系统的研究基础。随着对风毛菊属植物的深入研究,发现该属植物广泛分布于泛北极植物区,以亚洲分布物种最多,在青藏高原及周边地区尤为丰富,约有230 个地方性种[4-6],是风毛菊属的“现代分布中心”[7-9]。由于该属物种较多,分布广泛,其生境特殊而多样,既有高山和草地,也有荒漠与冰川等。不同生境下的物种,植株形态特征变异较大,表现出丰富的形态特征多样性[10-11]。此外,同域分布的一些物种为适应共处的极端环境,采取了相似的适应策略[12],导致种间界限模糊,如分布于雪山环境下的苞叶雪莲(S.obvallata)和宝璐雪莲(S.luae)等,均具有形态相似的膜质苞叶,为物种鉴定带来了困难[13-15]。
花粉形态特征具有稳定的遗传保守性,在植物分类及亲缘关系重建方面提供重要佐证[16-17]。花粉形态大小、萌发沟以及外壁表面纹饰等特征在种间差异显著,是物种划分的重要依据[18-20]。孙会忠等[21]对菊科绢蒿属(Seriphidium)10 种植物的花粉形态进行研究,结果表明10 种绢蒿属植物花粉粒较小,呈长球形、球形或近球形,赤道面观为圆形或椭圆形,极面观为三裂片圆形,认为绢蒿属和蒿属(Artemisia)有着较近的亲缘关系。王铁娟等[22]对6 种沙蒿花粉形态的比较研究,认为6 种沙蒿的花粉形态高度一致,极面观均为三裂圆形,赤道面观为圆形或椭圆形,具三孔沟,外壁纹饰在光镜下为表面颗粒状,在电镜下为排列较疏的刺,刺基外延。王一峰等[23]对青藏高原东缘28种风毛菊属植物花粉形态研究认为花粉形态的刺间表面纹饰、萌发沟的宽窄、刺的大小、分布密度和花粉大小在种间存在差异,组内花粉形态多样性丰富。目前,除王一峰等[23]外,对风毛菊属植物不同种间的孢粉学特征研究涉及仍然较少,不能准确地定义、分类,且缺乏对本属植物不同种类花粉形态的详细数据。
色季拉山区风毛菊属植物种类丰富,但目前对色季拉山区风毛菊属植物的研究、保护及利用仍然不足[24]。色季拉山地势南高北低高差起伏大,地形地貌复杂,山系纵横交错。印度洋季风暖湿气流和北方高原寒流在此交汇,形成独特的湿润半湿润气候,冬温夏凉,干湿季明显,形成了许多复杂的区域性小气候[25]。年降水量600~1 000 mm,年均温8~10 ℃,积温为3 000~3 500 ℃[26]。土壤类型丰富,同时由于高原隆升,海拔从低到高,依次受到热带、亚热带、温带和寒带气候的控制,孕育了该区极为丰富的植物资源[27-28]。
