追毒方调控c-JUN抑制糖基化异常逆转三阴性乳腺癌耐药的机制
2023-06-10李雨璇刘春亮袁琴陈婷蒋言涛姚霏孙华郝敏侠张露蓉刘敏
李雨璇,刘春亮,袁琴,陈婷,蒋言涛,姚霏,孙华,郝敏侠,张露蓉,刘敏
(1.南京中医药大学附属苏州市中医医院,江苏 苏州 215009;2.苏州大学附属第一医院,江苏 苏州 215123)
三阴性乳腺癌(Triple-negative breast cancer,TNBC)是乳腺癌亚型中的一种,表现为雌激素受体(ER)、孕激素受体(PR)和人表皮生长受体2(HER-2)均是阴性,是乳腺癌中预后较差的一种[1]。其特征是复发风险高、无进展生存期和总生存期较短[2]。因为TNBC特殊的表现,目前临床主要以化疗手段为主,但随着表阿霉素等化疗药物大范围应用于患者,大量患者出现耐药性,导致化疗无法起到相应治疗效果[3-5]。当下寻找逆转TNBC耐药新的靶点和治疗方法是现在临床的热点与难点。
近年来,中医针对耐药性乳腺癌的治疗取得不错的成绩[6-8]。历代的诸家医学书籍将所有类型的乳腺癌统一归属为“乳岩”范畴,为聚痰成积、凝结于乳中而成癌。对于TNBC耐药的逆转,课题组长期的临床实践发现,除加强解毒破瘀清除癌毒,还需要祛湿利水消肿。因此,课题组选择雷公藤为君,干蟾皮为臣,红豆杉为佐,以1∶1∶1比例为核心方药,辨证加减应用于临床,表现出增强解毒破瘀抗癌,又兼有祛湿利水消肿的作用[9]。结合张伯礼院士提出有效组分组方研究现代中药模式,具有疗效好、用量少、质量可控等优势的观点[10-11],课题组取三药有效成分提取物雷公藤甲素、华蟾素毒基和紫杉醇(质量比1∶1∶1)组成“追毒方”[12],展开相应的实验研究。结果显示,追毒方可抑制β1,3-N-乙酰氨基半乳糖基转移酶-8(β3GnT8)逆转MDA-MB-231/TAX的耐药性[13];成分雷公藤甲素能抑制N-乙酰氨基半乳糖转移酶1/2(ppGalNAc T1/2)和β3GnT8,影响糖蛋白CD147的糖基化修饰,使得TNBC细胞的侵袭性与迁移能力得以削弱[14]。前期的研究表明追毒方可逆转TNBC的耐药性,阐明了部分机制,但深层机制并不明朗。
课题组前期研究显示c-JUN可直接作用于直肠癌细胞的β3GnT8启动区,导致β3GnT8转录激活[15],且追毒方可抑制c-JUN和β3GnT8的异常[13]。因此,本研究旨在明确以下几点:①明确追毒方下调TNBC的耐药糖蛋白与下调β3GnT8和ppGalNAc-T1/2表达相关;②明确追毒方逆转TNBC耐药是通过关键调控因子c-JUN,导致β3GnT8和ppGalNAc-T1/2表达下调,进而下调CD147的表达所致。为后续追毒方在TNBC的实验研究以及临床治疗方面提供更多的实验依据。
1 材料
追毒方中的华蟾素毒基、雷公藤甲素和紫杉醇由苏州大学附属第一医院提供,购于南京乐泊美医药科技有限公司,MDA-MB-231耐药细胞由上海莼试生物技术有限公司,0.25%Tripsin-EDTA(南京凯基生物科技发展有限公司,KGY001),胰酶(BIOSHARP公司),PBS(南京凯基生物科技发展有限公司,KGB500),胎牛血清(浙江天航生物科技股份有限公司),L-15培养基购于柯意哲(上海)机电工程有限公司,BCA蛋白浓度测定试剂盒(碧云天生物技术有限公司,P0010),转染试剂(Invitrogen公司,货号:11668-019),GALNT1 Polyclonal Antibody、GALNT2 Polyclonal Antibody、β3GNT8 Polyclonal Antibody均购于武汉三鹰公司,BCRP、ABCG2 Polyclonal Antibody购于proteintech公司,HRP山羊抗兔IgG(H+L)(碧云天生物技术有限公司,A0208),MDR1/ABCB1(E1Y7B) Rabbit(CST公司,13342S)。
