外源一氧化氮对中度干旱胁迫下越橘幼苗生理生化特性的影响
2023-08-11王明月陈明唐雪东刘炳含周思佳陈国双
王明月,陈明,唐雪东,刘炳含,周思佳,陈国双
(1.吉林农业大学园艺学院,吉林 长春 130118;2.中国科学院东北地理与农业生态研究所,吉林 长春 130118)
越橘为杜鹃花科(Ericaceae)越橘属(Vaccinium)多年生灌木果树,因具有较高的食用及药用价值[1-3]在我国快速发展,栽培面积不断扩大[4]。越橘根系较浅,主要分布在0 ~20 cm 土层,且主根不明显、无根毛,不能更好地吸收土壤深层水分,导致其不能有效抵御干旱胁迫伤害[5],在我国的种植推广及发展受到严重影响。
近年来,外源物质在提高植物抗逆性等方面受到广泛关注。 NO 作为一种重要的外源信号分子,用于保护植物免受生物、非生物胁迫诱导的损伤[6-8]。 NO 具有双重性作用,即低浓度促进、高浓度抑制[9,10]。 沈宏伟等[11]研究发现,刺五加种子的酶活力在施用一定浓度的硝普钠(SNP)下可以显著增加,且发芽率提高,种子发芽时间缩短。 张文博等[12]研究表明,适宜浓度的外源NO 可以保护低温胁迫下黄瓜叶片的质膜透性,抑制MDA 积累,提高其耐寒性。 杨玉花等[13]发现,一定浓度的外源NO 可以促进大豆种子萌发,降低大豆根尖活性氧含量,提高SOD、POD 等抗氧化酶活性,以降低镉胁迫对大豆造成的不利影响;Azizi 等[14]在夏季香薄荷的研究中也得到相同结果。 徐严等[15]研究显示,外源NO 可以缓解越橘叶片受干旱胁迫的伤害,并且与外源NO 的浓度显著相关。 王延秀等[16]对楸子的研究也有相似结果,即适宜浓度外源NO 处理可以提高其对干旱胁迫的忍耐性。 喷施多效唑[17]、H2S[18]、Si[19]、褪黑素[20]和ABA[21]对提高果树的抗旱性有显著效果。
本试验以二年生越橘‘蓝丰’为材料,研究喷施不同浓度外源NO 对中度干旱胁迫下越橘幼苗叶片渗透调节物质、抗氧化物质、抗氧化酶活性的影响,并通过各项指标的主成分分析探讨外源NO 对越橘抗旱性的保护作用,为外源NO 提高越橘抗旱性的生产实践提供理论参考。
1 材料与方法
1.1 材料
本试验选择二年生北高丛越橘品种‘蓝丰’的健壮、树势一致、生长结果良好且无病虫害、无机械损伤的植株为材料。 NO 供体:硝普钠(SNP)。
1.2 试验设计及方法
于2021 年5 月4 日,将越橘植株转移到口径24 cm、底径18 cm、高度20 cm 的塑料盆中单株定植,置于试验地(吉林农业大学东山温室)常规管理,预培养至叶片完全展开后开始试验(7 月21日)。 试验采用持续称重法控制土壤含水量[22]。首先确定盆栽越橘最大持水量(饱和含水量),每隔3 d 傍晚称重并保持水分至土壤最大持水量的45%~55%即中度干旱胁迫[23-25]。 将正常供水(土壤最大持水量的70%~80%)及中度干旱胁迫(土壤最大持水量的45%~55%)下健康均一的越橘苗分成7 个处理组(表1),每处理20 盆,重复3次,即每处理组60 盆,每隔3 d 喷施1 次SNP,共4 次。

