饥饿胁迫对粘虫幼虫糖原、海藻糖含量和蜕皮激素滴度变化的影响
2023-06-26李文平张亚玲杨顺义王贞霖金秋彤
李文平,张亚玲,杨顺义,王贞霖,金秋彤
(甘肃农业大学 植物保护学院,甘肃省农作物病虫害生物防治工程实验室,兰州 730070)
粘虫Mythimnaseparata(Walker)为鳞翅目Lepidoptera夜蛾科Noctuidae昆虫,是著名的迁飞性、暴食性、多食性害虫。主要危害玉米、小麦、水稻等禾谷类粮食作物[1]。据史料记载,中国曾有多项粘虫暴发成灾的记录,且在多个省份均有发生,造成了严重的经济损失,甘肃等西北地区的发生为害以2代粘虫为主[2-3]。
粘虫暴发时,幼虫扩散和成虫迁飞的规模庞大,极易出现食物短缺的现象[4]。Gaurav等[5]研究了粘虫食性的能量学、不同龄期组织生长的能量学以及同化速率和呼吸速率,发现幼虫的代谢率主要取决于食物的消耗。昆虫以糖原(glycogen)形式将能量储存在脂肪体中。孟竹等[6]发现连续饥饿72 h的家蚕Bombyxmori幼虫脂肪体中糖原和甘油三酯的含量持续降低;Lutz等[7]研究发现非洲果甲虫Pachnodasinuata在饥饿状态下血淋巴中碳水化合物含量降低,脂肪体中糖原和总脂含量减少。昆虫的海藻糖(trehalose)和糖原之间可以相互转化,在逆境中起着关键作用[8-9]。孟竹等[6]发现在饥饿家蚕幼虫48~72 h之间血淋巴中海藻糖浓度升高;Hibshman等[10]发现秀丽隐杆线虫Caenorhabdataselegans的FoxO/daft 6能通过转移碳代谢促进海藻糖合成以提高线虫抗饥饿能力。蜕皮激素(ecdysone,E)能够参与调控昆虫的生长发育,对环境胁迫进行响应[11]。蜕皮激素主要由幼虫前胸腺(Prothoracic gland,PG)及成虫生殖腺合成,前胸腺主要合成前体E,之后释放到血淋巴变成有活性的20-羟基蜕皮酮(20-hydroxyecdysone,20E)[12-13]。已有研究表明,当黑腹果蝇Drosophilamelanogaster遭遇食物缺乏,饥饿信号会激活蜕皮激素的合成,使20E浓度明显升高[14];Chen等[15]发现在饥饿条件下,家蚕幼虫会通过增加蜕皮激素的生成来提高存活率。蜕皮激素在昆虫体内与海藻糖共同作用,互相影响。Tan等[16]从绿盲蝽Apolyguslucorum中克隆获得AlEcR-B,并对其表达谱进行了研究,发现在降低AlEcR-B后,海藻糖分解代谢增加。