本文以色季拉山区丰富的风毛菊属植物种类为前提,调查和收集了22 种风毛菊属植物花粉和1 种须弥菊属(Himalaiella)植物花粉,将须弥菊属植物花粉与风毛菊属其他种一起观察,以了解其花粉微形态水平上是否同样存在分化。通过扫描电镜观察记录它们的花粉形态和大小、外壁纹饰特征,包括萌发孔沟、网眼、网脊、刺长和刺宽等特点,比较花粉形态在种间的差异,通过聚类分析,探讨其亲缘关系,拟解决以下问题:(1)观察23 种植物的花粉形态特征;(2)比较种间、组间和各亚属间花粉微形态的异同,探究其分类学意义。
1 材料与方法
1.1 研究材料
材料为采自色季拉山区22种风毛菊属植物和1 种须弥菊属植物的花粉(表1)。22 种植物分别来自3 个亚属:雪兔子亚属(Subgen.Eriocoryne)、雪莲亚属(Subgen.Amphilaena)、风毛菊亚属(Subgen.Saussurea)。其余1 种为白背须弥菊(Himalaiella auriculata),原放置在风毛菊属齿冠亚属(Subgen.Florovia),后来被修订为须弥菊属成员[13]。研究材料里,有13 个中国特有种,包括:拉萨雪兔子(S.kingii)、绵头雪兔子(S.laniceps)、硕首雪兔子(S.grandiceps)、多鞘雪莲(S.polycolea)、毛背雪莲(S.pubifolia)、尖叶风毛菊(S.ciliaris)、黑苞风毛菊(S.melanotricha)、薄苞风毛菊(S.leptolepis)、川滇风毛菊(S.wardii)、革苞风毛菊(S.coriacea)、弯齿风毛菊(S.przewalskii)、紫苞风毛菊(S.purpurascens)与粗裂风毛菊(S.grosseserrata),其中有4 个西藏特有种:拉萨雪兔子、硕首雪兔子、毛背雪莲和紫苞风毛菊[5-6]。
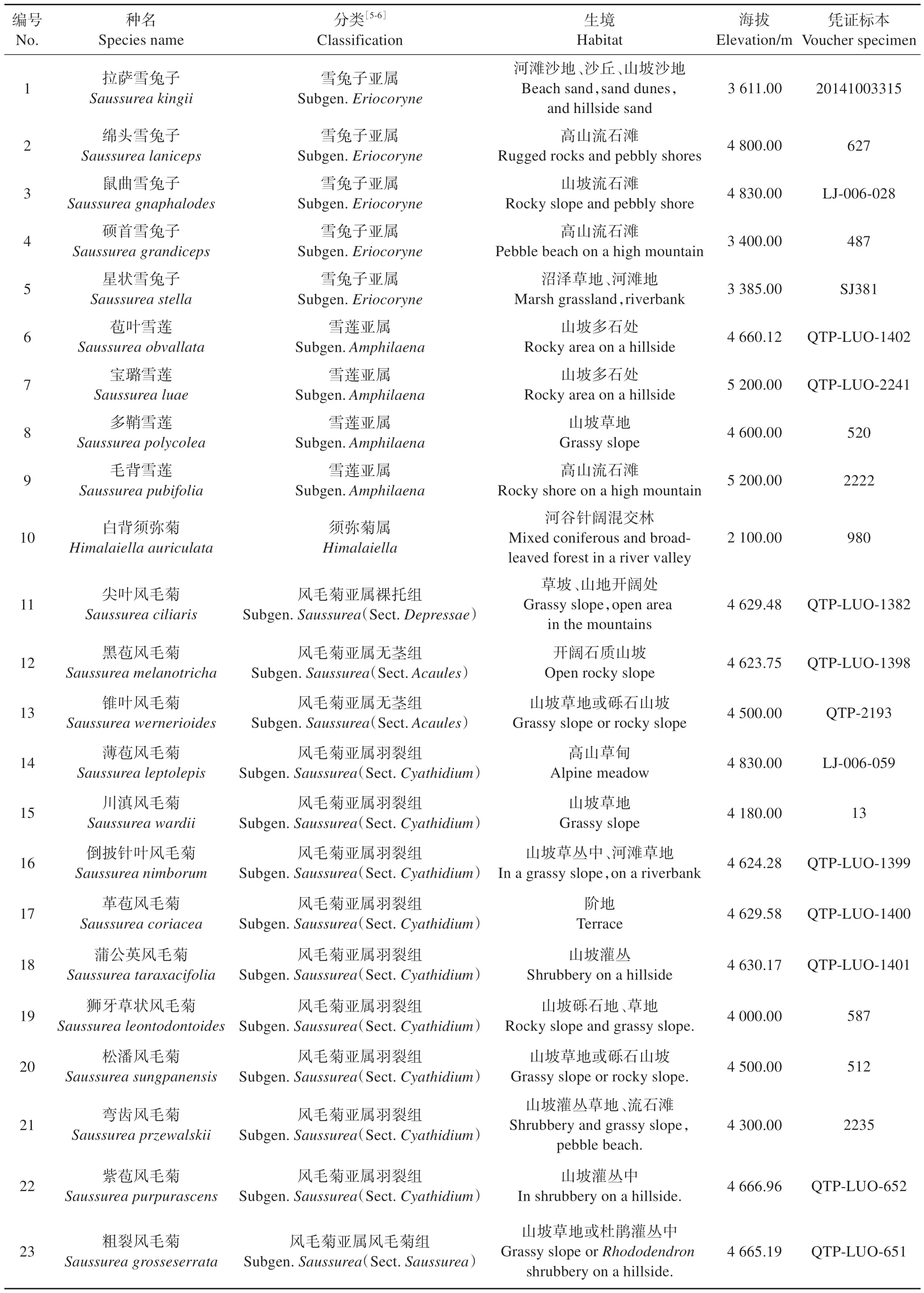
表1 实验材料基本信息Table 1 Basic information of experimental materials
所有研究材料物种的凭证标本均保存于西藏高原生态研究所标本室(XZE)。
1.2 花粉形态观察
