大豆蛋白组分制备及其功能性的研究进展
2023-08-12栾滨羽黄雨洋夏晓雨朱秀清
栾滨羽,张 波,孟 昂,黄雨洋,夏晓雨,朱秀清,*
(1.哈尔滨商业大学食品工程学院,黑龙江省谷物食品与谷物资源综合加工重点实验室,黑龙江 哈尔滨 150028;2.中国农业科学院农产品加工研究所,农业农村部农产品加工综合性重点实验室,北京 100193)
大豆是重要的植物蛋白源和油料作物,我国进口的转基因大豆主要用于榨油,其豆粕用于制作饲料,而国产非转基因大豆榨油后的豆粕可以制备大豆分离蛋白(soybean protein isolate,SPI)、大豆浓缩蛋白、脱脂大豆粉等进入食品链[1-3]。大豆中蛋白主要存在于籽粒子叶组织的蛋白质体中,约占籽粒干物质总量的40%[4-5],根据其在0.5 mol/L离子强度下的沉降系数可分成2S、7S、11S和15S球蛋白4 种组分[6];也可按功能分成结构蛋白、贮藏蛋白和调节代谢、合成储存物质、创建细胞结构的酶等代谢蛋白[7]。其中7S和11S球蛋白是大豆主要的贮藏蛋白,约占蛋白总量的80%[8]。此外,近年在大豆蛋白分离过程中还发现一种包裹在籽粒油体周围的膜蛋白——亲脂蛋白(lipophilic proteins,LP),该组分是由大豆球蛋白、β-大豆伴球蛋白和油球蛋白等油体结合蛋白与磷脂形成的复合物[9-10]。
大豆蛋白组分结构、功能性等对豆腐、豆浆、植物肉等大豆或大豆蛋白制品的感官品质均有较大影响[11-13]。王喜波等[14]研究发现当大豆蛋白的含硫氨基酸相对含量高于2%、11S与7S球蛋白比例高于1.88、平行式β-折叠结构相对含量高于39.96%时,生产的千叶豆腐品质较好;James等[15]分析了大豆蛋白含量与球蛋白亚基组成对豆腐凝胶性质的影响,发现蛋白含量越高豆腐硬度越大,11SA4亚基的缺失与豆腐硬度和持水性呈正相关;Zhang Qing等[16]对近年来原料品质和加工条件对豆腐品质的影响进行综述,提出豆腐的品质主要与大豆球蛋白和β-大豆伴球蛋白有关,一般大豆球蛋白影响豆腐的硬度,而β-大豆伴球蛋白影响豆腐的弹性,故11S与7S球蛋白比例可视为筛选适宜生产豆腐的大豆品种的一个指标。
本文综述大豆蛋白组分结构的研究现状,重点阐述大豆蛋白组分制备工艺,总结大豆蛋白不同组分的功能性,旨在为大豆或大豆蛋白制品质量控制提供参考。
1 大豆蛋白组分的结构
11S球蛋白是大豆中含量最多的球蛋白,主要为含硫氨基酸较多的大豆球蛋白,分子质量介于320~375 kDa之间,其结构在中性条件下为由5 种亚基构成的空心六聚体[17],每种亚基由一条酸性多肽和一条碱性多肽通过二硫键连接形成,等电点处pH值约为5.8[8,18-19]。7S球蛋白主要由β-大豆伴球蛋白、γ-大豆伴球蛋白和碱性7S球蛋白组成[8]。