为探究饥饿对粘虫不同龄期幼虫代谢的影响,本研究对4~6龄幼虫进行饥饿处理,分别测定正常和饥饿条件下幼虫体内糖原、海藻糖含量和蜕皮激素滴度的变化情况,旨在探讨粘虫幼虫受到饥饿胁迫时主要能源物质的变化规律,为进一步揭示饥饿肋迫下粘虫幼虫体内关键因子的调控机制以及繁殖适应性奠定基础。
1 材料与方法
1.1 材 料
1.1.1 供试昆虫 粘虫由甘肃农业大学植物保护学院农药实验室自种的新鲜玉米叶饲养。
1.1.2 供试药剂及试剂 乙醚、氢氧化钾、无水乙醇、浓盐酸、蒽酮、浓硫酸、三氯乙酸,苯基硫脲,均为国产分析纯;昆虫蜕皮激素ELISA检测试剂盒(上海优选生物科技有限公司)。
1.1.3 主要仪器 人工气候箱、电热恒温鼓风干燥箱、高压灭菌锅、高速冷冻离心机、电热恒温水浴锅、电子天平、分光光度计等。
1.2 试验方法
1.2.1 饲养条件 1~3龄幼虫用养虫缸(d=20 cm,h=30 cm)群体饲养,4龄开始用餐盒 (15 cm×10 cm×5 cm)饲养,密度为10头/盒,温度为(25±1)℃,相对湿度为(70±5)%,光周期为L14∶D10,光照度2 000~3 000 lx,在实验室人工气候箱中饲养[17]。
1.2.2 饲养方法 将卵卡放入养虫缸中,养虫缸内加适量鲜玉米叶,湿棉球保湿,缸口盖以黑布,保持空气通透,用皮筋扎紧以防初孵幼虫逃逸;幼虫孵化后,采用新鲜幼嫩玉米苗饲喂,每日更换玉米苗并清除粪便(用筛子筛出),保持养虫缸的良好环境,防止幼虫感染疾病。4龄开始转移至餐盒饲养[1]。
1.2.3 饥饿处理 幼虫4龄刚蜕皮开始,将试验分为两组,以正常饲养为对照,另一组进行饥饿处理,饥饿时间过后正常饲养[18]。
5龄和6龄处理方法同4龄。
1.3 糖原及海藻糖含量
1.3.1 不同组织样品的收集 分别选取生长健康、大小基本一致的粘虫幼虫4~6龄幼虫,用乙醚麻醉至死,在70 ℃干燥箱中烘干,碾磨成粉,待用。每处理取10头幼虫,以正常饲养为对照,重复3次[19]。
1.3.2 试剂 0.2 mg/mL葡萄糖标准溶液:取葡萄糖200 mg,用双蒸水定容至1 000 mL,待用;0.2% 蒽酮试剂:将0.2 g蒽酮溶于100 mL 95% 硫酸溶液中。