每种在3 个不同植株的花序上分别选取5 朵成熟花,在解剖镜下,用镊子和解剖针将花药挑破置于载玻片上,滴上体积分数95%乙醇,等待干燥后,将饱满花粉均匀散布在贴有导电双面胶的样品台上,在离子溅射仪(SBC-12,产地)中真喷镀金膜,再利用电子扫描电镜(JCM-6000 型,产地)观察、拍照记录花粉形态特征。
1.3 花粉粒特征描述及数据统计
每种植物取20 粒花粉,使用SMILE VIEW Standard 软件测量其极轴长(P)、赤道轴长(E)、萌发孔宽(G)、萌发沟长(S)、体积(V)、网眼宽(M)、网脊宽(W)、刺数/400 μm2(NS)、刺宽(WS)、刺长(LS)等数量性状,花粉形态学描述术语参考王伏雄等[16]、王开发等[17]和韦仲新[29]。利用Excel 2016软件将数据处理成电子表格,花粉形状以P/E来表示,计算各项指标的平均值±标准误及偏差系数(Cv),根据公式πPE2/6[30]计算各个种的花粉体积。随后,采用SPSS 27 软件进行多重比较与聚类分析。依据P/E、花粉粒形状、萌发孔宽/萌发沟长(G/S)、刺宽/刺长(WS/LS)、萌发沟数量5 个数量性状,完成数据标准化处理,采用组间连接与欧氏距离法对23种植物花粉进行聚类分析[31]。
2 结果与分析
2.1 花粉形态
扫描电镜下,23种植物的花粉均为单粒花粉,花粉极轴和赤道轴范围为11.927~47.233 μm 和10.970~62.303 μm,其中极轴最短的花粉出现在宝璐雪莲(11.927 μm),极轴最长的见于毛背雪莲(47.233 μm);赤道轴最短者为宝璐雪莲(10.970 μm),赤道轴最长者为紫苞风毛菊(62.303 μm)。供试花粉中宝璐雪莲的极轴与赤道轴均为最小,大部分不同种间花粉粒极轴与赤道轴均存在显著差异(表2~3 和图版:I~V)。在花粉外观形态上宝璐雪莲的花粉粒体积最小(751.503 μm3),紫苞风毛菊的花粉粒体积最大(87 802.105 μm3),两者大小差异明显。宝璐雪莲的花粉为小型花粉;紫苞风毛菊和川滇风毛菊(78 456.015 μm3)为大型花粉;其他属于中等型花粉,体积大小在17 905.204~50 765.823 μm3。

表2 23种植物花粉形态特征(1)Table 2 Morphological characteristics of pollen from 23 plant species(1)
