‘阳光玫瑰’葡萄不同节位冬芽激素含量差异分析
2023-07-07李志成田淑芬王荣王超霞李洋穆丁郁
李志成,田淑芬,王荣,王超霞,李洋,穆丁郁
(天津农学院/天津市葡萄遗传与育种重点实验室,天津 300384)
‘阳光玫瑰’葡萄具有甜度高、香味浓、抗病性强等优势,现已成为最受欢迎的葡萄品种之一。在我国多地均有种植[1-3],目前已成为鲜食葡萄的主栽品种[4]。部分产区由于栽培措施不当,导致品质下降和严重减产减收[5-6],因此提升‘阳光玫瑰’葡萄花芽分化质量成为亟待解决的问题。
植物的花芽分化通常受植物内部因子和外界环境因素共同调控。激素作为活性信号分子,在植物的整个发育过程中都起着非常重要的作用。吲哚乙酸(IAA)、赤霉素(GA3)、细胞分裂素(CTK)、脱落酸(ABA)作为4种重要的植物内源激素,在葡萄成花的生理活动中呈动态变化,不同激素响应外界信号刺激,单独或协同作用于植物的特定部位,通过不同的信号通路控制开花基因(SOC1、LFY、FLC等)的表达,从而起到调节植物成花的作用[7]。Luckwill在1970年提出激素平衡假说[8],CTK促进成花,GA抑制成花,CTK/GAS的比值与成花能力呈正相关,即该值高时促进形成大量花芽。也有学者从ABA/GA、(IAA+GA3+CTK)/ABA等激素比值反映植物成花趋势[9-11]。由于营养水平、激素供应或外界环境因子等的差异,植株不同部位形成的芽体存在明显的异质性,同一结果母枝不同节位的冬芽发育并不完全一致[12]。众多研究发现,不同节位冬芽内源激素含量差异明显,花芽分化时间存在差异[13-15]。但也有研究发现,虽然葡萄不同节位花芽间发育进程有差异,但最终会趋向一致[16]。
本研究从植物内源激素层面分析天津地区‘阳光玫瑰’葡萄不同节位间花芽分化的差异,探究4种激素与葡萄花芽分化的相关性,为实际生产提供一定的理论依据和数据支撑。
1 材料与方法
1.1 试验材料
试验于天津市葡萄遗传育种重点实验室进行。供试材料为日光温室栽培的9年生‘阳光玫瑰’葡萄,株距1.5 m,棚架栽培。试验园位于39°27′N,117°07′E,属暖温带半湿润大陆性季风气候,年均气温12.1 ℃,日照时数2730 h,降水584.9 mm,年均无霜期216 d。
1.2 试验设计及样品处理
试验于2022年6月6日至10月5日进行。选择树势良好的‘阳光玫瑰’葡萄植株,对保留7个以上冬芽节位的枝条,每15 d左右取样一次,每次采集20组枝条,共采集9次。带回实验室,用刀片切下完整的冬芽,10组样品用于徒手切片,10组样品用于激素测定。用于激素测定的冬芽样品在处理后快速放入液氮中保存。
