6年生思茅松半同胞子代生长性状的遗传分析及选择*
2023-09-02陈伟孟梦李江刘永刚冯弦
陈伟,孟梦,李江,刘永刚,冯弦
(云南省林业和草原科学院,云南 昆明 650201)
思茅松(Pinuskesiyavar.langbianensis)是我国特有的一个速生针叶树种,主要分布于云南南部,具有速生、适应性强、材性优良、松脂产量高等特点,围绕思茅松已形成涵盖林化、林纸、林板、林下经济等门类较全的林产业体系[1-2]。前期思茅松的研究主要服务于产业需求,围绕苗木培育[3]、空间配置[4]、病虫害防治[5]及科学经营[6]等人工林培育技术开展,在遗传改良方面起步较晚,20世纪90年代建成了第一个无性系初级种子园,但长期停滞于一代改良阶段[7],相较于进入第三代遗传改良的杉木(Cunninghamialanceolata)[8-9]和全面进入第二代遗传改良的马尾松(Pinusmasonnianna)[10-11]等针叶树种,具有较大的差距,已成为制约思茅松下游相关产业快速发展的一个重要因素。
常规的林木遗传改良是一个长周期的过程,遗传测定是其中的核心工作[12-13]。在一个林木育种计划中,加性方差、遗传力、遗传相关、育种值等各种遗传参数的精准估算是科学制定育种策略的重要依据[14]。子代测定是林木育种中使用较多的一种遗传测定方法[15],其中,半同胞子代测定在林木遗传改良早期阶段使用较多,在初级种子园经营管理、二代种质创制亲本选择及二代育种材料来源方面都发挥着巨大的贡献。测定是为了更好地选择,基于育种值的选择以基因型进行性状的选择,能够极大地提高选择的效率和精确度,育种值无法直接观测,需要通过表型性状信息进行估算,育种值估算方法的不同可能导致预测精度的差异[16]。育种值估算的方法有很多[16],从早期的小区平均值法,到20世纪中叶的选择指数法,到现在使用较为广泛的最佳线性预测法(BLP)和最佳线性无偏预测法(BLUP),因地制宜的选择评估方法有利于准确预估育种值。在针叶树种遗传改良实践中,BLUP方法应用较广,该方法在群体规模大,结构复杂,数据不平衡等条件下仍然能够较为准确地估算育种值,其方法的可靠性在日本落叶松(Larixkaempferi)育种值的估算中得到了很好的验证[16]。科学估算育种值、基于育种值开展性状的选择对于思茅松遗传改良工作具有重要的意义。
应用好前期试验数据,科学制定长期的育种策略,是推动林木遗传改良快速发展的关键工作[13,17]。思茅松一代改良材料的广泛应用,积累了大量的表型数据,如何利用好这些数据进行科学分析,对前期遗传改良的成效进行评估,更好地制定出针对思茅松树种特性和现有基础的长期育种策略,是当前思茅松遗传改良工作中亟待解决的科学问题。基于此,对2000年定植的思茅松半同胞子代测定林数据进行分析,科学评估参试材料及其亲本的遗传特性,基于育种值开展优良遗传材料的选择,以期更好地服务于思茅松高世代遗传改良工作。
1 材料与方法
1.1 试验地概况