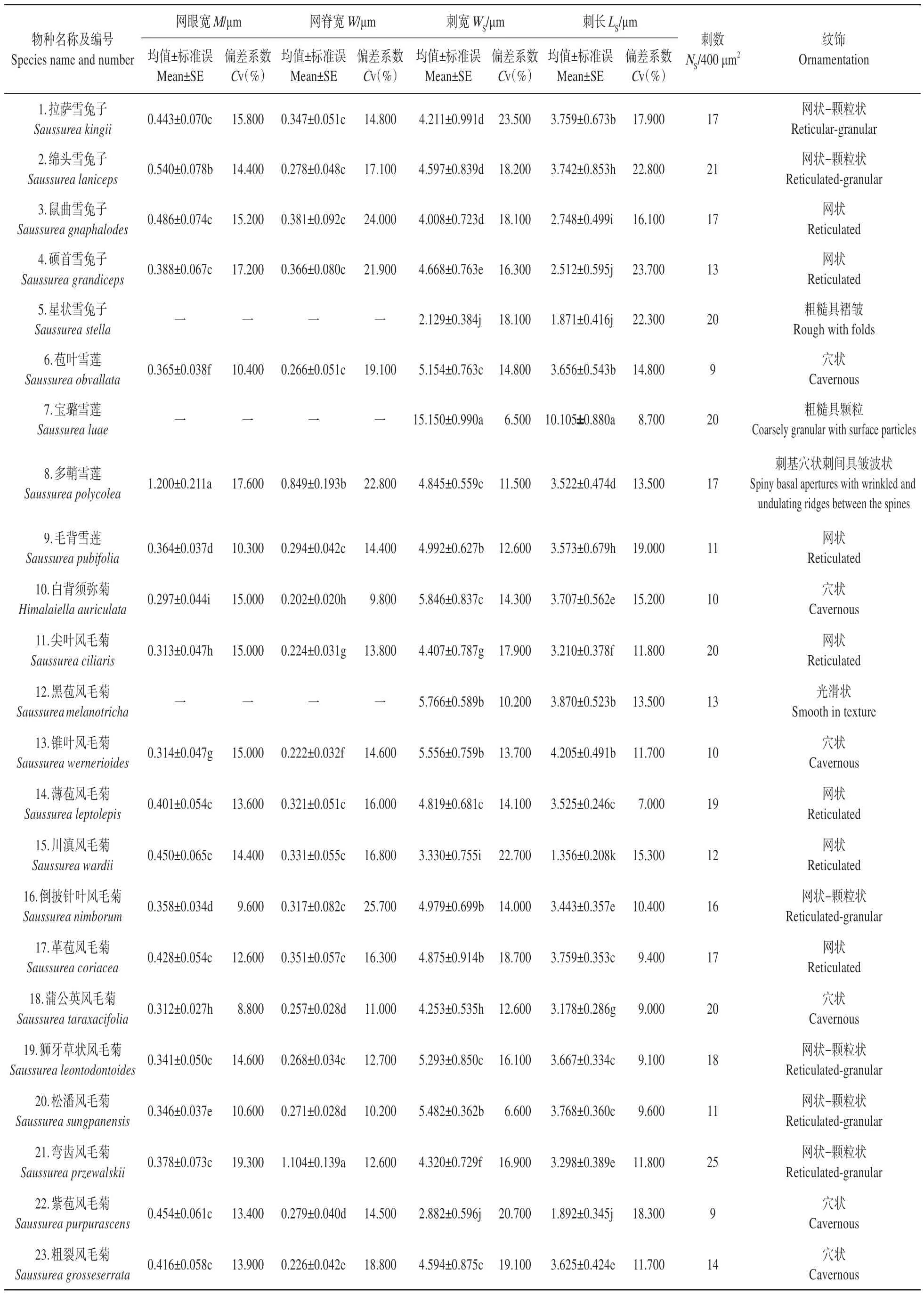
表3 23种植物花粉形态特征(2)Table 3 Morphological characteristics of pollen from 23 plant species(2)
花粉的形状根据P/E值大小来表示,当0.88≤P/E<1.14 为近球形,0.50≤P/E<0.88 为扁球形[17]。23 种植物花粉粒P/E值在0.692~1.123,分为扁球形和近球形。扁球形花粉见于:尖叶风毛菊、川滇风毛菊、紫苞风毛菊。其中,川滇风毛菊的花粉P/E值最小,仅为0.692。近球形花粉见于20 种,P/E值在0.88~1.14。其中,绵头雪兔子的花粉P/E值最大为1.123。23 种植物都为单粒花粉,极面观多为近圆形,赤道面观为近球形。说明风毛菊属植物花粉形态大小较为相似,可以作为该属花粉的普遍特征。
2.2 花粉萌发沟
花粉萌发孔沟在23种植物里除薄苞风毛菊为4孔沟外,其余22种植物的花粉均为3孔沟。花粉粒萌发沟长度为7.680~46.893 μm,在种间具显著差异性,最长和最短的长度的差值为39.213 μm,接近7 倍左右。其中,平均萌发沟最短见于宝璐雪莲,为(7.680±0.548)μm;最长的出现在川滇风毛菊,为(46.893±0.948)μm。花粉粒萌发孔宽度范围为2.603~12.588 μm,种间也有显著差异,花粉粒萌发孔宽差值为9.985 μm,其中平均萌发孔宽度最短的是尖叶风毛菊和宝璐雪莲,分别为(2.603±0.059)、(2.709±0.268)μm;平均萌发孔宽最长的是鼠曲雪兔子,为(12.588±0.880)μm。因此,花粉萌发孔沟的长度和宽度在种间差异明显。
2.3 花粉粒外壁纹饰特征
花粉粒外壁纹饰在种间差异明显,可分为7类[16-17,29](表2~3 和图版Ⅰ~Ⅴ):网状、网状—颗粒状、穴状、光滑状、刺基穴状及刺间具皱波状、颗粒状、褶皱状。