1.3 冬芽切片观察
对各时期不同节位冬芽进行纵切,置于体式解剖镜下观察,并拍照。
1.4 花芽率的测定
在切片中,将芽体饱满、鳞片状托叶顶部较为圆润、或能直接观察出花序原基结构的记为花芽;芽体不饱满、芽内鳞片状托叶顶部较尖、无明显花序结构记为叶芽。统计不同时期内花芽叶芽的数量,使用公式计算。
花芽率(%)=(花芽数量/总芽数)×100
1.5 激素含量的测定
本试验采用酶联免疫分析(ELISA)试剂盒测定植物内源激素IAA、ABA、CTK、GA3含量。
将葡萄冬芽从液氮中取出,放入预冷的研钵中快速研磨,称取0.05 g粉末至预冷的离心管中,加入0.45 mL PBS缓冲液(pH 7.4),涡旋30 s,保证样品充分混匀。以3000 r·min-1离心30 min,仔细收集上清液,分装,进行不同激素含量的测定。试验方法参照试剂盒说明书进行。每个样品3次重复。
酶联免疫试剂盒由武汉纯度生物公司提供,使用Spectra Max M5酶标仪在波长450 nm下测定标准样品及待测样品吸光度值,制作标准曲线,计算待测样品激素含量。
1.6 数据分析
采用SPSS 26软件进行方差分析(P<0.05),Microsoft Excel 2019软件进行数据分析和图表制作。
2 结果与分析
2.1 冬芽形态观察
通过对冬芽的切片观察发现,葡萄冬芽随着物候期的变化不断成熟。图1可知,在6月6—21日,冬芽整体呈现嫩绿色,芽体内各部分分化程度较浅;在7月4日,着生在鳞片状托叶上呈绒毛状的毛被较为明显,芽体内呈现红褐色;随着冬芽生长发育,毛被逐渐增多,在生长后期,葡萄冬芽内花序原基逐渐发育变大。在10月5日时期,冬芽逐渐进入休眠。
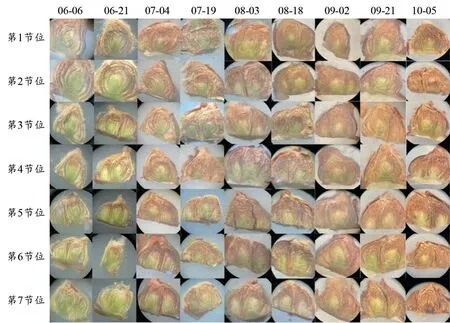
图1 不同时期葡萄冬芽的形态Figure 1 Morphology of winter buds of grapevine at different times
同一时期,不同节位间芽存在明显的异质性。由图1可以看出,第2~6节位冬芽发育程度高,芽体饱满,且发育较快;第1节位发育程度低,且发育慢;第7节位冬芽发育程度较好,但是在发育后期,出现主芽坏死的情况。
2.2 花芽率调查