1.3.3 糖原的提取 取0.02 g样品移入1.5 mL离心管中,加入30% KOH 1 mL,沸水浴30 min,加无水乙醇0.5 mL混匀后离心10 min (4 000 r/min),弃上清液,再在沉淀物中加0.6 mol/L盐酸1 mL,沸水浴2 h后离心去沉淀,定容至1 mL,待用[19]。
1.3.4 海藻糖的提取 取0.02 g样品移入1.5 mL离心管中,加入10%三氯乙酸溶液1 mL于 5 000 r/min离心5 min,将上清液转入1.5 mL离心管中,定容至1 mL,待用[19]。
1.3.5 测定方法 糖原和海藻糖测定均采用蒽酮比色法[20],取样100 L,加蒽酮试剂2.4 mL,摇匀后沸水浴10 min,立刻冰浴2 min,冷却后于620 nm测定吸光度,标样为葡萄糖。糖原值为所得数值乘以0.927[19]。
1.4 蜕皮激素滴度测定
1.4.1 样品收集 分别选取生长健康、大小基本一致的粘虫幼虫4~6龄幼虫,每24 h取样1 次,至该龄期结束。收集血淋巴,于幼虫腹部节间膜处扎一小洞,然后轻压腹部,将流出的血淋巴滴入置于冰浴上的离心管内,每毫升血淋巴加5 L苯基硫脲,备用。每龄取刚蜕皮成功的幼虫做试虫,取样时间分别为1 d,2 d,3 d。每处理取10头幼虫,以正常饲养为对照,重复3次[21]。
1.4.2 蜕皮激素滴度测定 根据昆虫蜕皮激素ELISA检测试剂盒使用说明书测定不同时期粘虫20E滴度。
1.5 数据处理
试验数据采用Excel 2016和 SPSS 24.0软件进行统计分析,采用Duncan’s新复极差法 (P<0.05)进行差异显著性分析。
2 结果与分析
2.1 糖原含量测定结果
由图1可知:各龄期饥饿第1天时的糖原含量均低于对照组,且4龄饥饿组相比对照组显著降低,各龄期饥饿第2天和第3天时的糖原含量均显著低于对照组。从变化规律来看,对照组 4~6龄糖原含量均呈上升趋势,各龄期饲喂3 d的糖原含量分别达到43.06 mg/g、43.18 mg/g和37.30 mg/g;饥饿组随处理天数的增加,各龄期幼虫的糖原含量逐渐减少,饥饿第3天时分别为15.11 mg/g、16.13 mg/g和15.50 mg/g。除5龄1 d和6龄1 d的处理组与对照组无显著差异外,其余饥饿组的糖原含量均显著低于对照组。
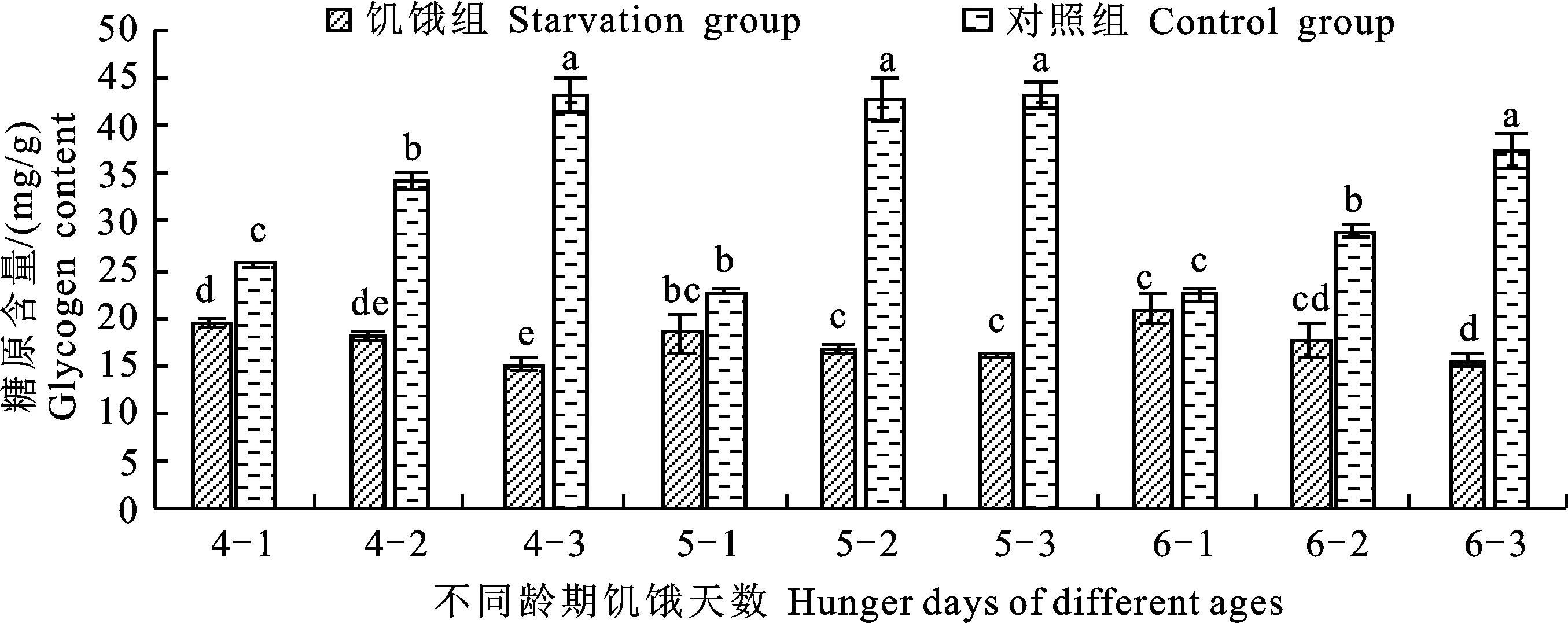