2 方法
2.1 追毒方逆转TNBC耐药的细胞实验
2.1.1 追毒方逆转TNBC耐药的细胞培养 人乳腺癌细胞阿霉素耐药株MDA-MB-231/ADR常规培养于含100 mL·L-1小牛血清且含有250 ng·mL-1阿霉素的L-15培养基中,于实验前脱阿霉素1周后使用。37 ℃的5%CO2培养箱中培养,每2 d换液1次,并在显微镜下观察细胞贴壁生长情况,当细胞生长至约75%~80%时进行传代,传代培养后取活力较强的耐药细胞株进行实验。
2.1.2 MTT法检测细胞的活性 将耐药MDA-MB-231/ADR细胞消化、计数、配制成浓度为:3×104~5×104mL-1的细胞悬液,96孔板中每孔加入100 μL细胞悬液,第2天待细胞贴壁后,小心将上清倒掉,分成5组:空白组、阳性对照组(多西他赛1.7 μg·mL-1)、追毒方按照3个单成分的质量比1∶1∶1配置并分为3个浓度(0.375、0.75、1.5 μg·mL-1),用完全培养基配制好的各组药物溶液每孔加入100 μL,在37 ℃、5%CO2及饱和湿度条件下分别培养24、48、72 h。培养结束后,加MTT溶液(5 mg·mL-1) 20 μL·孔-1,37 ℃、5%CO2孵箱中继续孵育4 h。离心后弃去培养液,用PBS冲2~3遍,每孔加入150 μL二甲基亚枫充分溶解。在酶联免疫检测仪490 nm处测量各孔的吸光值。按下式计算细胞生长抑制率:生长抑制率(%)=(1-药物组OD值/对照组OD值)×100%,绘制生长曲线。
2.1.3 瞬时转染c-JUN过表达质粒建立高表达c-JUN的MDA-MB-231/ADR细胞 转染质粒c-JUN的构建由Invitrogen公司完成,将MDA-MB-231/ADR细胞分为6组:MDA-MB-231/ADR空白对照组、空白质粒对照组、转染c-JUN质粒对照组,MDA-MB-231/ADR加药组、空白质粒加药组、转染c-JUN质粒加药组。转染前1 d,接种适当数量的MDA-MB-231/ADR细胞至细胞培养板中,每孔中加入不含抗生素的培养液,使转染时的细胞密度能够达到30%~50%。设置A管:将5 μg DNA(c-JUN质粒和空白质粒)分别溶于250 μL Opti-men无血清培养基中,设置B管:将125 μL lip3000溶于125 μL Opti-men无血清培养基中,混匀室温放置5 min,将A、B两管混合成C管,再放置20 min。转染期间,将6孔板培养基换成无血清培养基,每孔400 μL。将C管配好的DNA-脂质体复合物分别加入6孔板对应孔中,4~6 h后换成有血清培养基,加药组加0.75 μg·mL-1的追毒方。置于37 ℃的CO2培养箱中培养48 h。用MTT法检测细胞活性。
2.2 追毒方抑制裸鼠耐药TNBC原位模型的实验
将耐药MDA-MB-231/ADR细胞消化、计数、配制成浓度为:3×104~5×104个细胞·mL-1的细胞悬液。取5~6周龄雌性裸鼠,随机分成5组:[空白组、阳性对照组(多西他赛10 mg·kg-1)、追毒方分为3个浓度(0.1、0.5、2.5 mg·kg-1)],细胞接种:于裸鼠的乳腺脂肪垫内0.1 mL细胞悬液(1×106个细胞),观察瘤子生长情况,1周可明显见瘤块突起于皮肤。开始给药,雷公藤甲素、华蟾素毒基及紫杉醇按质量比1∶1∶1配置成浓度为0.01、0.05、0.25 mg·mL-1的追毒方,每天按照10 mL·kg-1灌胃量给药1次;阳性对照组多西他赛采用腹腔注射,每周2次;模型组分别对应给予溶媒。2周后处死小鼠,取出瘤子并拍照。取出肿瘤称质量,计算抑瘤率,抑瘤率=(1-实验组瘤质量/对照组平均瘤质量)×100%。留取部分用于蛋白表达分析。