现在的网状7 种(图版Ⅰ:A~E,图版Ⅱ:F~G),有鼠曲雪兔子、硕首雪兔子、毛背雪莲、尖叶风毛菊、薄苞风毛菊、川滇风毛菊和革苞风毛菊;网状—颗粒状6 种(图版Ⅱ:H~J,图版Ⅲ:K~M),有拉萨雪兔子、绵头雪兔子、倒披针叶风毛菊、狮牙草状风毛菊、松潘风毛菊和弯齿风毛菊;穴状6 种(图版Ⅲ:N~O,图版Ⅳ:P~S),有苞叶雪莲、锥叶风毛菊、蒲公英风毛菊、紫苞风毛菊、粗裂风毛菊和白背须弥菊;光滑状1 种(图版Ⅳ:T),为黑苞风毛菊;刺基穴状及刺间具皱波状1 种(图版Ⅴ:U),为多鞘雪莲;颗粒状1种(图版Ⅴ:V),为宝璐雪莲;褶皱状1种(图版Ⅴ:W),为星状雪兔子。
20 种花粉粒外壁具网眼、网脊,网眼大小在0.297~1.200 μm,多鞘雪莲网眼(1.200 μm)最大,白背须弥菊网眼(0.297 μm)最小,二者相差0.903 μm;网脊大小在1.104~0.202 μm,弯齿风毛菊网脊(1.104 μm)最大,白背须弥菊网脊(0.202 μm)最小,最大与最小网脊之间相差0.902 μm;星状雪兔子、宝璐雪莲和黑苞风毛菊3 种不具网眼与网脊。网眼差(0.903 μm)与网脊差(0.902 μm)相近。同样的,通过网眼与网脊的大小还不能将风毛菊属内各亚属清晰地区别开,但仍可作为各种间的鉴定依据,值得注意的是白背须弥菊在网脊网眼特征方面与风毛菊属其他种出现了明显的差异。
23 种植物的花粉外壁均有刺状突起,外壁刺长度为1.356~10.105 μm,其中刺长度最小的是川滇风毛菊,刺长度最大的是宝璐雪莲;表面刺基部直径在2.129~15.150 μm,其中表面刺基部直径最小的是星状雪兔子,表面刺基部直径最大的是宝璐雪莲。观察400 μm2面积的刺数,绵头雪兔子、星状雪兔子、宝璐雪莲、尖叶风毛菊、蒲公英风毛菊、弯齿风毛菊等相对较大(NS≥20),苞叶雪莲、白背须弥菊、锥叶风毛菊、紫苞风毛菊相对较小(NS≤10)。从刺状突起的3 项指标来看,植物花粉存在种间差异。白背须弥菊刺状突起与风毛菊属其他种相近似,该特征尚不能将其与风毛菊属区分开,但其刺突密度偏小,可作参考。
2.4 基于花粉性状的聚类分析
经聚类分析(图1)可知,在辅助线1(遗传距离4.0)处,将23种植物的花粉形态分为7类。第一类为雪兔子亚属中的硕首雪兔子、绵头雪兔子、拉萨雪兔子、星状雪兔子、鼠曲雪兔子性状接近;第二类为雪莲亚属(宝璐雪莲、苞叶雪莲、多鞘雪莲、毛背雪莲)、风毛菊亚属羽裂组(倒披针叶风毛菊、松潘风毛菊、狮牙草状风毛菊、弯齿风毛菊、蒲公英风毛菊、革苞风毛菊)、风毛菊亚属无茎组的黑苞风毛菊、风毛菊亚属风毛菊组的粗裂风毛菊聚集在一起;第三类为风毛菊亚属羽裂组的川滇风毛菊和紫苞风毛菊,因性状相似,都为扁球形且G/S较小聚为一类。