不同小写字母表示差异显著(P<0.05);图中4-1,4-2等分别表示4龄饥饿1 d,4龄饥饿2 d等。下同
2.2 海藻糖含量测定结果
如图2所示:各龄期饥饿第1天时的海藻糖含量均低于对照组,且5龄和6龄饥饿组相比对照组显著降低,各龄期饥饿第2天和第3天时的海藻糖含量均显著高于对照组。从变化规律来看,随饥饿天数的增加,各对照组的海藻糖含量变化均呈现先下降再升高的趋势,饥饿组4~6龄的海藻糖含量均逐渐升高,各龄期饥饿3 d的海藻糖含量分别达到35.41 mg/mL、39.56 mg/mL和40.06 mg/mL。
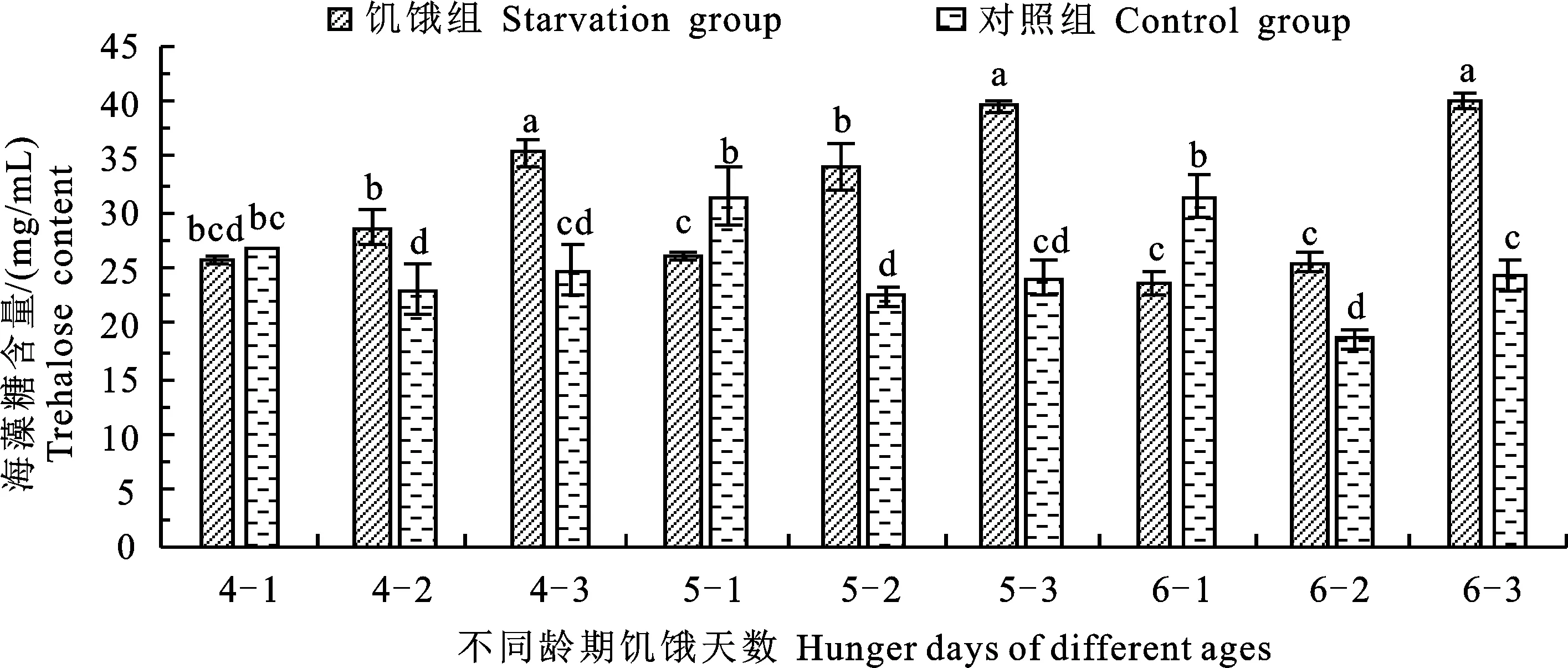
图2 不同龄期及不同饥饿天数处理下粘虫幼虫的海藻糖含量变化Fig.2 Changes of trehalose content in M.separata larvae under different days of starvation and different instars
2.3 蜕皮激素含量测定结果
由图3可知:4龄饥饿组在处理第1天和第3天时的20E滴度均高于对照组,且饥饿第3天时与对照组差异显著,饥饿第2天时的20E滴度则显著低于对照组。从变化规律来看,饥饿组在第1~3天时的20E滴度逐渐升高,对照组的20E滴度则是1~2 d时逐渐升高,在第2天时达到最大值49.71 ng/L,第3 天时显著降低,对照组有部分幼虫在4龄第3 天时即蜕皮进入5龄,而饥饿组20E滴度一直处于不断上升的状态,在第3 天时20E滴度达到39.92 ng/L,较对照第2天时的49.71 ng/L低9.79 ng/L,饥饿组在处理第3天时未见蜕皮,其生长发育明显滞后于对照组。5龄饥饿组饥饿2 d和3 d的20E滴度均显著低于对照组,从变化规律来看,5龄对照组的20E滴度随天数的增加逐渐升高,饥饿组亦呈升高趋势,但其第2 天和第3 天的20E滴度均显著低于对照组,说明5龄幼虫受饥饿胁迫后也出现生长发育减缓的现象。6龄饥饿组饥饿1~3 d的20E滴度均显著高于对照组,从变化规律来看,6龄对照组的20E滴度随天数的增加而逐渐升高,但增长幅度较小,饥饿组20E滴度则均显著高于对照组,说明6龄幼虫受到饥饿胁迫后,会通过增加体内蜕皮激素含量以便提前进入蛹期,以此来应对饥饿的环境压力。