2.3 Western blot法检测c-JUN、β3GnT8和ppGalNAc-T1/2、CD147及相关耐药蛋白的表达
将各组细胞用胰酶消化收取于EP管;将裸鼠组织蛋白匀浆后备用,加入适量的RAPI buffer(50 mmol·L-1Tris pH8.0、150 mmol·L-1NaCl、1%Triton X-100、0.5%Sodium deoxycholate、0.1%SDS、2 mmmol·L-1EDTA、5%Glycerol、1× protease inhibitors cocktail、1 μmol·L-1DTT和PMSF),冰上30 min,裂解提取全细胞总蛋白/裸鼠组织蛋白,12 000 r·min-1离心15 min,收取上清,用Bradfold法检测总蛋白浓度,取50 μg蛋白的细胞裂解液与蛋白上样缓冲液4∶1混合,煮沸使蛋白变性,配制SDS-聚丙烯酰胺凝胶,放入电泳槽中,加满SDS电泳缓冲液,在上样孔中加入蛋白样品,电泳至跑出凝胶,转膜,封闭,浸在一抗中过夜,漂洗3次后浸于二抗中室温孵育1~2 h,再漂洗3次。将洗好的膜放置于荧光可见光成像系统主机内,加显色液,曝光,获取目标蛋白条带。
2.4 数据分析
3 结果
3.1 追毒方对MDA-MB-231/ADR耐药的逆转效应
不同浓度追毒方作用于耐药MDA-MB-231/ADR细胞,可显著抑制MDA-MB-231/ADR细胞的增殖(P<0.01),具有时效和量效关系(图1A);也可降低抑制MDA-MB-231/ADR细胞中耐药蛋白BCRP和MDR1表达(P<0.01)。(图1B)

注:Model.模型组;DX.多西他赛组;与模型组比较,**P<0.01。图1 追毒方对MDA-MB-231/ADR耐药的逆转效应Fig.1 Reversal effects of Zhuidu Formula on MDA-MB-231/ADR drug resistance
对于耐药MDA-MB-231/ADR腋下移植瘤裸鼠模型,追毒方可降低瘤体质量,而对荷瘤裸鼠的体质量无影响(图1C~D);当给药剂量为2.5 mg·kg-1时可显著抑制瘤体生长(P<0.01)。且对比模型组,追毒方可明显降低肿瘤组织中耐药蛋白BCRP和MDR1表达(P<0.01,图1E~G)。提示追毒方对MDA-MB-231/ADR的耐药性具有逆转的效应,且在体内体外皆有逆转效应。
3.2 追毒方对耐药调控通路c-JUN,β3GnT8和ppGalNAc T1/2、CD147蛋白的影响
耐药MDA-MB-231/ADR腋下移植瘤裸鼠模型的肿瘤组织进一步研究结果显示,追毒方在降低耐药蛋白表达的同时,可明显降低c-JUN、β3GnT8、ppGalNAc-T1/2和CD147蛋白的表达(P<0.01)(图2)。提示追毒方可能通过下调c-JUN表达,进而抑制β3GnT8-ppGalNAc T1/2-CD147通路,逆转MDA-MB-231/ADR的耐药。
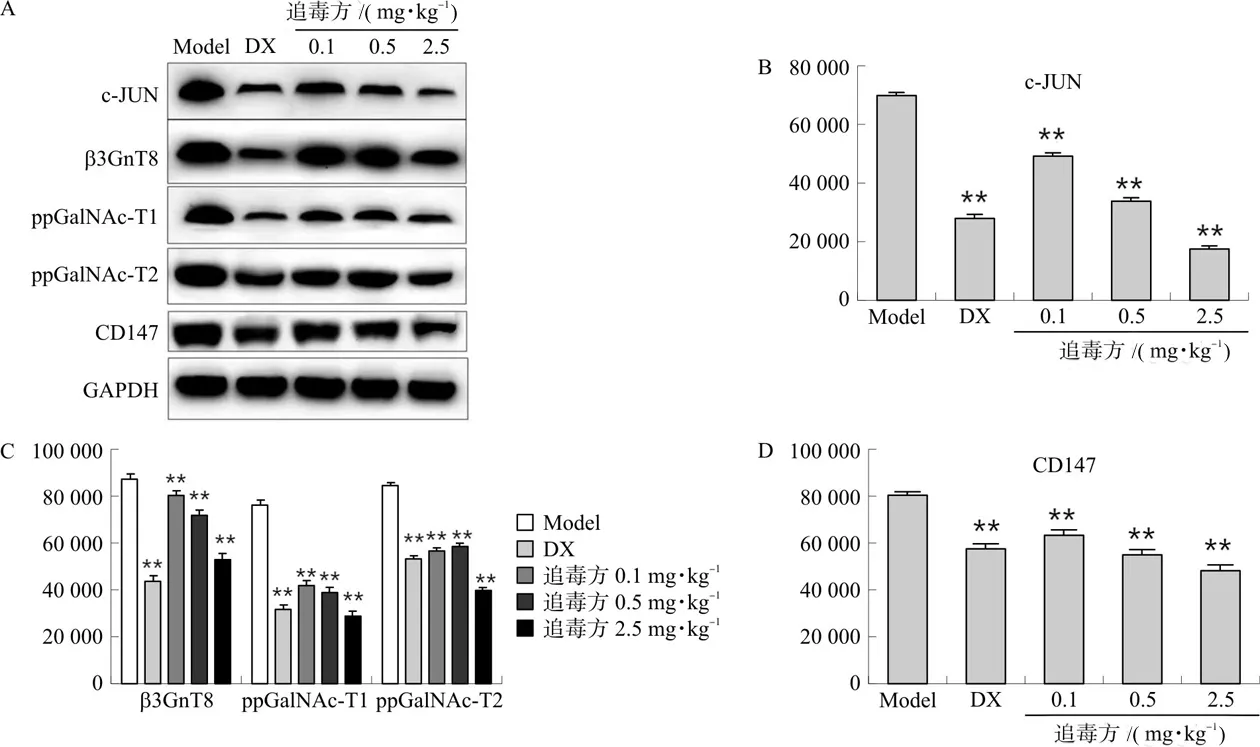
注:Model.模型组;DX.多西他赛组。与模型组比较,**P<0.01。图2 追毒方对c-JUN,β3GnT8和ppGalNAc T1/2、CD147蛋白的影响Fig.2 Effects of Zhuidu Formula on c-JUN, β3GnT8, ppGalNAc T1/2 and CD147 proteins