表1 试验设计
1.3 测定项目及方法
干旱胁迫处理0、3、6、9、12 d 分别取样,参考张治安等[26]的方法测定各项生理指标。
1.4 数据处理与分析
利用Microsoft Excel 2016 进行数据处理及作图,采用SPSS 25.0 软件进行数据显著性分析和主成分分析[27]。
2 结果与分析
2.1 不同浓度外源NO 对中度干旱胁迫下越橘叶片渗透调节物质含量的影响
2.1.1 对可溶性糖(SS)含量的影响 由图1 可知,随着干旱胁迫天数的增加,CK2 与外源NO 处理组越橘幼苗叶片的可溶性糖含量均表现为逐渐增加趋势,胁迫后3、6 d 均显著高于CK1,并且外源NO 处理组具有明显的浓度效应。 胁迫9 d,各处理组的可溶性糖含量达到最大值,T4 处理最高(1.88%)且显著高于T5 处理外的其余处理,相比于CK1、CK2 其可溶性糖含量分别增加0.68、0.55个百分点。 表明干旱胁迫可以增加叶片的可溶性糖积累,外源NO 处理也会促进干旱胁迫下可溶性糖积累,从而提高越橘叶片的渗透保护能力,提高抗旱性。

图1 外源NO 对中度干旱胁迫下越橘叶片可溶性糖含量的影响
2.1.2 对可溶性蛋白(SP)含量的影响 由图2可知,随着干旱胁迫时间延长,越橘叶片可溶性蛋白含量多呈现先增加后下降趋势,胁迫9 d 时达到最高。 CK1 可溶性蛋白含量始终显著低于CK2及外源NO 处理组,T4 处理的可溶性蛋白含量在胁迫3、6、9 d 时均显著高于其它处理。 胁迫9 d,T4 的可溶性蛋白含量达到最大(1.72 mg/g),与CK1、CK2、T1、T2、T3、T5 相比,分别增加120.51%、68.63%、32.31%、22.86%、18.62%、17.81%。 表明干旱胁迫可促进可溶性蛋白在越橘叶片中积累,外源NO 处理则增加干旱胁迫下可溶性蛋白积累,从而缓解干旱胁迫对叶片的伤害。

图2 外源NO 对中度干旱胁迫下越橘叶片可溶性蛋白含量的影响
2.1.3 对脯氨酸(Pro)含量的影响 由图3 可知,随着干旱胁迫时间延长,越橘叶片的Pro 含量始终呈现增长趋势,胁迫12 d 时达到最高。 CK1的Pro 含量始终显著低于CK2 及外源NO 处理组。 T4 处理的Pro 含量各胁迫时间点均最高,胁迫9、12 d 显著高于其它处理,对于缓解干旱胁迫的效果最好。 胁迫12 d,CK2、T1、T2、T3、T4、T5的Pro 含量与CK1 相比分别增加13.43%、36.26%、42.17%、79.40%、104.73%、71.57%。 说明外源NO 可以通过提高越橘叶片的Pro 含量而达到维持细胞渗调平衡的效果。

图3 外源NO 对中度干旱胁迫下越橘叶片脯氨酸含量的影响
2.2 不同浓度外源NO 对中度干旱胁迫下越橘叶片抗氧化系统的影响
2.2.1 对丙二醛(MDA)含量的影响 由图4 可知,随着干旱胁迫时间延长,各干旱处理越橘叶片的MDA 含量始终显著高于CK1。 干旱胁迫6 ~12 d,CK2 的MDA 含量显著高于不同浓度外源NO 处理,其中T4 处理含量最低。 胁迫9 d 时,CK2 的MDA 含量显著高于CK1,增加60.62%;T1、T2、T3、T4、T5 处理的MDA 含量与CK1 相比分别增加50.44%、48.15%、48.6%、44.01%、46.45%,均达显著水平,与CK2 相比分别下降6.33%、7.76%、7.48%、10.34%、8.82%,也均达显著水平。 表明外源NO 可以缓解干旱胁迫对越橘幼苗叶片膜脂过氧化的伤害。

