不同密度点篮子鱼与文蛤混养对鱼、贝生长和肌肉营养的影响
2023-09-06孙艳秋刘鉴毅黄晓荣冯广朋
李 琪,孙艳秋,刘鉴毅,3,4,杨 俊,庄 平,3,4,赵 峰,3,4,黄晓荣,3,4,冯广朋,3,4,秦 博
(1.上海海洋大学水产科学国家级实验教学示范中心,上海 201306;2.中国水产科学研究院东海水产研究所,上海 200090;3.农业农村部东海与长江口渔业资源环境科学观测实验站,上海 200090;4.上海长江口渔业资源增殖和生态修复工程技术研究中心,上海 200090)
近年来,由于人类生产活动增多的影响,海水富营养化现象严重,海洋生态环境遭到破坏[1]。每年夏季,大型海藻肆虐,海洋环境受到严重污染,大量贝类养殖户因此备受困扰。目前处理大型海藻主要依靠药物或人工捞除的方法,然而,频繁使用药物会影响水质,加重环境负担;人工捞除则要耗费大量人力,且残留在水中的藻丝极易再次大量繁殖。点篮子鱼(Siganus guttatus),属鲈形目(Perciformes),篮子鱼科(Siganidae),篮子鱼属,为杂食性鱼类,喜食大型海藻[2-3],具有“清道夫”的美称,是当前我国健康生态养殖的混养品种之一[4],近年来被大量用于网箱和池塘的混养[5]。提高养殖资源利用率和养殖效益的有效途径之一是建立互利种类的混养[6],在贝类养殖池中混养一定数量的点篮子鱼,不需要单独投喂饵料,还可防止养殖池内藻类过度繁殖,达到养殖环境生态平衡。
目前,国内外在点篮子鱼形态、生长、繁育、食性及分子生物学等方面已有较多研究,养殖方式也从原来的单养、少量混养发展为现在的海水池塘与虾蟹混养,如黄伟卿等[7]研究了海水池塘鱼贝混养养殖模式;胡成硕等[8]探索了点篮子鱼幼鱼的行为习性及其对浒苔的摄食量,结果表明,利用点篮子鱼生态防治浒苔等大型藻类能起到很好的效果;宋超等[9]对分别摄食浒苔和人工饵料的点篮子鱼的肌肉营养成分进行了比较,结果表明,浒苔可以满足点篮子鱼的生长需求,且浒苔可以通过点篮子鱼的转化对整个生态链起到积极作用;朱晓梅等[10]研究了不同营养素配比的饲料对点篮子鱼生长及肌肉的影响,结果表明,饲料脂肪水平低有利于鱼体生长,而蛋白水平过高或过低均不利于鱼肌肉蛋白的沉积;王妤等[11]研究了不同养殖模式对点篮子鱼生长和摄食的影响,结果显示,与拟穴青蟹(Scylla paramamosain)混养的点篮子鱼生长情况优于单养模式的点篮子鱼;黄晓荣等[12]在凡纳滨对虾(Penaeusvanmamei)中套养点篮子鱼,既能改善水质,又提高了凡纳滨对虾的养殖产量和效益;柴雪良等[13]开展了在滩涂贝类池塘中养殖黄斑篮子鱼(S.oramin)以生物防治浒苔的研究,结果表明,通过混养黄斑篮子鱼可以对浒苔进行很好的防控;高宇等[14]在海参养殖区投放点篮子鱼,结果表明,浒苔覆盖率快速下降;周胜杰等[15]将白蝶贝(Inctadamaxima)与点篮子鱼混养,结果表明,丝状藻的生长可以得到有效抑制,且不影响大珠母贝的生长、存活,两种养殖物种均能正常生存。上述研究均为本实验奠定了基础。