试验地位于云南省普洱市思茅区南屏镇(22°40′~22°44′N、100°50′~100°58′E),气候类型为南亚热带季风气候,干湿季分明,具有优越的水热条件,年均气温 17.7 ℃,≥10 ℃ 有效积温6 253.5 ℃,无霜期长达350 d,年均降水量 1 547 mm,年平均相对湿度 82%[18],适宜发展热区人工林。土壤类型为赤红壤,地带性的原生植被为季风常绿阔叶林。造林前植被类型是以思茅松为主要成分的针阔混交林,主要乔灌木树种有西南木荷(Schimawallichii)、麻栎(Quercusacutissima)、云南黄杞(Engelhardiaspicata)、红皮水锦树(Wendlandiatinctoria)、西南山茶(Camelliareticulata),草本植物主要有紫茎泽兰(Ageratinaadenophora)、山红稗(Carexbaccans)、野拔子(Elsholtziarugulosa)等。
1.2 试验林设置情况
试验林共40个家系参与测定,其中32个来源于西双版纳傣族自治州景洪市普文镇思茅松无性系种子园,为普文测定点初步选出的优良家系,8个为新测定的家系,其母本优树采用5株优势木对比树法在普洱市和西双版纳州思茅松天然林中选出,以1个当地商品种作对照。塑料容器育苗,选择“百日苗”中的一级苗造林,随机完全区组设计,5次重复,6株方形小区,株行距2 m×3 m,在试验区的四周各设置一行保护带,定植穴规格40 cm×40 cm×40 cm,每株底肥施复全肥150 g,定植当年9月和11月各除草一次,以后每年除草2次,连续抚育3年。
1.3 数据调查与分析方法
1.3.1 表型数据测定
定植当年测定试验林的保存率和生长情况,2006年对试验林进行生长性状调查,2022年再对试验林开展调查,受前期自然灾害和人为因素的影响,试验林保存率低于40%,缺数据不平衡较为严重,无统计价值,故仅以6 a生的数据进行后续分析。
数据测定时,树高数据采用激光测高仪测量,胸径数据采用胸径尺测定,单株材积按原林业部颁布的标准[19]公式①计算:
V=0.000 051 577 714D1.985 218×H0.920 350 96
①
1.3.2 遗传参数估计
采用单株线性随机效应模型和限制性最大似然估计方法估算随机效应中的方差分量[20]。
y=μ+Xb+Z1f+Z2fb+e
②
②式中:y为观测值向量,μ为家系性状观测值的平均值,b为固定的区组效应,f为随机的家系效应向量,fb为家系与区组互作效应向量,e为随机的残差,X、Z1、Z2分别为相对应效应的关联矩阵[14]。
利用似然比检验各方差组分的统计显著性,利用泰勒级数展开法计算遗传参数的标准误[20]。
③
④


⑤
⑥
遗传增益[21]:
⑦
表型相关系数(rp),遗传相关系(rg)数按公式⑧、⑨计算:
⑧
⑨
育种值通过BLUP法进行估算[22]。
1.4 数据处理与分析
采用Excel进行数据的录入和转换,方差分析、遗传参数和育种值的估算采用R软件和ASReml-R包进行分析[23]。
2 结果与分析
2.1 半同胞家系间生长性状的差异性分析
对参试的思茅松半同胞家系子代生长性状进行统计(表1),6 a生植株家系树高、枝下高、胸径和材积的平均值分别为6.16 m、1.78 m、9.00 cm和23.26 ×10-3m3,在生长性状的变异性方面,材积的变异系数最大,达到了41.70%,枝下高和胸径次之,树高相对较小,但也超过了15%,观测的4个表型性状丰富的变异适宜开展遗传分析和选择。进一步的方差分析显示,树高、胸径和材积在不同家系间的差异达到了极显著的水平(P<0.01)。
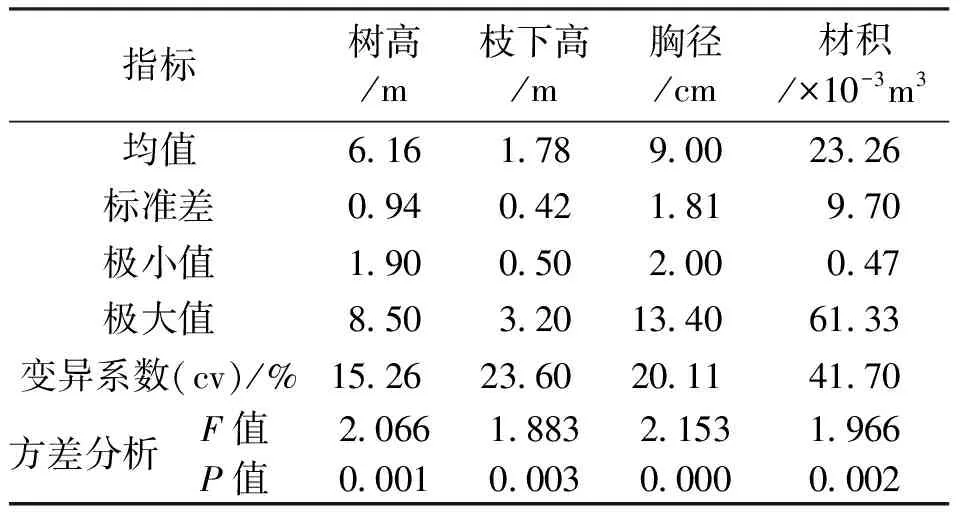