葡萄冬芽花芽率整体随着生长发育逐渐升高,多数节位在6月21日—7月19日花芽率大幅升高(图2)。在7月19日,第3节位花芽率最先达到100%,第2节位在8月18日达到100%,但第1节位在9月2日花芽率稳定在80%,第7节位花芽率在10月5日降低。
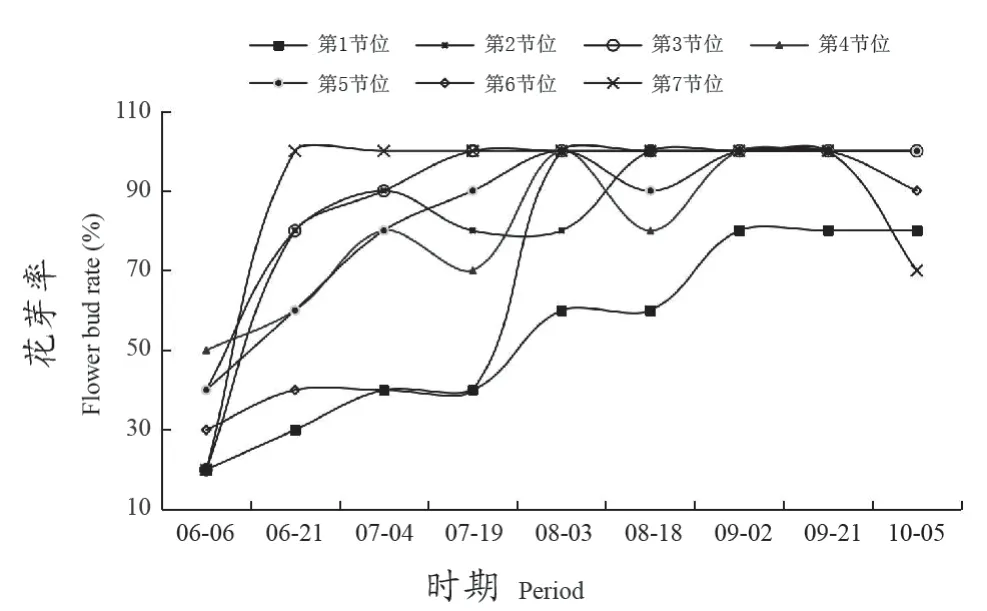
图2 不同节位各时期花芽率Figure 2 Flower bud rate at different nodes and stages
2.3 不同节位冬芽中IAA含量的动态变化
如图3所示,从6月6日—10月5日,‘阳光玫瑰’葡萄冬芽中IAA含量在63.85~137.119 mmol·g-1范围内变化。不同时期,第1~7节位冬芽中IAA含量呈下降-升高-下降-升高-下降的变化趋势。在6月6—21日,IAA含量下降,结合花芽分化情况,此时的第1、4、5、6节位冬芽花芽分化不活跃,花芽率增长缓慢。在7月4日,IAA含量出现一个峰值,在6月21日至7月19日期间,第2、3、4、5、7节位冬芽分化迅速,花芽率迅速提高;在7月19日至9月2日,各节位冬芽IAA含量逐渐升高,第1、2、6、7节位冬芽花芽率也随之升高。
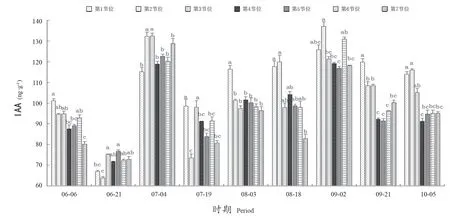
图3 不同节位各时期IAA含量Figure 3 IAA content at different nodes and periods
在6月21日,第2节位冬芽中IAA含量最低,显著低于其他6个节位。7月4日,第2节位和第3节位冬芽中IAA含量较高;8月3日,第1节位冬芽IAA含量显著高于其他节位。
2.4 不同节位冬芽中ABA含量的动态变化
如图4所示,从6月6日至10月5日,‘阳光玫瑰’葡萄冬芽中ABA含量在325.87~593.01 ng·g-1范围内变化。不同时期,第1~7节位冬芽中ABA含量呈升高-下降-升高-下降的变化趋势。整个时期中,葡萄冬芽在6月6日ABA含量最低;在6月6日—7月19日,ABA含量升高,此时期第2、3、4、5、7节位冬芽花芽率升高;各节位冬芽在9月2日时ABA含量达到最大值,在525.44~593.01 ng·g-1,第2、4节位在此时期花芽率也达到100%。随着果实的进一步成熟,各节位ABA含量逐步降低。
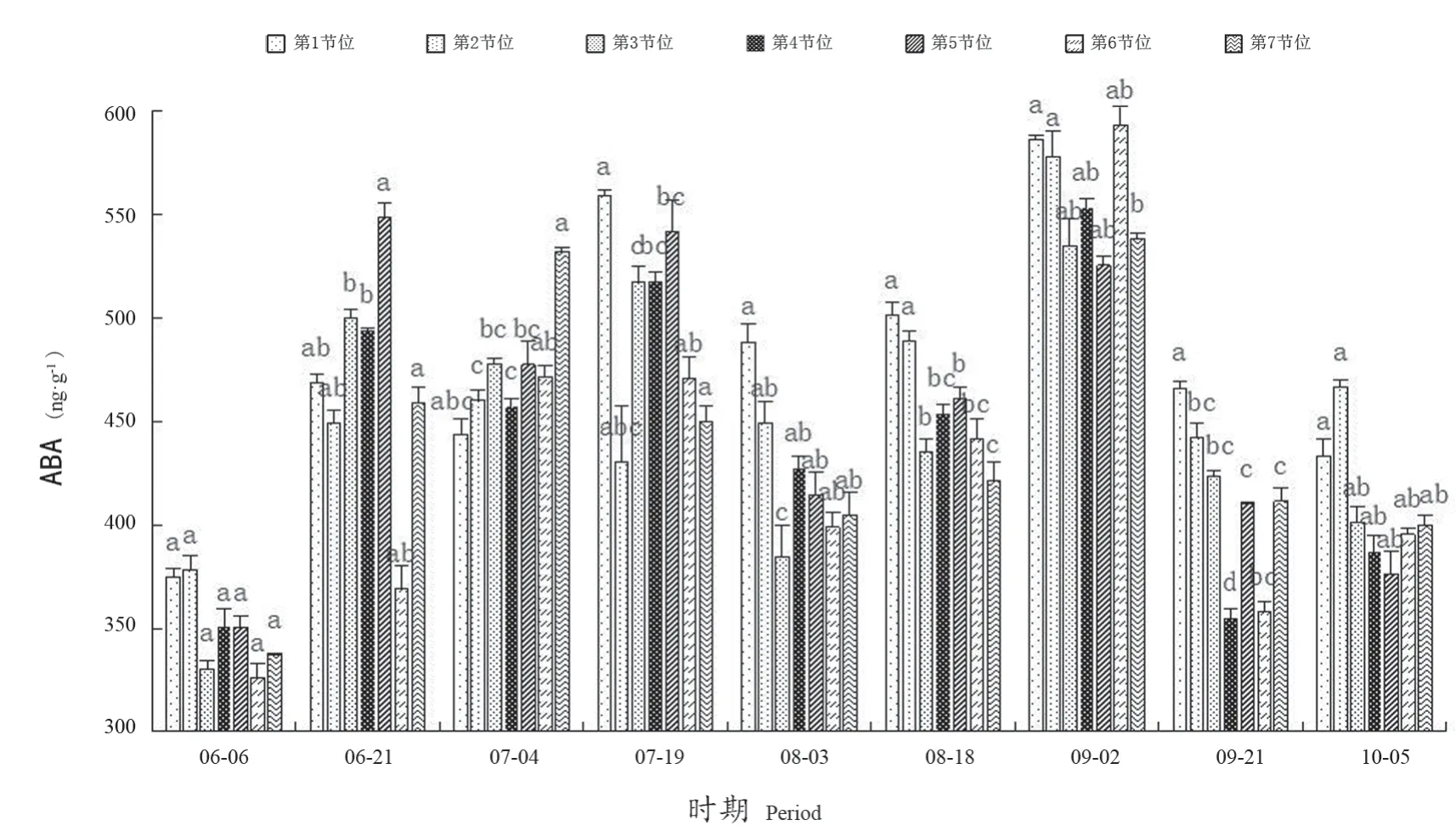
图4 不同节位各时期ABA含量Figure 4 ABA content at different nodes and periods