其中β-大豆伴球蛋白是7S球蛋白的主要组分,分子质量约为180~210 kDa,在中性条件下以三聚体形式存在,由α、α’、β3 种亚基经疏水相互作用聚合形成,α、α’亚基等电点分别为5.2和5.3,而β亚基由等电点在5.8~6.2的4 种组分(β1~β4)构成[5,20-22];β-大豆伴球蛋白结构尤其特殊,表现在α和α’亚基除了核心区外还存在延伸区,且3 种亚基均为N-糖基化,即3 种亚基N末端均与高甘露糖聚糖分子相连,这使它们在溶解性上与大豆球蛋白截然不同[21,23-25]。11S和7S球蛋白的变性温度在不同实验中也有所差别,一般β-大豆伴球蛋白在68~72 ℃变性,大豆球蛋白在86~90 ℃变性[9,26]。大豆籽粒中11S与7S球蛋白的含量比通常在0.5~1.3之间,具体要归因于生长环境和基因型的差异[27-29]。
此外,大豆蛋白中15S球蛋白含量最少,约5%,但凝胶过滤层析分析其分子质量表明15S球蛋白可视为由11S球蛋白亚基构成的二聚体[8,30]。2S球蛋白主要由胰蛋白酶抑制剂等抗营养因子构成,虽然其含量仅为8%,但其可引发人体呼吸道过敏反应,故加工中必须除去[8]。LP中约含蛋白70%、脂质10%,主要由脂肪氧合酶和膜蛋白组成[9,19]。其蛋白根据分子质量可分成来自油体的油质蛋白(24、18 kDa)、源于液泡中的蛋白(34 kDa)以及7S和11S球蛋白中各亚基和多肽等,其中7S和11S球蛋白亚单位约占LP总蛋白含量的60%[9]。LP中的蛋白对考马斯亮蓝染液的敏感性较低,因此通常十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)无法对其蛋白组成进行粗略定量。Samoto等[19]在2008年才通过凯氏定氮法和薄层色谱定量脱脂过程中未被正己烷除去的LP,最终发现7S、11S球蛋白及LP分别占脱脂豆粕总蛋白含量的23%、46%、31%。
2 大豆蛋白组分的制备
目前常见的大豆蛋白商品主要有脱脂豆粉、大豆浓缩蛋白、SPI 3 种。其中SPI的蛋白含量最高,可达90%以上;大豆浓缩蛋白次之,约70%;而脱脂豆粉仅去除了原料中绝大部分脂质,残留约50%的多糖等非蛋白组分,但它是SPI的原料[5],它们均具有完备的工艺流水线生产方案。但7S、11S球蛋白及LP的提取方法还远未达到流水线生产的标准。
Nagano等[31]为测定7S大豆球蛋白凝胶的动态流变参数,首次提出了一个针对7S和11S球蛋白的提纯方案。该方案以脱脂豆粕为原料,将其与蒸馏水混合后,室温静置1 h,筛去不溶性组分后离心;离心后对上清液添加终质量浓度为0.98 g/L的亚硫酸氢钠(sodium bisulphite,SBS),调整pH值至6.4后冰浴过夜储存,再次离心,此时得到的沉淀物为11S球蛋白;上清液再加入0.25 mol/L NaCl溶液并调整pH值至5.0,再次离心;将离心后的上清液与2 倍体积的蒸馏水混合,调整pH值至4.8,再次离心,此时的沉淀为7S球蛋白。这种方法获得的11S和7S球蛋白在脱脂豆粕中的提取率分别为10%和6%,纯度均可达到90%以上。其中0.25 mol/L NaCl、pH 5.0和4 ℃被认为是分离7S球蛋白的最佳条件,而SBS则起到还原剂的作用,从而使蛋白提取率提高[32]。该方法是大豆蛋白“三步酸沉”的基本分离方案。