表1 思茅松半同胞家系生长性状的描述性统计
2.2 生长性状的遗传参数估算
对参试的思茅松半同胞家系子代生长性状数据进行正态性分析,树高、胸径和材积数据的残差值均符合正态性(图1),适宜使用线性随机效应模型进行后续分析。利用限制性最大似然估计方法对3个生长性状中随机效应的方差分量进行估算(表2)。

图1 子代生长性状数据的正态分布

表2 半同胞家系生长性状遗传参数估算
树高的表型变异系数和遗传变异系数分别为15.29%和10.03%,胸径的表型变异系数和遗传变异系数均比树高大,说明思茅松在幼龄林时其径向生长的遗传变异程度较高生长更大,差异更为丰富,开展胸径选择的价值相对于树高也更大;单株材积性状的遗传变异较树高和胸径大,其表型变异系数和遗传变异系数分别达到了41.73%和20.82%,是胸径表型变异系数和遗传变异系数的2.06倍和1.85倍。遗传变异幅度越大,选择潜力越大[12],材积性状更适宜作为遗传选择的性状。
在遗传力方面,树高、枝下高、胸径和材积的家系遗传力均超过了0.4,分别达到了0.521、0.564、0.540和0.504,按遗传力高低评判标准[12],4个性状的家系遗传力均属于高遗传力,受较强的遗传控制;树高性状的单株遗传力0.430,属于高遗传力;枝下高、胸径和材积的单株遗传力分别为0.270、0.311和0.249,属于中等遗传力。
思茅松4个生长性状在不同选择强度下的遗传增益(表3)显示,生长性状随着选择强度的降低,其遗传增益相应降低,树高性状在5%、10%和20%入选率条件下,遗传增益分别可达到14.91%、12.66%和10.13%;鉴于较高的遗传力,枝下高、胸径性状在入选率降低到20% 的情况下,遗传增益仍然可以大于10%;而以材积作为生长性状的主要选择指标进行选择,可获得较好的遗传增益,5%入选率的遗传增益可达到30.47%,降低到20%的入选率后仍然可以获得大于20.71%的遗传增益,选择效益较好。

表3 不同选择强度下的遗传增益
基于生长性状的加性遗传方差、两两性状的协方差和误差方差计算出了树高、枝下高、胸径和材积性状间的遗传相关和表型相关关系(表4)。在遗传相关关系上,4个性状间均具有极显著的正相关关系(P<0.01),而以材积与胸径间遗传相关系数最大,达到了0.981,材积与枝下高的遗传相关系数最低,仅0.146;表型相关方面,4个性状间也具有极显著的正相关关系(P<0.01),相关系数在0.341~0.957之间,以材积与胸径性状间最大。
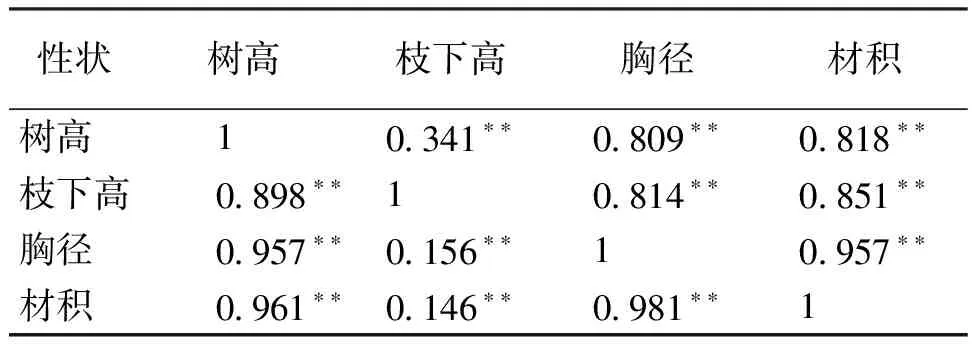
表4 不同生长性状相关分析
2.3 优良家系的选择
鉴于材积性状具有较高的遗传变异系数和遗传力,以材积性状指标开展优良家系的后向选择,对40个参试家系材积性状的均值、变异系数进行统计,采用BLUP(最佳线性无偏预测)法完成参试家系的育种值估算(表5)。思茅松不同家系6 a生植株材积均值的变幅为10.95×10-3~28.43×10-3m3,变异系数的变幅6.54%~83.30%,家系间和家系内部都具有较大的变异幅度,其中有33个家系的材积均值超过了对照(20.45×10-3m3)。育种值是选择育种中的重要参数[24],以材积育种值为选择指标,对参试家系的育种值进行排序,按10%的入选率,选择出4个优良家系,分别为06号、24号、35号和21号,入选家系材积均值较对照高30.93%~39.07%。