本实验选取贝类文蛤(Meretrixmeretrix)为对象,与不同密度点篮子鱼混养,在室内初步探索贝类养殖池暴发浒苔的生物防治方法,以使能量和营养物质充分循环利用,提高养殖系统的生物容量[16-17],在单位面积上创造最大的生态效益,为构建贝类养殖池稳定的生态系统提供参考。
1 材料与方法
1.1 苗种来源
实验用点篮子鱼由中国水产科学研究院东海水产研究所海南琼海研究中心人工孵化,空运至赣榆暂养1个月后用于本实验。文蛤购自赣榆当地人工养殖池塘,浒苔来自赣榆当地海参养殖池塘。
1.2 实验方法
实验前点篮子鱼暂养于室内水泥池中,每日投喂2次人工饲料。实验于2020年8月开始,在中国水产科学研究院东海水产研究所赣榆研究中心进行了为期56 d的养殖。实验在大棚内开展,养殖桶(圆柱形,直径1 m、高1.2 m,注水高1 m)内铺10 cm厚泥沙,流水一周使泥沙完全沉降,投放文蛤密度40粒·m-2[18],设置5个点篮子鱼密度:T0(0 尾· m-3,对照组)、T1(10尾·m-3)、T2(30 尾· m-3)、T3(50尾·m-3)、T4(70尾·m-3)。每个密度设置3个重复。初始点篮子鱼体质量(2.61±1.00)g,全长(54.37±5.83)mm,体长(46.27±5.13)mm,体高(21.05±2.47)mm;初始文蛤体质量(8.67±2.01)g,壳长(31.36±2.10)mm,壳高(25.52±1.09)mm,壳厚(14.30±1.09)mm。
1.3 日常管理
关注实验桶内鱼、贝的生长情况,记录死亡个体并及时清理。间隔3 d换一次水,每次换水0.1 m3。每天投放2次新鲜浒苔(09∶00、15∶00各一次表观饱食投喂),实验期间溶解氧≥5 mg·L-1,养殖水温20~30℃,盐度19~23,光照强度100~4 663 lx。
1.4 样品采集
实验结束后,点篮子鱼饥饿处理24 h,经MS222(100 mg·L-1)麻醉,分别随机测定每个养殖桶内10尾点篮子鱼体质量、全长、体长、体高以及文蛤体质量、壳长、壳厚、壳高。随后在冰盘上迅速解剖,取出鱼内脏,用电子分析天平称取内脏重、肝重、肠重,测量肠长;取鱼体背部两侧肌肉,去鳞、去皮,置于-20℃冰箱保存待测。
1.5 指标测定及计算方法
1.5.1 生长性能、摄食指标、形体指标测定
鱼、贝增重率(weight gain rate,WGR)、特定生长率(specific growth rate,SGR)、摄食率(feed rate,FR)、饲料系数(feed conversion ratio,FCR)、存活率(survival rate,SR),以及点篮子鱼肥满度(condition factor,CF)、脏体比(viseraosomatic index,VSI)、肝体比(hepatosomatic index,HSI)、肠体比(ratio of intestine weight to body weight,ISI)和肠长比(ratio of intestine length to body length,IBR)计算公式如下:
式(1)~式(10)中,Wt为鱼、贝终末体质量(g);W0为鱼、贝初始体质量(g);t为饲喂天数(d);FI为实验期间总摄食量(g);Nt为终末个体数;N0为初个体数;L为鱼终末体长(cm);Wv为鱼内脏重(g);Wh为鱼肝脏重(g);Wi为鱼肠重(g);Li为鱼肠道长度(cm)。
1.5.2 肌肉常规营养成分测定
水分测定采用105℃干燥法(GB 5009.3—2016);粗蛋白含量测定采用凯氏定氮法(GB 5009.5—2016);粗脂肪含量测定采用索氏抽提法(GB 5009.168—2016);粗灰分含量测定采用马弗炉550℃灼烧法(GB 5009.3—2016)。
1.5.3 肌肉氨基酸测定
组织烘干后,精确称取样品置于水解管中,加入6mol·L-1盐酸5mL混匀,放入冷冻剂中冷冻,凝固后取出,在真空泵抽气管上抽真空封管。在110℃恒温干燥箱内水解24 h,冷却后定容至10 mL,0.45 μm 水系滤膜过滤除杂。吸取0.5 mL滤液置于EP管中,在真空浓缩仪内真空干燥。残留物用1 mL去离子水溶解,再干燥,进行2次溶解干燥的重复操作。最后加入1 mL pH 2.2的样品缓冲液,0.22μm水系滤膜过滤,用氨基酸自动分析仪(德国塞卡姆,S-433D型)进行分析。
1.6 肌肉营养价值评价
根据1973年联合国粮农组织/世界卫生组织(FAO/WHO)[19]以及1991年中国预防医学科学院营养与食品卫生研究所提出的评价标准[20],计算氨基酸评分(AAS)、化学评分(CS)及必需氨基酸指数(EAAI)[21],公式如下:
式(11)~式(14)中,aa为检测样品中某种氨基酸含量(mg·g-1),AA(FAO/WHO)为氨基酸评价标准含量(mg·g-1),AA(egg)为全鸡蛋蛋白中同种氨基酸含量(mg·g-1),n为比较的必需氨基酸个数,A、B、C、……、H分别为鱼肌肉蛋白中某种必需氨基酸含量(mg·g-1),AE、BE、CE、……、HE分别为全鸡蛋蛋白中各类必需氨基酸含量(mg·g-1)。
1.7 数据统计分析
实验数据先用Excel 2019处理,然后用SPSS 24.0进行单因素方差分析(one-way ANOVA),并利用Duncan多重比较法分析不同处理组之间的差异,以0.05作为差异显著水平。所有数据采用平均值±标准差(mean±SD)表示。
2 结果与分析
2.1 点篮子鱼生长、摄食、形体指标
由表1可知,T1组点篮子鱼终末体质量、增重率均显著高于T4组(P<0.05),而与T2、T3组差异不显著(P>0.05)。T4组存活率显著低于其他3组(P<0.05)。T1组特定生长率、摄食率最高,饲料系数显著低于其他3组(P<0.05)。