其他4 种则单独聚为一类。以辅助线2(遗传距离11.0)来看,研究材料聚集分成3个类群,其中风毛菊亚属羽裂组的薄苞风毛菊花粉粒具4 条萌发沟,单独聚合为一类;风毛菊亚属羽裂组的川滇风毛菊、紫苞风毛菊与风毛菊亚属裸托组因性状相似,都为扁球形且G/S较小聚为第二类;风毛菊亚属无茎组锥叶风毛菊P/E值较大、G/S较大,其他性状与前面17 种风毛菊属植物相近,首先与之聚合,其次须弥菊属的白背须弥菊具备P/E较小、WS/LS较大随之聚合,形成第三类。在辅助线3(遗传距离24.0),将试供材料分成了2 个类群,其中风毛菊亚属裸托组尖叶风毛菊的P/E较小、椭圆形、G/S较小与风毛菊亚属羽裂组川滇风毛菊、紫苞风毛菊花粉特征上相似聚合在一起形成一类,另外20 种聚为一类。说明了风毛菊属花粉根据P/E、花粉粒形状、G/S、WS/LS和萌发沟数量特征,可为风毛菊属植物进行传统分类提供重要划分依据。

图1 23种植物花粉形态聚类图Fig.1 Pollen morphology clustering map of 23 plant species
3 讨论
(1)从花粉体积演化趋势的一般特征来看[16-17,29,32],宝璐雪莲体积最小,应为较进化类型;紫苞风毛菊体积最大,应为较原始类型。本次研究观测到的23种植物,除宝璐雪莲为小型花粉,紫苞风毛菊和川滇风毛菊具大型花粉,其余20 种植物的花粉极轴与赤道轴大小在25~50 μm,均属于中型花粉[17]。供试植物花粉在不同种间均有不同程度差异,花粉的极轴与赤道轴大小可作为种间划分的依据,但亚属间未表现出明显的区别,这可能与风毛菊属植物的快速辐射与趋同进化有关[33-34],23 种植物花粉极轴、赤道轴长度和花粉粒大小呈现出一定的分异,但在亚属间界限并不分明。
(2)花粉的萌发沟类型都是赤道延伸向两极并且在两极不发生联合的萌发沟,薄苞风毛菊为4 孔沟花粉,属于较为进化的种类[17,35],其余22 种花粉具3 个萌发孔沟。23 种植物花粉的在萌发沟长与萌发孔宽之间具有一定的区别,可以为该属植物系统分类提供有效的依据。