自1992年起,众多研究致力于优化大豆蛋白的提取效果,主要基于提取过程中温度、pH值和离子强度等工艺参数对蛋白构象的影响[31,33-35]。Deak等[33]曾为简化提取过程采用CaCl2代替NaCl,由于表面电荷密度差异,pH 6.4时钙离子更易与11S球蛋白结合,此方法虽提高了蛋白产量,但所提取的蛋白纯度较低。Liu Chun等[34]选取碱性提取液的种类、碱溶pH值、提取温度、脱脂豆粉与Tris-HCl缓冲液的比例、SBS浓度5 个因素,以11S和7S球蛋白含量、纯度和提取率为指标依次进行单因素优化试验,结果表明,以0.3 mol/L Tris-HCl缓冲液作为提取液可使11S和7S球蛋白的产量分别显著提高2.01%和1.16%;碱溶pH值从7.5升高至9.0的过程中,两种蛋白的产量和纯度均先升高再降低,在pH 8.5时提取效果最好;提取温度是指碱溶温度,在由25 ℃升温至45 ℃的过程中,两种蛋白产量均显著上升,并在45~65 ℃时显著下降,相比Nagano等[31]在室温下提取的方法效果更好,但45~55 ℃并未达到两种蛋白通常的变性温度;增加单位质量豆粉所需缓冲液的质量对增大蛋白溶解度从而提高产率和纯度是最简便有效的方法,但这还需考虑所用容器容量问题;通过SBS破坏二硫键从而降低蛋白聚集度同样可提高蛋白溶解性,但随着SBS浓度的提高,两种蛋白的产量、蛋白含量、纯度均呈先升高后降低趋势,并在SBS浓度为0.01 mol/L时均达到最大值。在此基础上,Din等[36]于2021年注意到前处理条件、原料种类对蛋白提取效果的影响,并将二次酸沉得到的中间产物回收用于提取11S球蛋白;结果表明用植酸酶前处理大豆籽粒、脱脂豆粕和SPI均可使11S和7S球蛋白的纯度显著提高,其中用SPI提取11S球蛋白效果最好,纯度可达97.16%,产率可达48.92%,但酶处理后7S球蛋白产率会显著下降。
实际上SPI中不仅有7S和11S球蛋白,在籽粒蛋白体和油体周围还存在着大量膜蛋白[9]。研究表明通过传统碱溶酸沉方式提取的大豆蛋白味道不佳,还残留部分与脂类结合的蛋白,即LP[32]。受此启发,Samoto等[19]重点探讨了蛋白提取过程中LP的反应,为保证蛋白品质设计了一种新提取方案。该方案以经过70~80 ℃预热处理的低温脱脂豆粕为原料,与Nagano等[31]方法的明显区别在于:选用5 mol/L稀硫酸和氢氧化钠调节pH值;将pH值调整到5.8后提取11S球蛋白,之后调节pH值至5.0,再调至pH 5.5后进行离心提取LP,最后将pH值调至4.8得到7S球蛋白;此方法不需要大量的盐和冷却分离过程。该方法能够提取LP的关键是利用了LP具有强疏水性且易于发生盐析的特性。在pH 5.0时LP与7S球蛋白都处于不可溶状态,pH 5.5时达到7S球蛋白溶解而LP不溶的状态。此外预热处理可保证7S和11S球蛋白的提取效果,但温度过高会降低LP提取率。LP的发现间接解释了以往方法中提取的11S与7S球蛋白不纯且量少的原因,同时也促使一些研究者开始探讨其功能性。