表1 点篮子鱼生长和摄食指标比较Tab.1 Comparison of grow th and feeding indices of Siganus guttatus
由表2可知,各试验组间点篮子鱼肥满度、肝体比、肠体比及肠长比均无显著差异(P>0.05);T1组脏体比显著高于T3、T4组(P<0.05),但与T2组差异不显著(P>0.05)。
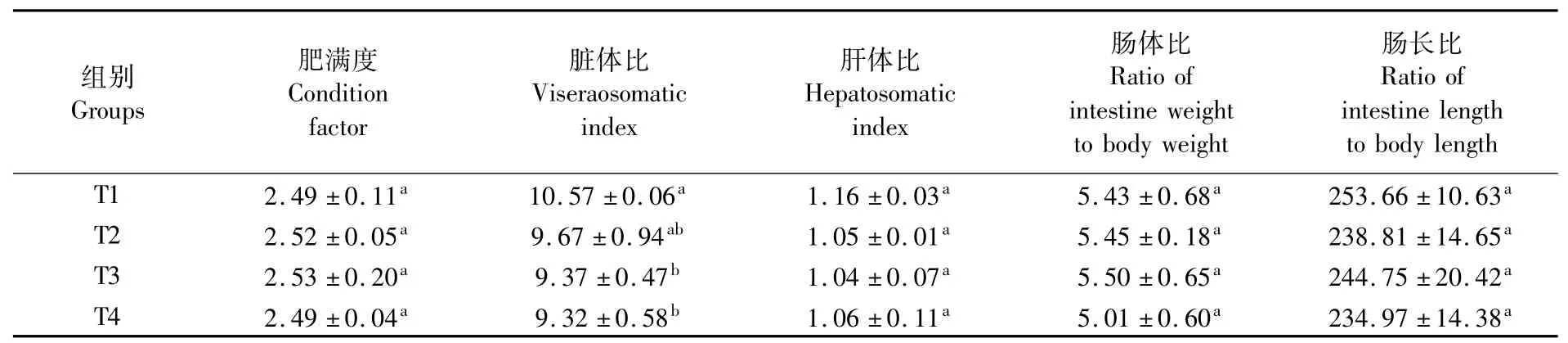
表2 点篮子鱼形体指标比较Tab.2 Com parison ofmorphometrical param eters of Siganus guttatus(%)
2.2 点篮子鱼肌肉常规营养成分
不同养殖密度下点篮子鱼幼鱼肌肉中常规营养成分如表3所示。水分含量随点篮子鱼密度升高而增加,T4试验组肌肉水分含量显著高于其他组(P<0.05);粗脂肪含量随密度升高先降低后升高,T3组粗脂肪、粗蛋白含量均显著低于其他试验组(P<0.05),T4组粗蛋白含量显著高于其他试验组(P<0.05);粗灰分随密度升高呈现先升高后降低的趋势,并在T3组达到最高且显著高于其他各组(P<0.05)。
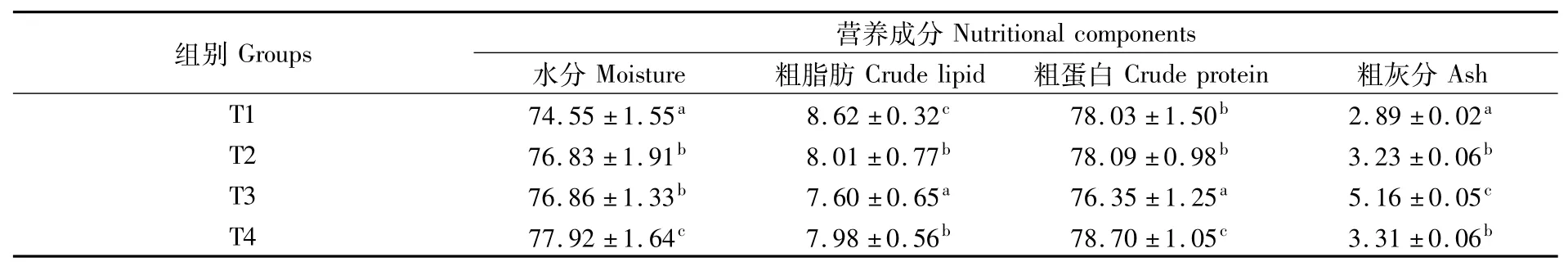
表3 点篮子鱼肌肉营养成分(干物质基础)比较Tab.3 Com parison of nutritional com ponents of Siganus guttatus(DM basis)(%)
2.3 点篮子鱼肌肉氨基酸组成与营养价值分析
2.3.1 点篮子鱼肌肉氨基酸组成
不同养殖密度下点篮子鱼肌肉中的氨基酸组成及含量见表4。所有肌肉样品中均检测到17种氨基酸(色氨酸除外),包括苏氨酸、缬氨酸、蛋氨酸、异亮氨酸、亮氨酸、苯丙氨酸、赖氨酸7种人体必需氨基酸,以及天冬氨酸、丝氨酸、谷氨酸、甘氨酸、丙氨酸、酪氨酸、脯氨酸7种非必需氨基酸和胱氨酸、组氨酸、精氨酸3种半必需氨基酸。
T3、T4组肌肉中总氨基酸含量显著高于T1、T2组(P<0.05)。在各组所有呈鲜味氨基酸中,谷氨酸含量均是最高的;T4组谷氨酸、甘氨酸含量均显著高于其他3组(P<0.05);T3、T4组天冬氨酸含量显著高于T1、T2组(P<0.05),并在T4组最高,为(8.86±0.09)%。在半必需氨基酸组成中,精氨酸的含量在各组均是最高的;其中,T4组精氨酸含量最高,为(4.97±0.11)%,显著高于T1、T2组(P<0.05),但与T3组差异不显著(P>0.05);T4组组氨酸、胱氨酸含量最低,但4组间组氨酸、胱氨酸含量均无显著性差异(P>0.05)。非必需氨基酸组成中,T3、T4组丝氨酸含量显著高于T1、T2组(P<0.05),且T4组最高。在必需氨基酸组成中,赖氨酸的含量在各组中均最高;苏氨酸、缬氨酸、亮氨酸、异亮氨酸、苯丙氨酸以及赖氨酸含量均在T4组中最高,但均与T3组无显著性差异(P>0.05);蛋氨酸含量在T3组最高,为(2.61±0.08)%,但与T2、T4组无显著性差异(P>0.05)。