整个生理分化时期中,在6月6日,各节位冬芽间ABA含量差异不显著;在9月2日,第6节位ABA含量最高,第5节位最低。随着果实进一步成熟,到9月21日第1节位冬芽ABA含量最高,为466.22 ng·g-1,显著高于第4~7节位冬芽。
2.5 不同节位冬芽中CTK含量的时期变化
如图5所示,从6月6日至10月5日,‘阳光玫瑰’枝条冬芽中CTK的含量在162.33~280.86 ng·g-1范围内变化。不同时期,第1~7节位冬芽中CTK含量呈下降-升高-下降的变化趋势。6月6日—7月19日葡萄冬芽中CTK含量低于8月3日—10月5日。在8月3日,第1~7节位冬芽CTK含量迅速升高,在9月2日达到最大值。
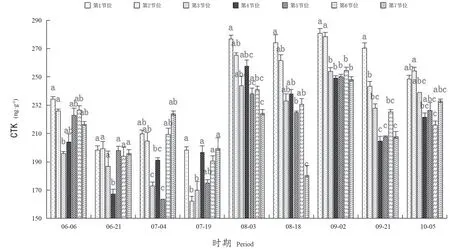
图5 不同节位各时期CTK含量Figure 5 CTK content at different nodes and periods
在整个生理分化时期中,6月21日—7月19日,第1~7节位冬芽CTK含量降低,不同节位间差异不显著。在9月2日,第1~7节位冬芽CTK含量最高,其中,第1、2节位含量较高,分别为280.86、278.33 ng·g-1,显著高于其他5个节位。在9月21日,第1节位冬芽CTK含量最高,为270.32 ng·g-1,显著高于其他节位。
2.6 不同节位冬芽中GA3含量的动态变化
如图6所示,从6月6日至10月5日,‘阳光玫瑰’葡萄冬芽中GA3含量在529.43~838.71 pmol·g-1范围内变化。不同时期,第1~7节位冬芽中GA3含量呈升高-下降-升高-下降的变化趋势。在6月6日,第1~7节位冬芽GA3含量最低。在6月21日,不同节位冬芽GA3含量升高,且较高节位达到最高。在7月4—19日,第1~7节位冬芽GA3含量整体呈现下降趋势。在9月2日,较低节位冬芽GA3含量最高。
在整个生理分化时期,GA3含量在6月6日最低,其中第4节位GA3含量最低,第2~7节位GA3含量显著低于第1节位。在6月21日,第3、4节位冬芽GA3含量显著低于第5、7节位。7月19日之后,第1~7节位冬芽GA3含量迅速升高;在9月2日,第1、2节位冬芽GA3含量在整个时期达到最高值,显著高于其它5个节位。在9月21日,第4节位冬芽GA3含量显著低于其他节位。
2.7 不同节位冬芽发育中ABA/GA3比值变化