不同研究中大豆蛋白组分的提取流程和参数分别如图1和表1所示。

表1 大豆蛋白提取工艺参数Table 1 Process parameters of soybean protein extraction
3 大豆蛋白组分功能性
加工条件会导致蛋白质由天然状态转换为中间状态,最终彻底变性,蛋白质的功能性也由此变化[37]。在该转变过程中蛋白质分子质量和一级结构通常不变,主要变化体现为蛋白质二级和三级结构改变引起的表层氨基酸组成改变,这为蛋白与蛋白之间以及蛋白与其他组分的相互作用提供条件。故蛋白质的天然状态并不能完全决定其功能性,还需了解溶液、界面和凝胶等环境下蛋白质在折叠与展开过程中的结构转变行为。
3.1 溶解性
蛋白质的溶解性通常是凝胶、乳液、饮料等产品生产所需考察的首要特性,可通过蛋白质在特定溶液中处于可溶状态的氮含量占样品总氮含量的比例表示。溶解性不仅与蛋白本身的氨基酸组成、序列、分子质量、蛋白构象有关,还与离子强度、pH值、温度等环境因素有关[38-39]。根据蛋白结构可推测中性条件下β-大豆伴球蛋白N末端所连多糖会使其在水中的溶解度增大,这一结果已得到部分研究证实[40-41]。Jiang Jiang等[40]采用同种脱脂豆粕分别制备出SPI及11S、7S球蛋白3 种蛋白,将所得样品以20 mg/mL质量浓度分散于pH 7.0、10 mmol/L磷酸钠溶液中,通过调节分散液pH值,发现在pH 7.0时7S球蛋白溶解度最大(90%),其次是SPI(80%)和11S球蛋白(60%)。3 种蛋白的溶解度随pH值的变化曲线均呈现为U型,7S球蛋白和SPI均在pH 4.5左右溶解度最低,11S球蛋白在pH 5.0处达到溶解度最低点,表明11S和7S球蛋白的等电点分别约为4.5和5.0。此外,随着分散液中盐浓度的增大,3 种蛋白的溶解度对pH值的敏感性都降低,在0、0.1、0.6 mol/L NaCl条件下,11S和7S球蛋白的溶解度变化差值分别为82.2%、82.0%、53.7%和93.9%、88.7%、8.2%。这与NaCl的离子效应有关。在等电点(pH 4~5)内,较高浓度的Na+和Cl-与蛋白质表面带电基团相互作用,可在晶体-溶液界面形成双电子层,从而增加等电点处大豆蛋白质的表观溶解度。在等电点外,高浓度的离子会中和蛋白质表面异种电荷,并因此降低了调节溶液的pH值时引起的净电荷增益。