3.3 追毒方干预c-JUN调控的β3GnT8和ppGalNAc T1/2-CD147表达逆转MDA-MB-231/ADR耐药的机制
应用转染技术,观察过表达c-JUN对追毒方逆转MDA-MB-231/ADR耐药的影响,进一步明确其机制。结果显示:与空白对照组比较,空白质粒对照组和转染c-JUN质粒对照组对细胞增殖均无显著影响(P>0.05)。给予追毒方后,空白加药组、空白质粒加药组、转染c-JUN质粒加药组的细胞增殖,均有明显的抑制作用(P<0.05,P<0.01);抑制率空白加药组、空白质粒加药组、转染c-JUN质粒加药组依次降低(图3A~B)。提示过表达c-JUN可影响追毒方对MDA-MB-231/ADR的抑制效应。

注:1~6.空白对照组、空白质粒对照组、转染c-JUN质粒对照组、空白加药组、空白质粒加药组、转染c-JUN质粒加药组。与空白对照组比较,*P<0.05,**P<0.01;与空白质粒对照组比较,#P<0.05,##P<0.01;与转染c-JUN质粒对照组比较,^P<0.05,^^P<0.01。图3 追毒方干预c-JUN调控的β3GnT8和ppGalNAc T1/2-CD147表达逆转MDA-MB-231/ADR耐药的机制Fig.3 Mechanism of Zhuidu Formula intervening c-JUN-regulated β3GnT8, ppGalNAc T1/2, and CD147 expression and reversing MDA-MB-231/ADR drug resistance
进一步的Western blot检测结果:①空白质粒对照组与对空白对照组相比显示,空白质粒转染对c-JUN、β3GnT8、ppGalNAc T1/2、CD147、BCRP和MDR1的蛋白表达没有影响(P>0.05)。②与空白质粒对照组比较,转染c-JUN质粒对照组中β3GnT8、ppGalNAc T1/2、CD147、BCRP和MDR1的蛋白表达均显示增加(P<0.05,P<0.01)。③给予追毒方后,可显著抑制正常表达和过表达c-JUN的MDA-MB-231/ADR细胞中c-JUN、β3GnT8、ppGalNAc T1/2、CD147、BCRP和MDR1的蛋白表达(P<0.05,P<0.01)(图3C~I);但转染c-JUN质粒加药组表达水平显著高于空白质粒加药组(P<0.05,P<0.01)。综上所述,追毒方至少部分通过干预c-JUN调控的β3GnT8和ppGalNAc T1/2激活,从而抑制CD147表达,进而逆转MDA-MB-231/ADR耐药。
4 讨论
TNBC是乳腺癌中最具侵袭性的分子亚型[5],目前治疗仍以化疗为主,但因为大部分患者存在化疗耐药问题,寻找逆转TNBC耐药新的靶点和治疗方法已是刻不容缓。中医治疗耐药性TNBC有着自己独特的[16-17]优势。基于我们长期的临床实践和前期的实验研究,追毒方能逆转TNBC耐药,但机制阐述不够清晰。检索文献,相关研究显示雷公藤的有效成分雷公藤甲素,对于耐药的肺癌[18]、前列腺癌[19]、人骨肉瘤细胞[20]等均可实现逆转。干蟾皮中的有效成分华蟾素毒基,可以抑制乳腺癌细胞的侵袭和转移功能[21],也是一种有效的肿瘤多药耐药逆转剂[22]。红豆杉的有效成分紫杉醇,能有效抑制耐药乳腺癌细胞的增殖、迁移和侵袭,并可逆转其对乳腺癌耐药细胞的耐药和上皮细胞间充质转化(EMT)[23]。据此我们把三药有效成分雷公藤甲素、华蟾素毒基和紫杉醇(质量比1∶1∶1)组成追毒方。本文的研究结果再次显示无论体内体外,追毒方都可以逆转TNBC的阿霉素耐药,且毒害副作用较小。