图4 外源NO 对中度干旱胁迫下越橘叶片丙二醛含量的影响
2.2.2 对超氧阴离子(O2·-)产生速率的影响 由图5 可知,干旱胁迫显著增加越橘叶片的O2·-产生速率。 随着胁迫时间延长,各干旱胁迫处理的O2·-产生速率均不断提高,并且显著高于CK1。CK2 的O2·-产生速率比T1、T2、T3、T4、T5 处理更高,其中T4 处理最低。 干旱胁迫9 d 时,与CK1相比,CK2、T1、T2、T3、T4、T5 处理的O2·-产生速率分别增加191.67%、152.78%、130.56%、116.67%、77.78%、119.44%,均达显著水平;与CK2 相比,T1、T2、T3、T4、T5 处理的O2·-产生速率分别下降13.33%、20.95%、25.71%、39.05%、24.76%,均达显著水平。 表明外源NO 可以缓解干旱胁迫对越橘幼苗叶片膜脂的过氧化作用。

图5 外源NO 对中度干旱胁迫下越橘叶片超氧阴离子产生速率的影响
2.2.3 对过氧化氢含量(H2O2)的影响 由图6可知,随着干旱胁迫时间延长,干旱胁迫处理组(CK2、T1、T2、T3、T4、T5)越橘叶片的H2O2含量均呈上升趋势,且均显著高于CK1,而CK1 的变化不明显。 CK2 的H2O2含量始终高于不同浓度外源NO 处理组,其中T4 处理含量最低。 胁迫9 d 时,CK2 的H2O2含量显著高于CK1,增加195.40%;与CK1 相比,T1、T2、T3、T4、T5 处理的H2O2含量分别增加123.68%、108.11%、101.97%、83.55%、101.54%,均达显著水平;与CK2 相比,T1、T2、T3、T4、T5 处 理 的H2O2含 量 分 别 显 著 下 降24.28%、29.55%、31.63%、37.87%、31.77%。 表明外源NO 能有效降低干旱胁迫下H2O2在越橘幼苗叶片中的积累。
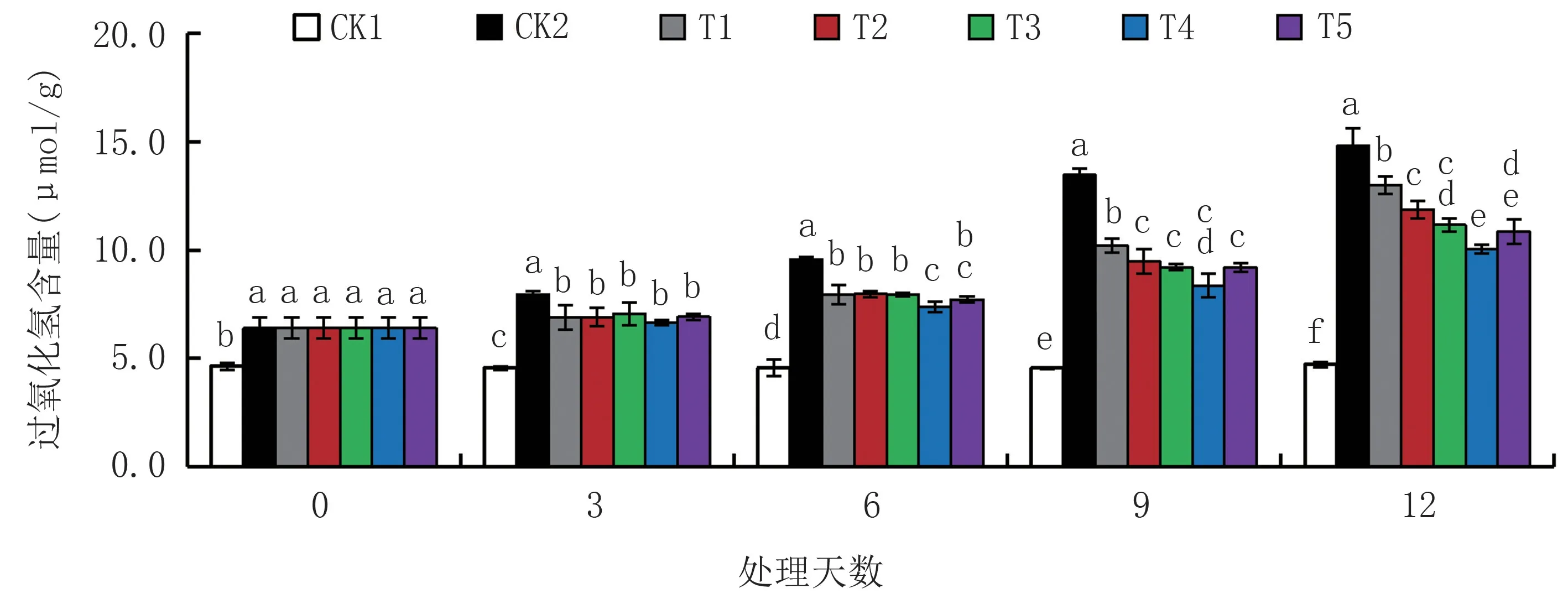
图6 外源NO 对中度干旱胁迫下越橘叶片过氧化氢含量的影响
2.3 不同浓度外源NO 对中度干旱胁迫下越橘叶片抗氧化酶活性的影响
2.3.1 对超氧化物歧化酶(SOD)活性的影响由图7 可知,越橘叶片的SOD 活性随着干旱胁迫时间延长呈现先升高后降低趋势,胁迫9 d 达到最高。 CK1 的SOD 活性始终显著低于其它干旱处理组,T4 处理的SOD 活性在胁迫3 ~12 d 均显著高于其它处理。 胁迫9 d 时,T4 的SOD 活性与CK1、CK2、T1、T2、T3、T5 相比分别增加108.41%、21.77%、17.09%、12.43%、8.60%、10.11%。 表明外源NO 可以在一定程度上提高干旱胁迫下越橘叶片的SOD 活性,以T4 处理效果最佳。