如图7所示,第1~7节位冬芽ABA/GA3比值整体较为稳定,在0.51~0.92范围内变化。在6月21日,第3、4节位ABA/GA3比值首先升高,显著高于其他节位。而同时期第6、7节位比值低,其中第6节位最低。冬芽发育至7月19日,第3、4、5节位ABA/GA3的比例升高,其中第3节位比值最大。在8月18日至9月2日,ABA/GA3比值整体随节位升高而增加。
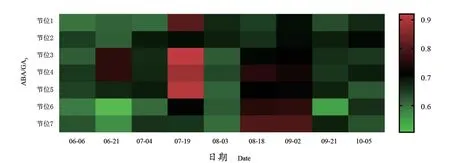
图7 不同节位各时期ABA/GA3Figure 7 ABA/GA3 at different nodes in different periods
2.8 不同节位冬芽发育中CTK/GA3比值变化
如图8所示,第1~7节位冬芽CTK/GA3比值呈现降低-升高的趋势,在0.24~0.41范围内变化。6月21日至7月19日,第1~7节位冬芽CTK/GA3比值较其他时期低;在6月21日,第2、3节位比值最高;在7月4日,第2节位最高;7月19日至8月3日,各节位CTK/GA3比值逐渐升高;在8月3日,低节位冬芽的CTK/GA3比值较高。
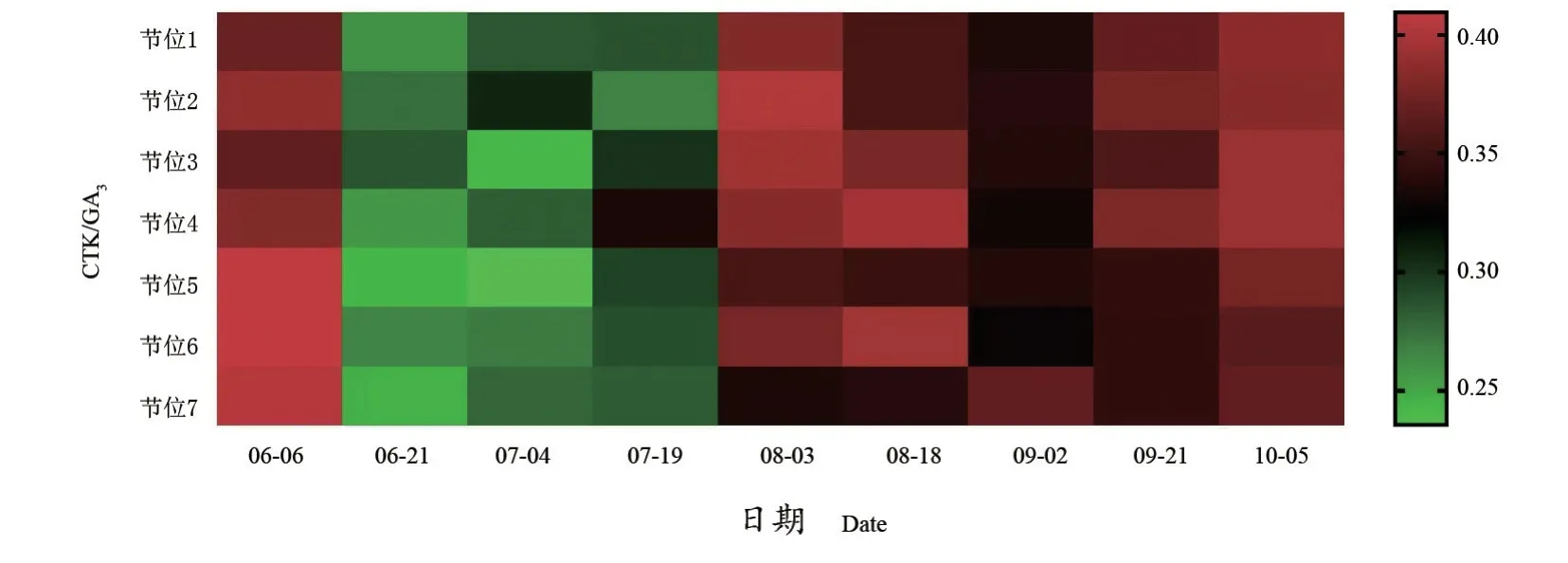
图8 不同节位各时期CTK/GA3Figure 8 CTK/GA3 at different nodes in different periods
2.9 不同节位冬芽发育中(IAA+GA3+CTK)/ABA比值变化
如图9所示,第1~7节位冬芽(IAA+GA3+CTK)/ABA比值呈现下降-上升-下降趋势,在1.59~2.74范围内变化。第3、5节位在7月19日降至最低值,其中第5节位(IAA+GA3+CTK)/ABA比值最低,第6、7节位在8月18日—9月2日降至最低。

图9 不同节位各时期(IAA+GA3+CTK)/ABAFigure 9 (IAA+GA3+CTK)/ABA at different nodes in different periods
3 讨论
植物的成花是多种激素在一定的比例下对植物进行的调控[17]。在不同物种间,相同的激素对花芽分化的调控作用并不一致。
目前,IAA与花芽分化间的关系存在争议,高水平的IAA能够促进侧柏成花[18];而在枇杷中,低水平的IAA促进成花[19]。李秉真等[20]在对苹果梨研究中发现,短枝生长停止后,较低的IAA含量有利于芽体进入生理分化,而较高的IAA含量有利于进入形态分化期;在形态分化期,较低的IAA含量有助于形态分化各时期的完成。Zhang等[21]的研究中发现,IAA信号参与花柄和花冠伸长且均有略微延迟开花的作用;鲁荣海[22]在对葡萄二次开花结果的研究中发现,高含量的IAA促进葡萄花芽分化。本研究中,在生理分化时期,中部节位IAA含量较高,说明IAA促进葡萄的花芽分化,这与王海波等[23]的研究一致。