大豆蛋白的溶解性还会受到热聚集行为影响。一些研究者提出了Lumry-Eyring成核聚集模型,该模型中认为蛋白聚集由多个阶段组成,依次包括构象变化、成核前、不可逆的聚集体成核、聚合程度增大和聚集体自缔合[42-44]。Tang Chuanhe等[41]研究表明经过80 ℃预热处理后,SPI仅在97.6 ℃出现11S球蛋白对应的吸热峰,这可能是由于7 S 球蛋白在8 0 ℃下完全变性展开后与11 S 球蛋白形成了更稳定的聚集体。Guo Jian等[21]发现了β-大豆伴球蛋白和大豆球蛋白在加热过程中聚集行为的区别。β-大豆伴球蛋白在由50 ℃加热至100 ℃的过程中溶解度基本不变,而大豆球蛋白的溶解度随温度增加呈下降趋势。当大豆球蛋白与β-大豆伴球蛋白分别按照4∶1、2∶1、1∶1的比例混合时,随着β-大豆伴球蛋白的加入,复合物聚集体更小,溶解度提高。疏水相互作用可能是蛋白聚集的主要驱动力[45]。两种蛋白的热聚集机理如图2所示。当聚集发生时,对于β-大豆伴球蛋白,一旦疏水残基被覆盖并形成聚集体,表面的多糖和亲水基团就会提供排斥力抑制其他蛋白单体接近;但大豆球蛋白的碱性多肽中存在较多疏水氨基酸,因此展开后会暴露出更多活性位点。虽然部分活性位点在聚集过程中被覆盖,但聚集体表面仍存在疏水残基,导致活性位点持续聚集;在两种蛋白共存时,覆盖在球蛋白聚集体表面的不再是疏水残基,而是β-大豆伴球蛋白的亲水基团,从而终止了聚集行为。此外,这种蛋白质之间的热聚集行为还与浓度有关。增加蛋白浓度会减小蛋白间距,有效地促进聚集[46-47]。叶荣飞等[48]研究表明不同浓度的SPI经80、100、120 ℃热处理后的溶解度均随着蛋白浓度的增大而减小。Chen Nannan等[49]认为由于天然大豆蛋白在高浓度下自发结合,导致蛋白质浓度越大,形成的聚集体回转半径越大,蛋白间相互作用越强。
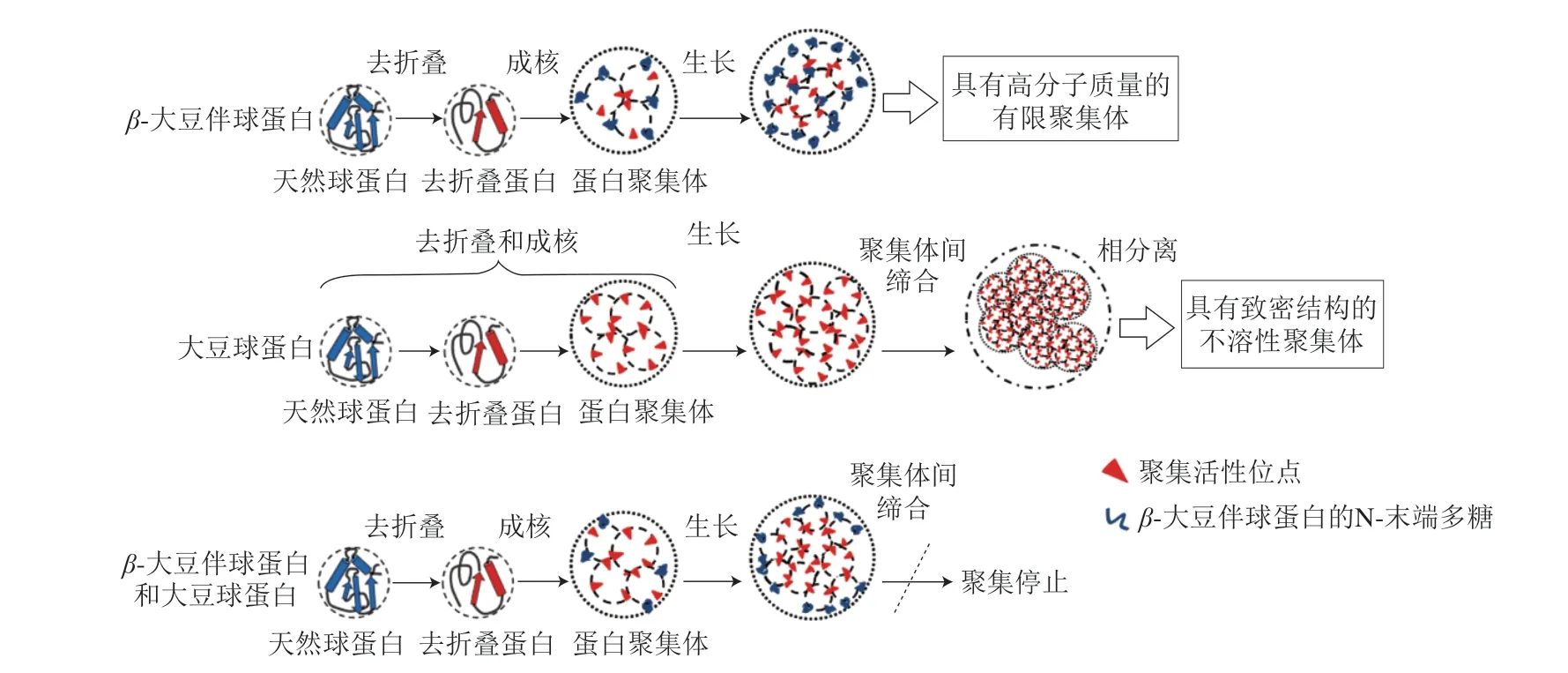
图2 pH 7.0条件下β-大豆伴球蛋白、大豆球蛋白及其混合物的热聚集行为[21]Fig.2 Thermal aggregation behavior of β-conglycinin,glycinin and their mixtures at pH 7.0[21]
育种和蛋白提取工艺的改进为获得纯度较高的某种大豆蛋白提供了更好的方法,Yuan Debao等[50]以经低热处理后的豆粕为原料,通过DEAE-Sepharose快速流动色谱结合固定化金属离子亲和色谱,成功获得了纯度较高的β-大豆伴球蛋白的3 种亚基,其中β亚基含量为91.4%、α亚基含量为95.0%、α′亚基含量为92.1%。在此基础上,He Xiuting等[25]发现7S球蛋白的热聚集主要由β亚基主导,11S球蛋白热聚集由碱性多肽主导。β亚基在50~90 ℃加热过程中分子质量较高(>669 kDa)的组分对应色谱峰的峰面积显著增加,而α及α’亚基的分子质量分布基本不随温度变化而改变;11S球蛋白的部分酸性多肽在加热过程中完全不聚集,碱性多肽在90 ℃加热30 min后即产生大量不溶物。