表5 半同胞家系材积变异与育种值估计
3 讨论与结论
3.1 讨论
6 a生思茅松的树高、胸径和材积家系遗传力均超过0.5,数值上与赵文书等[25]估算的遗传力有差异,这与遗传力为估算值有关,遗传参数的估算不仅与测定材料自身的变异特性有关,还受试验设计、立地条件、试验林的管理、测定年龄及估算方法等许多因子的影响,是与特定群体结构密切相关的一个动态函数[26],但从遗传力的强度划分上,两次估算出的6 a生思茅松3个性状的遗传力均属于高遗传力性状,在幼龄林阶段受到较高程度的遗传控制,但其遗传力是否会随着林龄的增长而变化,尚有待进行连续观测。
当两个性状存在相关,且相关系数趋近于1时,间接选择的效果能接近于直接选择[12]。本研究在6 a生思茅松生长性状中,胸径和材积的遗传相关性达到了极显著,胸径和树高的遗传相关性也达极显著,这为间接选择提供了可能。对于进入中龄林以后的思茅松测定林,当树高指标不易获得或是难以准确获得时,可以用胸径指标替代材积指标对材积进行间接选择。
遗传变异和选择是林木育种的两个关键环节。基于遗传测定结果进行科学的选择有利于最大程度的挖掘优质的遗传材料。育种值是选择育种中的重要参数,它是性状表型值中遗传效应的加性效应部分,它剔除了环境的影响,反映遗传效应的大小[12,16]。本研究基于BLUP法进行育种值估算与选择,发现参试家系按表型数据均值进行的排序与按育种值进行排序存在差异,说明环境效应对参试家系表型数据具有较大的影响,相较于以往研究中以表型数据进行的选择,剔除环境效应的选择更为必要,以育种值进行思茅松遗传材料的选择能更好反映参试材料的遗传效应,有利于提高思茅松遗传选择的精确度。此外,研究基于育种值选出的思茅松优良家系,还可用于指导种子园进行选择性采种,在生产用种需求不大情况下,选择性的采种用种能够最大程度的提高遗传增益。
在考虑经济和时间价值的前提下,采伐年龄的 25%~50%常被引用作为最佳选择年龄[27]。基于6 a生半同胞子代测定林生长数据选出的优良家系,可用于经营目标为10-15 a的短周期工业原料林的建设。由于该子测林在后期保存率低,无科学统计的意义,按目前的标准,该子测林暂不能有效支撑优良家系的决选工作。早期选择是缩短林木育种周期的重要手段[28]。在针叶树种方面,基于长期子测林数据相关性分析,火炬松(Pinustaeda)材积最佳的早期选择年龄为6 a[29];赵奋成等[28]认为湿加松杂种(Pinuselliottii×P.caribaea)为选择优良家系作当代利用,最佳选择年龄为3~4 a。速生的松树开展早期选择具有可行性,思茅松同样具有速生特性,6 a生数据是否为可用于开展早期选择,思茅松最佳选择年龄是多少,有待其他子测林长期连续观测数据的支持,也是思茅松长期育种需要进一步完善的方向。
3.2 结论
6 a生思茅松半同胞家系子代测定林,树高、枝下高、胸径和材积性状在不同家系间具有显著的差异,树高、枝下高、胸径和材积4个生长性状受中等以上的遗传控制,单株遗传力分别为0.430、0.270、0.311和0.249,材积性状的遗传变异系数相对较高,适宜作为家系选择的主要性状,在20%的入选率条件下,材积性状仍然可以获得大于20%的遗传增益,基于材积育种值后向选择出4个优良家系,入选的优良家系的材积均值较对照高30.93%~39.07%。