2.3.2 点篮子鱼肌肉营养品质评价
由表5可知,4个试验组点篮子鱼肌肉的缬氨酸含量均低于FAO/WHO氨基酸模式,其他各类氨基酸均高于FAO/WHO氨基酸模式。T3组氨基酸总量最高。
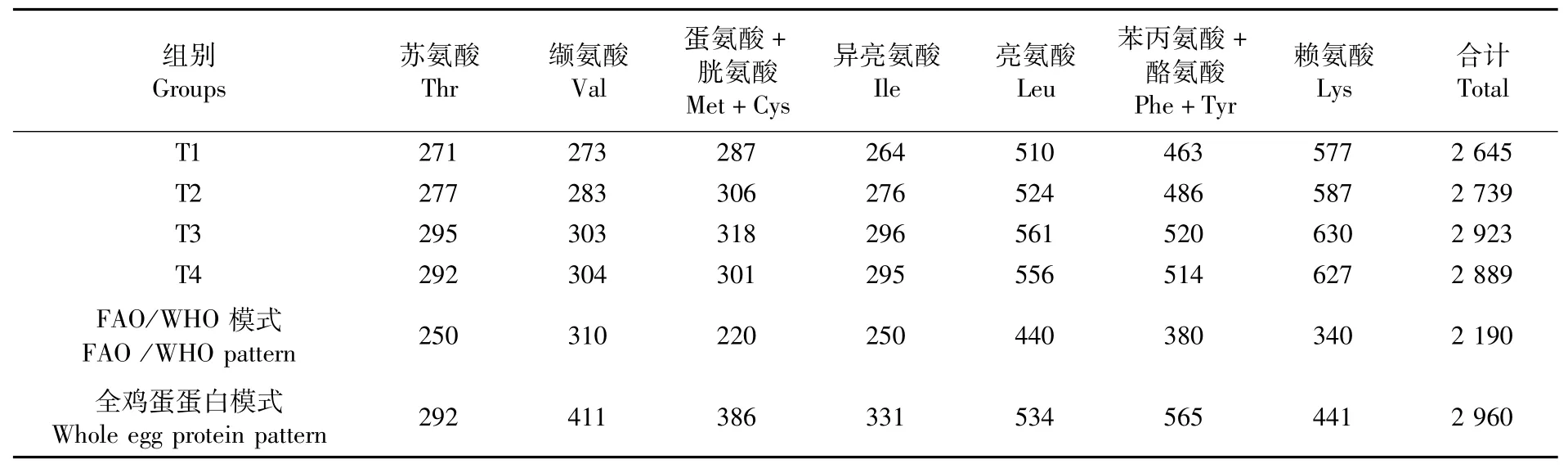
表5 点篮子鱼肌肉中必需氨基酸含量与FAO/WHO和全鸡蛋蛋白模式比较Tab.5 Comparison of essential am ino acid contents in muscle of Siganus guttatus w ith FAO/WHO pattern and whole egg protein pattern(mg·g-1)
根据公式分别计算出4组点篮子鱼肌肉的AAS、CS和EAAI,结果见表6。根据表6可以看出,4组氨基酸评分均是赖氨酸最高,缬氨酸最低,因而各组第一限制氨基酸均为缬氨酸。T1、T2、T3、T4组EAAI分别为94.90、86.92、96.11、90.16,从EAAI来看,T3组点篮子鱼肌肉必需氨基酸组成优于其他各组。
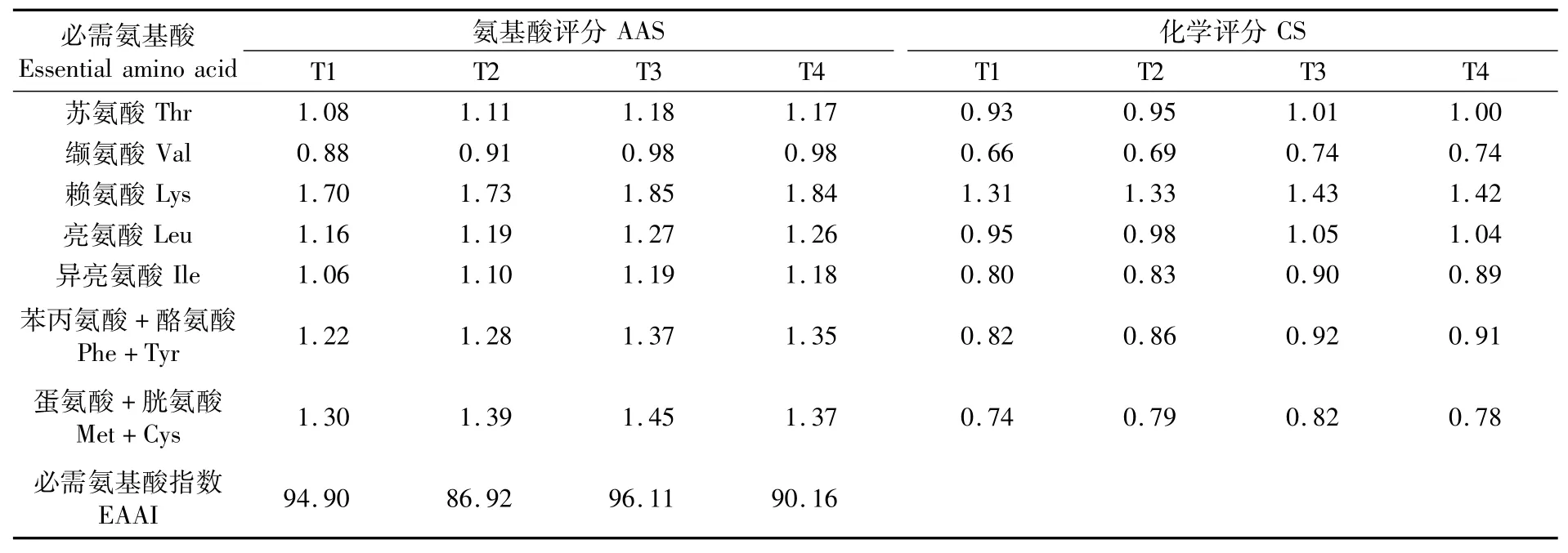
表6 点篮子鱼肌肉中必需氨基酸氨基酸评分(AAS)、化学评分(CS)和必需氨基酸指数(EAAI)比较Tab.6 Comparison of AAS,CS and EAAI of essential am ino acid in muscle of Siganus guttatus
2.4 文蛤生长指标
由表7可知,各组文蛤存活率、终末体质量及增重率均无显著差异(P>0.05)。

表7 文蛤生长指标比较Tab.7 Comparison of grow th indices of Meretrix meretrix
2.5 文蛤肌肉常规营养成分
实验结束时,不同点篮子鱼养殖密度下文蛤肌肉中的常规营养成分测定结果见表8。粗脂肪含量随点篮子鱼密度的升高而升高;T2组粗蛋白含量显著高于其他组(P<0.05),粗灰分含量显著低于其他组(P<0.05);T4组文蛤肌肉中水分、粗脂肪含量均显著高于其他组(P<0.05)。

表8 文蛤肌肉营养成分比较Tab.8 Comparison of common nutritional components of Meretrix meretrix(%)
2.6 文蛤肌肉氨基酸组成与营养价值分析
2.6.1 文蛤肌肉氨基酸组成
不同组文蛤肌肉中的氨基酸组成及含量见表9,所有文蛤肌肉样品中均检测到同点篮子鱼一样的17种氨基酸(色氨酸除外)。
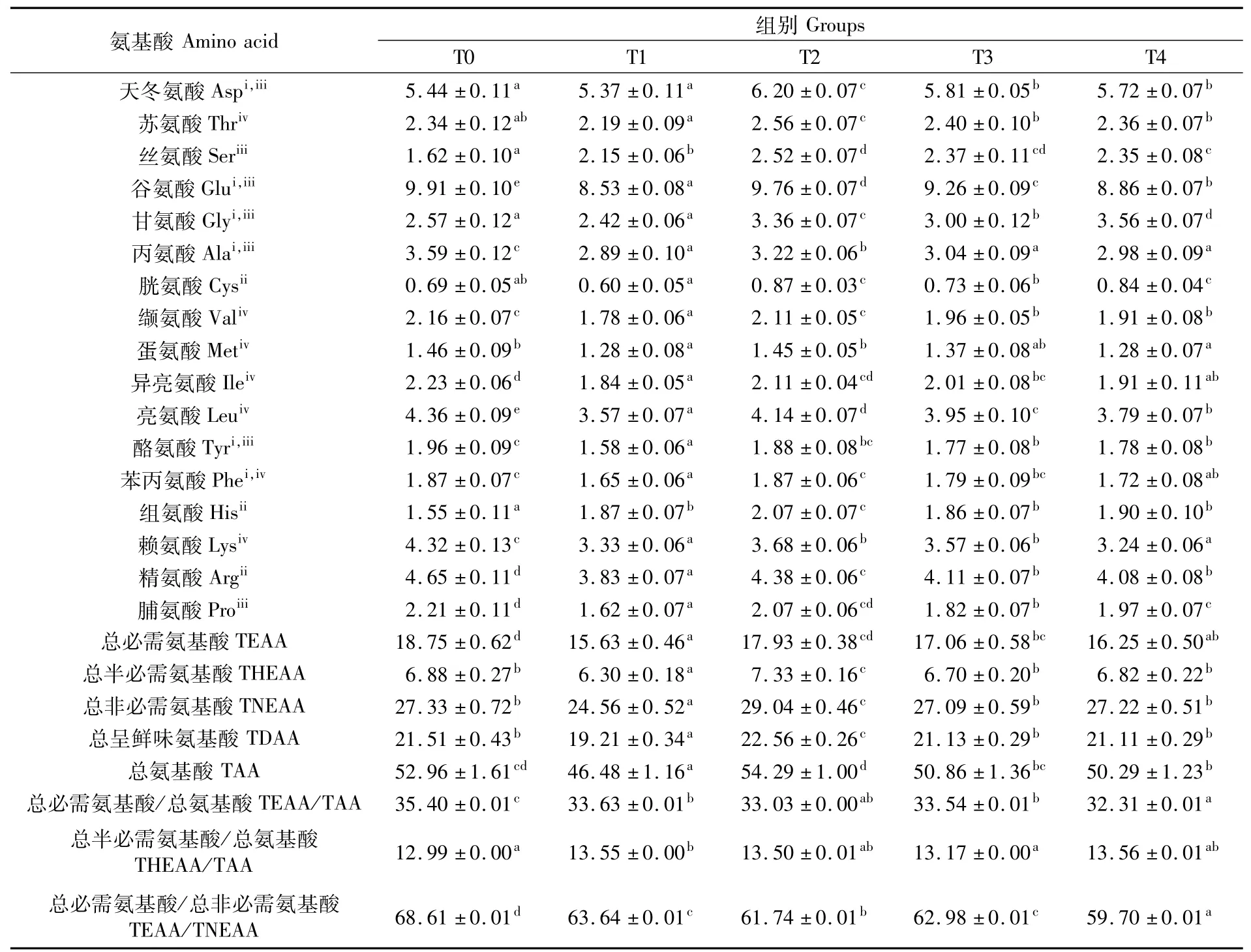
表9 文蛤肌肉氨基酸组成比较Tab.9 Com parison of am ino acid composition in muscle of Meretrix meretrix(%)