动态光散射结果表明从50 ℃至90 ℃,11S球蛋白和酸性多肽的粒径均变大,两者的Z-平均直径分别由56 nm增至158 nm(11S球蛋白)、由79 nm增至112 nm(酸性多肽);7S球蛋白粒径在加热过程中的增长较小(由29 nm增至44 nm),α及α’亚基的Z-平均直径在不同温度下始终稳定在29 nm左右,但β亚基的Z-平均直径在50 ℃加热时就达到了70 nm,在90 ℃时增大至158 nm,因此7S球蛋白中β亚基更易于聚集。对比不同蛋白亚基及多肽间表面疏水性发现,11S球蛋白的表面疏水性H0在60 ℃时约为5 000,后随温度升高急剧增加,在80 ℃时达到峰值12 000;酸性多肽加热过程中的H0变化较小,在80 ℃同样达到峰值,为4 200。7S球蛋白、β亚基和α及α’亚基在不同温度下加热的H0峰值分别在8 000、7 800和6 000左右波动。此外,蛋白、亚基和多肽的表面疏水性均在加热过程的前10 min内增长迅速。这些结论与Guo Jian等[21]相似,即球蛋白分子先经过去折叠,在变性温度下加热一定时间后球状结构展开,暴露出更多的疏水残基,使表面疏水性增加。这种亚基及多肽间的热聚集行为差异可能与蛋白一级结构氨基酸组成有关,例如含Val、Leu、Ala等疏水氨基酸较多的碱性多肽室温下即可形成不溶性聚集体[51],β亚基同样疏水氨基酸含量较多,且其结构中的延伸区仅有一个N-连接的高甘露糖多糖,但α及α’亚基各含有两个,因此β亚基比α及α’亚基更易于聚集。故大豆球蛋白的溶解性受温度影响较大,β-大豆伴球蛋白溶解性受离子强度影响较大。
3.2 凝胶性
凝胶是一种介于固态与液态之间的物质特殊状态,大多食品都可以凝胶状态食用。当蛋白溶胶的分子间相互作用增加,交联增大至一定程度时转化为凝胶[37]。大豆蛋白热诱导凝胶的基本形成过程如下:大豆蛋白分散在水中,先以卷曲紧密的形式构成溶胶,随温度升高蛋白逐渐变性去折叠,邻近分子间先形成聚集体,同时高温使分子运动加剧,蛋白与蛋白间疏水相互作用更频繁,达到平衡后形成具有一定网络结构的凝胶,部分游离水被截留在凝胶网络之中[52]。该形成过程中,共价相互作用二硫键起主导作用,同时也存在氢键、静电斥力等非共价相互作用[37]。其中11S和7S球蛋白在35 mmol/L磷酸盐缓冲液中(pH 7.6),100 ℃下形成凝胶的临界蛋白含量分别为2.5%和7.5%[53]。11S球蛋白热凝胶主要通过二硫键和静电相互作用形成,7S球蛋白热凝胶则通过氢键形成[16]。故pH值和温度同样可通过影响蛋白质聚集体的方式影响凝胶的结构和性质[54-55]。