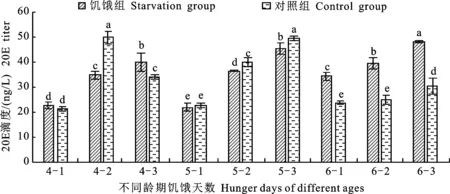
图3 不同龄期不同饥饿天数处理下粘虫幼虫的20E滴度变化Fig.3 Changes of 20E titers of M.separata larvae under treatment of different days of starvation and different instars
随处理天数的增加,4~6龄饥饿组的20E滴度均呈现逐渐升高的趋势,各龄期饥饿3 d的20E滴度分别达到39.92 ng/L、45.36 ng/L和48.03 ng/L。
3 讨论与结论
试验结果表明:4~6龄幼虫饥饿第1天时的糖原含量均低于对照组,且随处理时间的延长呈不断降低的趋势,说明幼虫受到饥饿胁迫后,糖原被消耗代谢,用于抵抗外界逆境;随饥饿时间的延长,4~6龄幼虫的海藻糖含量逐渐升高,梯度变化明显,说明在幼虫受到饥饿后,海藻糖含量会急剧增加,在第一时间参与代谢,供应机体需要以应对逆境条件。
4龄对照组的20E滴度先升高后降低,且在试验过程中发现有部分幼虫在4龄第3天即蜕皮进入5龄,而饥饿组第3 天时的20E滴度为 39.92 ng/L,较对照组的最大值49.71 ng/L低9.79 ng/L且差异显著,饥饿组在处理第3天时未见蜕皮,其生长发育明显滞后于对照组。5龄饥饿第2天和第3天的20E滴度均显著低于对照组,说明幼虫在饥饿胁迫下生长发育减缓,这与鞠珍等[22]发现饥饿后的美国白蛾Hyphantria cunea幼虫生长发育历期增长的研究结果一致。
6龄饥饿组20E滴度均显著高于对照组,有部分6龄幼虫在饥饿第3天时会进入预蛹期,说明20E可减少昆虫食物的消耗,使得末龄幼虫提前化蛹,在受到饥饿时起到一定的调控作用,这与Wang等[23]的研究结果一致,发现对家蚕末龄幼虫注射20E 24 h后,家蚕幼虫停止取食并进入游走期。
糖原、海藻糖和蜕皮激素在昆虫的生长发育、生理代谢等生长活动中都有着不可或缺的作用,当粘虫幼虫受到饥饿胁迫时,它们发挥着重要的平衡和协调作用。本试验通过对粘虫幼虫糖原、海藻糖和蜕皮激素的含量以及变化规律的观察分析,发现饥饿对粘虫幼虫正常生长代谢活动的影响较大,因此,在粘虫的综合防治中可以从饥饿方面通过抑制糖原、海藻糖的代谢及相互转化,或打乱粘虫生理代谢平衡的手段,开发昆虫生长调节剂等昆虫激素类杀虫剂。