T1组肌肉中总氨基酸含量显著低于T0、T2、T3、T4组(P<0.05);T2组总氨基酸含量显著高于T1、T3、T4组(P<0.05),但与T0组差异不显著(P>0.05)。与点篮子鱼相同,呈鲜味氨基酸组成中,谷氨酸含量在各组中均高于其他呈鲜味氨基酸;4个试验组天冬氨酸、谷氨酸和丙氨酸含量均随点篮子鱼密度升高呈现先升后降的趋势,T2组显著高于其他3组(P<0.05);甘氨酸在T4组中含量最高;T1组各类呈鲜味氨基酸含量均低于对照组,T2、T3、T4组谷氨酸和丙氨酸含量均低于对照组。半必需氨基酸组成中,各组精氨酸含量均高于其他半必需氨基酸;对照组精氨酸含量为(4.65±0.11)%,显著高于其他组(P<0.05);T2组胱氨酸和组氨酸含量均高于其他组。非必需氨基酸组成中,T2组丝氨酸含量高于其他组;对照组酪氨酸、脯氨酸含量均高于其他组。必需氨基酸组成中,4个试验组各类氨基酸含量均随点篮子鱼养殖密度的升高呈现先升后降的趋势;各组中亮氨酸含量均高于其他必需氨基酸;T2组苏氨酸含量显著高于其他组(P<0.05);缬氨酸、蛋氨酸、异亮氨酸、亮氨酸和赖氨酸含量均在对照组最高。
2.6.2 文蛤肌肉营养品质评价
由表10可知,各组必需氨基酸总量均低于FAO/WHO氨基酸模式。根据公式分别计算出各组文蛤肌肉中的AAS、CS和EAAI,结果见表11。氨基酸评分,T0、T1、T2、T3组均是赖氨酸评分最高,T4组蛋氨酸+胱氨酸评分最高;各组中评分最低的均为缬氨酸,这与点篮子鱼情况相同,因而各组第一限制氨基酸均为缬氨酸。T0、T1、T2、T3、T4组EAAI分别为68.20、61.01、66.03、64.89、64.21,从EAAI来看,T2组的必需氨基酸组成优于其他3个试验组。
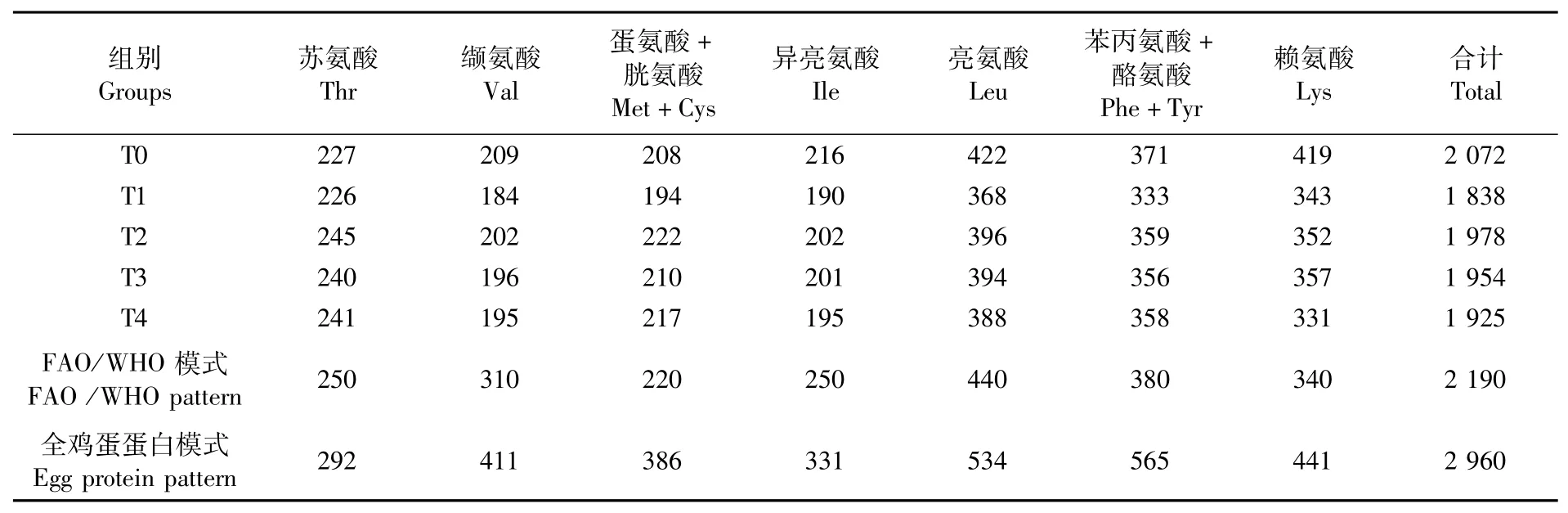
表10 文蛤肌肉中必需氨基酸含量与FAO/WHO和全鸡蛋蛋白模式比较Tab.10 Comparison of essential am ino acid contents in muscle of Meretrix meretrix w ith FAO/WHO pattern and whole egg p rotein pattern(mg·g-1)
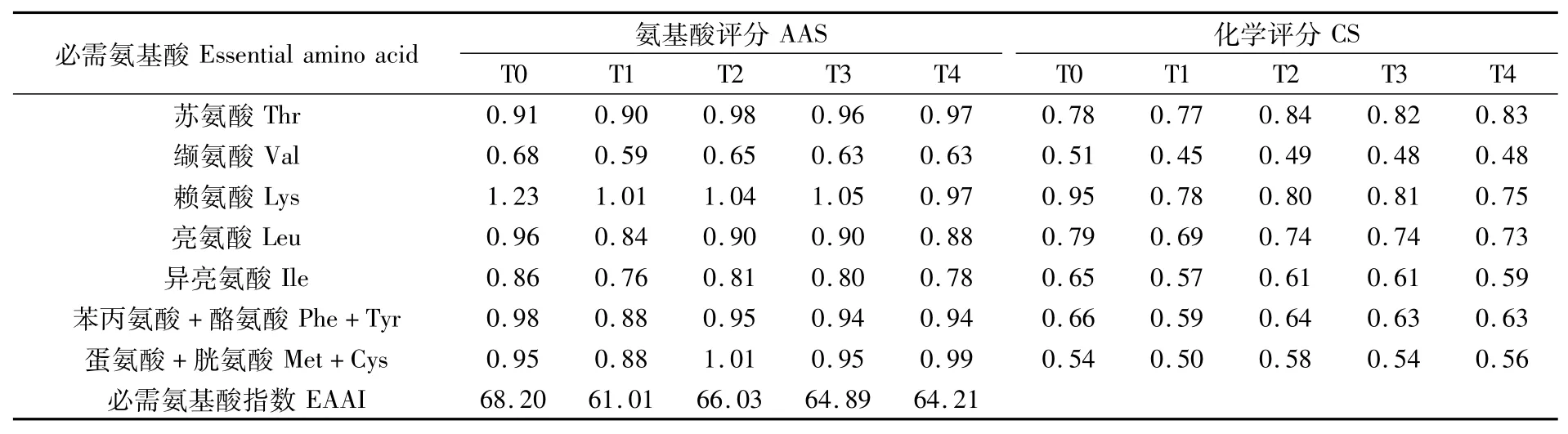
表11 文蛤肌肉中必需氨基酸的氨基酸评分(AAS)、化学评分(CS)和必需氨基酸指数(EAAI)比较Tab.11 Comparison of AAS,CS and EAAI of essential am ino acid in muscle of Meretrix meretrix
3 讨论
3.1 点篮子鱼放养密度对鱼、贝生长的影响