根据大豆球蛋白加热后聚集度更高可推测其热诱导凝胶的储能模量(G’)也应相对较大。Renkema等[56]证实了这一结论。同种大豆制备的大豆球蛋白(纯度95%)和β-大豆伴球蛋白(纯度60%)热诱导凝胶的温度扫描结果表明,在pH 3.8下大豆球蛋白和β-大豆伴球蛋白凝胶冷却结束时的G’分别为4 000 Pa和2 500 Pa,pH 7.6下前者的G’为7 400 Pa,后者为5 400 Pa。pH 3.8和pH 7.6的大豆球蛋白凝胶的断裂应力也显著大于β-大豆伴球蛋白凝胶,前者分别为46.2 kPa和18.1 kPa,后者分别为2.1 kPa和2.2 kPa。
这两种蛋白凝胶G’的差异也可能是由于含硫氨基酸含量不同,11S球蛋白每单位蛋白的蛋氨酸和半胱氨酸含量是7S球蛋白的3~4 倍[57],因此11S球蛋白在凝胶形成过程中有更强的交联网络。Bainy等[58]的研究结果也表明热诱导凝胶的G’与11S和7S球蛋白比例并不直接相关(r<0.50),还取决于一种半胱氨酸残基含量较少的A3酸性亚基的含量。由11 种不同大豆球蛋白、β-大豆伴球蛋白亚基缺失型大豆和1 种正常蛋白组成大豆制成的SPI热诱导凝胶的动态流变分析结果表明,当A3亚基占总蛋白含量小于2%时,11S球蛋白亚基含量与凝胶完全形成时的G’存在相关性(r>0.967);当A3亚基占总蛋白含量大于2%时,凝胶完全形成时的G’与大豆蛋白组成无相关性,且11S球蛋白含量高的品种与11S球蛋白含量低的品种凝胶G’间无显著差异,即7S球蛋白是热诱导凝胶的主要结构单体,这可能是90 ℃下11S球蛋白不足以完全展开造成的。在针对豆腐的研究中也有相似结论,即11S球蛋白亚基组成与凝胶硬度间存在相关性[15,59]。此外,由于蛋白去折叠是凝胶形成的先决条件,故可以预测热转变温度较低的β-大豆伴球蛋白凝胶形成时(损耗模量(G’)/G’=1)的温度低于大豆球蛋白。7S球蛋白含量较高的品种凝胶形成温度为74.2~82.2 ℃,11S球蛋白含量较高的品种凝胶形成温度则介于86.2~90.2 ℃之间。
3.3 流变学特性
流变性测试可通过施加较大的剪切力或剪切频率来破坏物料结构,从而在一定程度上反映原料在加工过程中的相变,具体通过黏度、G’、G’’以及扭矩等参数表征[60-62]。例如蛋白黏度过高往往会在加工中形成结块无法溶解。物料流变性变化既受工艺参数的影响也受物料本身组成的影响。
Sandoval等[60]的研究表明对于高水分挤压大豆蛋白产品,由黏度不同导致的速度梯度是其纤维状结构形成的关键。Wittek等[63]对来源和蛋白质含量基本相同(占干物质总量的90%以上)的4 种SPI凝胶的稳态扫描结果表明,相同条件下,处理60 s时SPI凝胶的复合黏度分别为44 kPa·s(SPI 1)、43 kPa·s(SPI 2)、34 kPa·s(SPI 3)和14 kPa·s(SPI 4),这种差异可归因于提取工艺不同导致的变性程度不一[64]。在Wittek等[63]的研究中,大豆浓缩蛋白的蛋白含量约占干物质总量的67%,但相同条件下其黏度(101 kPa·s)为SPI黏度的2 倍以上。