图7 外源NO 对中度干旱胁迫下越橘叶片超氧化物歧化酶活性的影响
2.3.2 对过氧化物酶(POD)活性的影响 由图8可知,随着干旱胁迫时间延长,越橘叶片的POD活性呈现先增加后降低趋势,9 d 时达到最高。各个干旱处理组的POD 活性一直显著高于CK1,T4 处理除0 d 外其它胁迫时间点均显著高于其它处理组。 胁迫9 d 时,与CK1 相比,CK2、T1、T2、T3、T4、T5 分别增加81.62%、87.90%、101.28%、115.36%、132.85%、111.49%;与T4 相比,CK1、CK2、T1、T2、T3、T5 处理分别降低57.05%、22.00%、19.30%、13.56%、7.51%、9.18%。

图8 外源NO 对中度干旱胁迫下越橘叶片过氧化物酶活性的影响
2.4 中度干旱胁迫下不同浓度外源NO 处理对越橘抗旱能力的综合评价
运用SPSS 25.0 软件进行主成分分析,通过计算不同浓度外源NO 处理下越橘幼苗各生理生化指标的贡献值,对不同处理越橘的抗旱性进行综合得分排序,得其抗旱能力大小。 由表2 可知,2 个主成分的特征值分别为5.611 和2.248,均大于1。 第1 主成分贡献率为70.143%,第2 主成分贡献率为28.104%,其方差累积贡献率达到98.247%,大于85%,可用于分析。

表2 越橘叶片各项生理指标主成分特征值及方差贡献率
由表3 可知,第1 主成分中SOD、POD、SP 系数较大,第2 主成分中H2O2、O2·-和MDA 系数均为正向较大,因此,选用SOD、POD、SP、H2O2、O2·-及MDA 作为反映干旱胁迫后外源NO 处理对越橘幼苗影响的重要指标。 用其对不同浓度外源NO 处理下越橘的抗旱性进行综合评价,结果(表4)为T4>T5>T3>T2>CK1>T1>CK2。