饲料、环境条件及群体密度等均是影响鱼、贝生长的重要因素[22]。养殖密度增加,养殖对象对饲料和空间的竞争加剧,引发摄食、代谢及免疫等多种变化。本实验显示,点篮子鱼放养密度可显著地影响鱼的生长,与邹雄等[23]的研究结果相似。在高密度养殖条件下,其增重率、特定生长率及摄食率较低,原因在于混养过多点篮子鱼加剧了空间竞争,摄食浒苔时点篮子鱼要随着浒苔在水中上下游动,消耗大量能量,进而影响鱼类的生长[24]。T1组点篮子鱼饲料系数最低,说明该组饲料转化率高,实现了饲料利用最大化。研究表明,动物体质量的变化取决于能量得失[25-26]。高密度组中点篮子鱼获得的能量主要用于维持正常的生命活动,没有过多用于自身生长。消化吸收能力会显著影响动物的能量收支[27],其重要衡量指标为形体指数。本实验中,T1组的肝体比、脏体比最高,是因为大量营养物质在鱼肝脏及内脏内聚集,从而呈现出较高的指标。个体通过调节消化道形态进而提升自身消化能力,获取更多能量以适应环境,是个体遭受环境胁迫时维持自身能量平衡的一种方式。小肠作为消化吸收的主要场所,其形态结构的改变能够反映鱼类对能量需求的程度。T4组点篮子鱼通过降低体质量和改变消化道形态结构,以减少基础能耗和提升摄入能量的能力,来缓解生存压力。关于鱼、贝与藻类混养过程中的能量代谢机制,仍需进一步深入研究。
本实验得出,点篮子鱼放养密度对文蛤的生长指标没有显著影响,可能是因为时间、环境等因素未使文蛤滤食的藻类大量繁殖,因而不同组文蛤生长差异不明显。但是从文蛤软体组织肌肉营养成分可以看出,点篮子鱼密度对其生长也产生了一定的影响。
3.2 点篮子鱼放养密度对鱼、贝肌肉营养的影响
肌肉营养成分组成可以体现其营养价值[28-29]。氨基酸是组成蛋白质的基本单位,参与蛋白质、酶类及激素的合成,鱼、贝对蛋白质的需求实际上是对必需氨基酸的需求[30-31],氨基酸质量决定着蛋白质质量及肌肉营养品质[21,32]。高密度养殖条件下,鱼体为适应环境分泌大量激素,因此本实验T4组点篮子鱼肌肉粗蛋白含量最高;由于该组点篮子鱼密度高,鱼排泄物较其他组多,有利于文蛤滤食的微型藻类繁殖,贝类也易获得更多营养,因此T4组文蛤肌肉粗蛋白、粗脂肪含量较其他组高。各组点篮子鱼肌肉中总必需氨基酸与总氨基酸的比值(TEAA/TAA)和总必需氨基酸与总非必需氨基酸的比值(TEAA/TNEAA)均符合FAO/WHO的理想模式,即TEAA/TAA 为40%左右、TEAA/TNEAA 在60%以上[21],表明各组点篮子鱼肌肉中优质蛋白质含量丰富。然而,T4组文蛤由于总非必需氨基酸含量高,总必需氨基酸含量略低,TEAA/TNEAA未达到60%。本研究中,各组鱼、贝肌肉中含量最高的氨基酸均为谷氨酸,这与宋超等[9]得出的结果相同。谷氨酸是重要的呈鲜味氨基酸[33],是多种生化代谢反应的原料[34]。呈鲜味氨基酸除谷氨酸外,具代表性的还有天冬氨酸,动物肌肉的鲜味取决于呈鲜味氨基酸在总氨基酸中的占比[35-36],它们的合成是通过调节相关酶的合成进而改变三羧酸循环以及糖酵解等途径来实现的[36]。鱼肌肉中酪氨酸含量反映了肌肉总蛋白的代谢水平,T4组点篮子鱼肌肉中酪氨酸含量较其他组高,蛋白质降解较快,导致鱼的生长速率较低。