(3)花粉外壁表面纹饰是植物分类和系统发育的重要特征之一[15-17]。本研究的23 种植物花粉为风毛菊属22 种和须弥菊属1 种,通过电镜扫描发现其花粉粒外壁纹饰在3 个亚属内、4 个组内具有多个纹饰类型,根据目前对花粉微形态研究结果,外壁纹饰不同的花粉是可能存在于同一科、属、亚属及种内[23,31]。研究材料各亚属内均出现多个纹饰类型,而亚属间部分种纹饰却有类似,这可能与属内种的分化受环境变化影响有关[36]。23种植物花粉刺状突起的形态、大小和密度等各项指标在种间均存在不同程度的变异,在个别亚属间也存在显著差异,如雪兔子亚属的刺密度较风毛菊亚属的刺密度明显偏小,可作为该属植物进行系统分类的重要依据。
(4)根据P/E、花粉粒形状、G/S、WS/LS和萌发沟数量特征进行聚类,聚类结果与供试风毛菊属植物的系统分类位置表现出了一定的一致性,较大程度上支持传统分类处理。23种植物花粉性状聚类没有完全将不同亚属(组)之间清晰区分开来,白背须弥菊与风毛菊属植物之间暂不能通过花粉形态以及聚类结果将其彻底区分,但在网脊、网眼、刺突密度特征方面与风毛菊属其他种出现了明显的分化,可以为其从风毛菊属划出,置于须弥菊属提供部分证据支持。
(5)本研究中风毛菊属植物花粉形态与前人对其观测的结果[23]基本吻合,23 种植物花粉表现出一定的共性,都属于单粒花粉,且为等极的辐射对称类型,形状为近球形或扁球形,与其同为菊科的绢蒿属植物和蒿属植物的花粉形态[21-22]也较为相似,故该区域的研究结果可以为本属植物的分类提供参考。
总的来说,本研究观察的23 种植物的花粉形态特征在种间存在一定差异,对本风毛菊属植物分类有积极的参考价值,但植物分类不能仅仅依靠孢粉学上的特征,在进行分类时仍需结合宏观形态特征、种子特征、分子生物学等多方面研究的结果进行综合考量。

图版Ⅰ 扫描电镜下的5种风毛菊属植物花粉微形态A.鼠曲雪兔子;B.硕首雪兔子;C.毛背雪莲;D.尖叶风毛菊;E.薄苞风毛菊;1~3分别为花粉粒极面观、赤道面观及花粉表面纹饰Plate Ⅰ Pollen micromorphology of five Saussurea species by scanning electron microscope A.S. gnaphalodes;B.S. grandiceps;C.S. pubifolia;D.S. ciliaris;E.S. leptolepis;1-3 were polar view,equatorial view and ultrastructural view respectively