表3 越橘叶片各项生理指标主成分分析
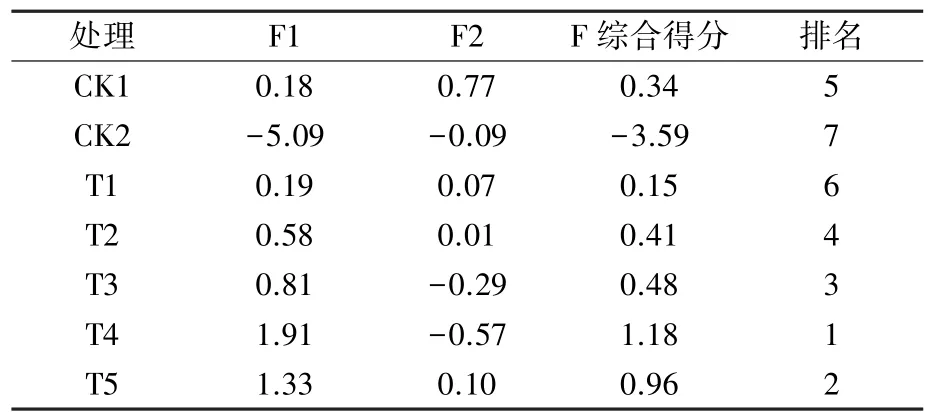
表4 不同浓度外源NO 处理越橘幼苗抗旱能力的综合评价
3 讨论
干旱胁迫时,植物启动抗氧化防御系统清除活性氧、产生逆境蛋白、积累渗透调节物质,以此保持细胞渗透势、维持细胞透性、提高细胞的抗氧化能力和保水能力, 以保证正常的生理活动[28-30]。 本试验中,总体来说,越橘叶片的Pro、SS、SP 含量随着干旱胁迫时间延长而增加,外源NO 处理能促进中度干旱胁迫下这些物质含量的积累,提高越橘叶片细胞膜的渗透保护能力,缓解干旱胁迫对越橘的伤害。 这与郝岗平等[31]在银杏和谢惠敏等[32]在核桃上的研究结果相似。
植物在遭受干旱等非生物胁迫时会积累大量的活性氧,加速体内O2·-的产生速率,加快细胞膜脂过氧化过程,进而增加MDA 含量,诱导SOD 活性增强,继而生成大量H2O2[33],使植物细胞膜结构遭到严重破坏。 SOD、POD 等抗氧化酶可以维持膜结构的完整性并且降低活性氧自由基对膜的攻击[34]。 本试验中,中度干旱胁迫下越橘叶片的SOD、POD 活性持续上升,说明抗氧化酶系统积极响应干旱胁迫对越橘叶片的伤害。 这与前人分别在兔眼蓝莓、葡萄、无花果、草莓干旱胁迫研究中的结果一致[35-38]。 本试验结果还显示,干旱胁迫下MDA 含量持续上升,进而越橘叶片的H2O2含量快速大量积累,O2·-产生速率快速上升,导致越橘叶片膜脂过氧化,膜系统遭到破坏。 这与岑晓斐[39]、姜晓丹[40]、Rigui[30]等在黑果腺肋花楸、果桑和黑麦草干旱胁迫研究的结果一致。 外源NO处理可以显著减缓MDA 含量、H2O2含量、O2·-产生速率,表明外源NO 直接参与植物的生理过程[41],抑制膜脂过氧化,维持细胞膜透性,减轻干旱胁迫的伤害[15,42],从而提高越橘的抗旱性。
许多研究表明,多种生理因素共同影响着植物的抗旱性。 本试验通过对越橘各生理生化指标的主成分分析以及抗旱能力的综合评价得出,SOD、POD、SP、H2O2、O2·-及MDA 是反映干旱胁迫后外源NO 处理对越橘幼苗影响的重要指标,同时得出不同浓度外源NO(SNP 浓度,μmol/L)处理的抗旱性排序为T4(400)>T5(500)>T3(300)>T2(200)>T1(100)。
4 结论
施加外源NO 可以有效缓解中度干旱胁迫下越橘生长受到的伤害,提高抗氧化酶活性,降低活性氧和自由基的积累,促进渗透调节物质含量增加,从而提高越橘抗旱性,并且具有明显的浓度效应,以外施400 μmol/L SNP 对越橘缓解中度干旱胁迫伤害的效果最好。