吴志祥等[24]在研究中发现:GA3促进了营养枝的伸长从而影响荔枝的花芽分化,低水平的GA3有利于荔枝的花芽分化。谭巍[25]的研究中发现,GA可以提高蝴蝶兰的抽薹率,促进早花。通常,GA对于非诱导条件下的开花、需低温抽薹的开花及日中性不需低温的园艺植物起促进开花的作用,对于多年生果树、林木的成花起抑制作用。在本研究中,分化初期冬芽内GA3含量较低,并且中部节位GA3含量较低,验证了GA3对于葡萄花芽分化的抑制作用。
目前,ABA对花芽分化的研究还不完善。吴志祥等[24]对荔枝的研究中发现,ABA能够促进果树的成花,且上部枝条的花芽分化程度也好于下部枝条。但在鲁荣海[22]的研究中,低含量的ABA促进‘夏黑’葡萄花芽分化,说明ABA抑制成花。本研究中,在分化前期,中部节位ABA含量高,说明ABA对葡萄花芽起促进作用。有研究证明,在生理分化期,高水平的CTK促进植物花芽分化[26],这与本研究结果一致。
果树的花芽分化是由多种激素共同调控。张瑞等[27]在对紫斑牡丹研究中发现,较高ZR/IAA、ZR/GA3、ABA/GA3、ZR/IAA比值有利于花芽分化,(GA3+IAA+ZR)/ABA与(IAA+ZR)/GA3比值减小分别促进重瓣花朵花瓣原基和雄蕊原基的形成。李志娟等[28]研究发现,低水平的ABA/GA3值均有利于萱草从营养生长转向生殖生长,较高水平的ZR/GA3、(GA3+IAA+ZR)/ABA值有利于萱草花器官原基的形态建成。在本研究中,(GA3+IAA+CTK)/ABA比值小有利于葡萄成花。CTK/GA3、ABA/GA3比值越大,越有利于成花。
4 结论
在生理分化后期,第1节位冬芽花芽率为80%,第2~6节位冬芽花芽率均能达到100%;在整个生理分化时期,第1~3节位葡萄冬芽IAA、ABA、CTK含量整体较高,且第2、3节位GA3含量较低;在花芽分化前期,第2、3节位冬芽的CTK/GA3、ABA/GA3比值整体较高,且第1~3节位(IAA+GA3+CTK)/ABA比值较低,说明第2、3节位冬芽的分化质量好且花芽率高。综上,在天津双街地区‘阳光玫瑰’葡萄冬剪建议保留3个冬芽。