图版Ⅱ 扫描电镜下的5种风毛菊属植物花粉微形态F.川滇风毛菊;G.革苞风毛菊;H.拉萨雪兔子;I.绵头雪兔子;J.倒披针叶风毛菊;示花粉粒的极面观(J1),赤道面观(J2)和花粉表面纹饰(J3)Plate Ⅱ The pollen micromorphology of 5 Saussurea species by scanning electron microscope F.S. wardii;G.S. coriacea;H.S. kingii;I.S. laniceps;J.S. nimborum;J1-3 were polar view,equatorial view and ultrastructural view respectively
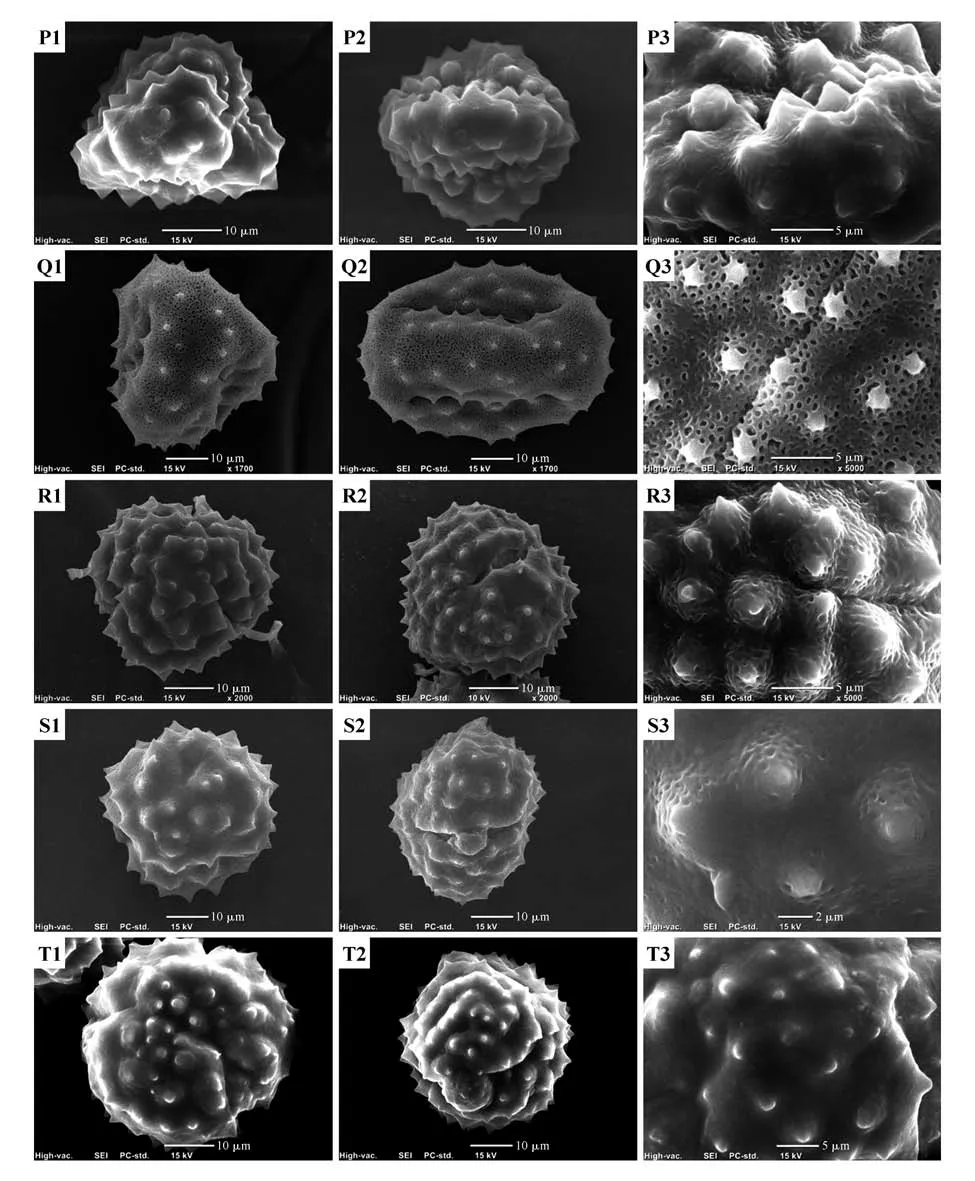
图版Ⅳ 扫描电镜下的5种风毛菊属植物花粉微形态P.蒲公英风毛菊;Q.紫苞风毛菊;R.粗裂风毛菊;S.白背须弥菊;T.黑苞风毛菊;示花粉粒的极面观(T1),赤道面观(T2)和花粉表面纹饰(T3)Plate Ⅳ Pollen micromorphology of 5 Saussurea species under SEM P.S. taraxacifolia;Q.S. purpurascens;R.S. grosseserrata;S.H. auriculata;T.S. melanotricha;T1-3 were polar view,equatorial view and ultrastructural view respectively

图版Ⅴ 扫描电镜下的3种风毛菊属植物的花粉形态U.多鞘雪莲;V.宝璐雪莲;W.星状雪兔子;示花粉粒的极面观(W1),赤道面观(W2)和花粉表面纹饰(W3)Plate Ⅴ Pollen micromorphology of 3 Saussurea species under SEM U.S. polycolea;V.S. luae;W.S. stella;W1-3 were polar view,equatorial view and ultrastructural view respectively