赵志军[37]指出,食物的易获程度、质量差异、分布范围等是影响动物能量获取的重要因素,T1组点篮子鱼密度低,食物易获得,能获取足够的能量用于自身生长,因此呈现出较好的生长状态。T4组鱼、贝肌肉中呈鲜味氨基酸含量较高,可能是因为环境有利于相关酶的合成,促进了呈鲜味氨基酸的合成。半必需氨基酸组成中,精氨酸的含量均比其他半必需氨基酸高,这与李鑫等[38]比较不同光周期下欧洲舌齿鲈(Dicentrarchuslabrax)肌肉营养成分得到的结果相同。本实验得出,缬氨酸的氨基酸评分最低,各组第一限制氨基酸均为缬氨酸,这与李鑫等[38]研究结果相似,而宋超等[9]在比较摄食浒苔和人工饵料的点篮子鱼肌肉营养成分中发现,蛋氨酸+胱氨酸评分最低,这可能是由于所摄食的浒苔种类不同造成的差异。必需氨基酸指数(EAAI)是评价蛋白质营养价值的指标之一[29],点篮子鱼肌肉氨基酸EAAI评分由高到低依次为T3、T1、T4、T2,文蛤肌肉氨基酸EAAI评分由高到低依次为T0、T2、T3、T4、T1,反映了各组鱼、贝必需氨基酸组成的优劣及蛋白质质量的高低。
3.3 与其他混养模式比较
本实验发现,是否混养点篮子鱼对文蛤生长没有显著影响,而点篮子鱼的生长则受到了显著影响,这可能是因为:一方面,水体中藻类没有足够的时间大量繁殖,对文蛤生长的影响不明显;另一方面,文蛤达到一定阶段后生长会变缓慢,本实验所用文蛤规格偏大,处于生长缓慢阶段。周胜杰等[15]将白蝶贝与点篮子鱼混养,贝类生长没有显著差异,而鱼类生长存在显著差异。常杰等[39]将凡纳滨对虾、青蛤(Cyclinasinesis)和菊花心江蓠(Gracilarialichevoides)以不同比例混养,提高了对初级生产力和凡纳滨对虾残饵的利用率,而各处理组青蛤产量没有显著差异,本实验研究结果与上述研究结果相似。张鹏等[40]对条石鲷(Oplegnathusfasciatus)、文蛤和龙须菜(Asparagusschoberioides)3种养殖生物进行了不同养殖模式的实验,发现混养可使文蛤和条石鲷都得到更好的生长。以上研究结果的差异可能是由于混养的鱼的种类、混养环境(温度、盐度等)不同而造成的。
通过在贝类养殖池套养点篮子鱼,不仅可以抑制养殖水体内浒苔暴发,增加水体利用率,改善养殖环境,提高尾水排放标准,还比采用人工打捞浒苔的方法降低了成本,相比使用除草剂则降低了药残风险,是一种绿色生态型水产养殖模式。
4 小结
不同密度点篮子鱼与文蛤混养对鱼、贝生长、摄食和肌肉营养成分会产生不同程度的影响。对点篮子鱼来说,放养密度在10 ~30尾·m-3时最利于其生长,50尾·m-3时肌肉营养品质最佳;对文蛤来说,点篮子鱼放养密度对其生长没有明显影响,放养点篮子鱼密度30尾·m-3时,既能控制贝类养殖池中浒苔的生长,又能使文蛤肌肉营养品质最佳。养殖户可根据自身需要投放适宜密度的点篮子鱼,以便既能防治浒苔暴发又可增加经济效益。