这表明原料中较高比例的多糖等其他成分对黏度也有影响,与Zhang Wei等[65]对挤压植物蛋白的研究结果相似,SPI的变性焓(ΔH)为0.72 J/g,将SPI、谷朊粉分别与小麦淀粉、玉米淀粉、马铃薯淀粉、甘薯淀粉、木薯淀粉、绿豆淀粉、豌豆淀粉、马铃薯直链淀粉、玉米支链淀粉共9 种淀粉混合(m(SPI)∶m(谷朊粉)∶m(淀粉)=65∶15∶20)后,混合物的ΔH介于1.24~2.42 J/g之间,混合物挤压时的黏度介于308.70~633.61 Pa·s之间,混合物的ΔH与黏度呈显著正相关(P<0.05)。
大多研究从蛋白质组分理化性质的角度讨论其对凝胶结构形成的影响,但这不足以解释凝胶加工过程的变化行为[66-69]。Mellema等[70]提出由展开的蛋白互相连接形成的链的曲率和连接方式决定着凝胶将以何种方式在微观水平上变形,例如具有弧形链的凝胶主要变形方式为弯曲,而直线型且互相连接的链以拉伸的方式变形。在此基础上,Renkama[71]的研究表明蛋白质链越弯曲,凝胶断裂时的应变越大,因为弯曲的链要先伸直再发生断裂。Xia Wenjie等[72]通过大振幅振荡剪切实验发现SPI、富11S蛋白(11S球蛋白含量72.1%、7S球蛋白含量3.3%)和富7S蛋白(7S球蛋白含量30.4%、11S球蛋白含量4.2%)三者形成热凝胶的固有黏度和分子间相互作用强度不同。低蛋白含量(6%)条件下,三者G’排序依次为富11S蛋白(600 Pa)>SPI(300 Pa)>富7S蛋白(60 Pa),较低蛋白含量时的G’可反映分子间不发生或较少发生碰撞、聚合等相互作用时分散相的固有黏度,故富11S蛋白的固有黏度较高,SPI次之。但富7S蛋白的分子间相互作用更高,这可通过3 种蛋白G’-蛋白浓度曲线体现,富7S蛋白对应曲线的斜率最大(281.25),其次是SPI(266.67),富11S蛋白最小(150),即每增加单位浓度蛋白,富7S蛋白分子碰撞、聚集程度更大,G’增长更快。这可能是由于大豆球蛋白展开不充分,刚性结构的球蛋白颗粒抑制了分子间交联[73-74]。对于富7S蛋白凝胶,可能是由于β-大豆伴球蛋白的柔性较强,形成的颗粒分支较多,相互间的联系也更紧密,随着应变增加,最初解离的颗粒会互相缠绕形成新团簇[75]。Gisler等[76]利用共聚焦激光扫描显微镜和扫描电子显微镜进行观察发现,SPI和富11S凝胶具有相对致密的网络,由较大的聚集体构成,而富7S蛋白凝胶更粗糙和不均匀,其组成的颗粒较小且分支较多。
4 结语
大豆虽在我国食品生产中有较多应用,但由于其组分和提取工艺的多样性,企业对于产品品质的调控仍多凭借经验选择,原料组成、结构、功能性与感官品质之间的相关性仍比较模糊。本文介绍了近年来有关大豆蛋白组成及工业生产的相关知识,主要侧重于蛋白提取工艺和功能性方面。有关大豆各组分的分离鉴定和蛋白功能性的表征手段仍留有空白。此外,为了达到绿色生产、减少碳排放的目的,应该鼓励豆渣、豆粕等副产品废弃物的利用,例如采用挤压蒸煮生产素肉产品、开发大豆活性肽、生产生物基塑料制品等。