蛋白质糖基化的变化是肿瘤细胞区别于正常细胞的特征之一[24],主要分为N-连接糖基化(N-糖基化)和O-连接糖基化(O-糖基化)[25-26]。众多研究表明,N-糖基异常化是肿瘤细胞耐药的分子生物学基础[27-28]。N-糖基化是通过β1,3-N-乙酰氨基葡萄糖基转移酶(β3GnTs)加入聚-N-乙酰乳糖胺(聚乳糖胺)进一步细长支链的N-聚糖[29]。β3GnT8是β3GnTs家族成员之一,β3GnT8在四丁烯N-聚糖上发挥活性并拉长聚乳糖胺链,其在多种肿瘤均有高表达[29-30]。Zhang等研究表明,β3GnT8在白血病阿霉素耐药细胞中高表达,下调β3GnT8的表达可增加其对化疗药物敏感性[27]。袁雅琴等[31]证实雷公藤甲素能显著抑制乳腺癌耐药细胞的增殖,是通过抑制β3GnT8等糖基转移酶基因的转录和表达,阻止多聚乳糖胺的合成所致。ppGalNAc-Ts是一类酶家族,可以启动黏蛋白O-糖基化,形成GalNAc-Ser/Thr的结构,称为TN抗原,TN抗原在肿瘤细胞中经常过度表达[32]。徐徐等[33]已经证实ppGalNAc-T1/2在TNBC中通过降低O-糖链表达,有利于药物与靶点接触,提高药物敏感性,进而逆转耐药;但其逆转耐药的准确机制仍不清晰。CD147膜蛋白通过基质金属蛋白酶(Matrix metalloproteinases,MMPs)的分泌,它在恶性肿瘤的病程进展中占有重要的地位[34]。β3GnT8可通过转化CD147的N-糖基化形式,改变CD147携带的多乳糖胺结构,进而影响乳腺癌的侵袭能力。但CD147与ppGalNAc-T1/2的关系只能推测为从ppGalNAc-T1作用于MMP14,而CD147又可以通过MMPs的分泌。因此,我们研究β3GnT8、ppGalNAc-T1/2和CD147在TNBC耐药中的表达变化。结果显示,在耐药MDA-MB-231/ADR细胞和相关肿瘤组织中,均显示高表达;追毒方在降低耐药蛋白表达的同时,可明显降低β3GnT8、ppGalNAc-T1/2和CD147蛋白的表达,表明追毒方可能通过抑制β3GnT8和ppGalNAc-T1/2,从而影响CD147,达到逆转TNBC耐药。
我们前期研究证实c-JUN可直接作用于结直肠癌细胞的β3GnT8启动子区域,导致β3GnT8的转录激活,从而促进肿瘤浸润和转移因子的表达[15]。本文研究也显示,通过转染技术过表达c-JUN后,可促使β3GnT8、ppGalNAc T1/2、CD147、BCRP和MDR1的蛋白表达均显示增加,表明c-JUN是引发TNBC耐药发生的启动蛋白。给予追毒方后,可降低c-JUN的表达,进而抑制β3GnT8、ppGalNAc T1/2、CD147、BCRP和MDR1的蛋白表达。因此,我们认为追毒方通过调控因子c-JUN,作用于糖基转移酶β3GnT8和ppGalNAc-T1/2,进而抑制N-糖基化和O-糖基化,改变相关聚糖胺结构,导致耐药蛋白表达下调、阻断耐药途径,进而逆转TNBC耐药。
本文在确定动物实验的给药方式时,考虑到临床使用的是三味中药饮片,给药方式是口服,为了使实验研究更贴近临床,因此采取灌胃给药方式。另外,我们的研究结果显示追毒方与阳性对照组多西他赛,对阿霉素引发TNBC耐药的逆转效应相近,但研究报道显示多西他赛在应用中也会产生多西他赛耐药,不利于临床治疗的进展[35]。而追毒方作为来源于中药成分的组方,我们的研究显示可扭转多种化疗药物引发的TNBC耐药性(如紫杉醇耐药[13],阿霉素耐药等),这也是中药追毒方的优势之一。在追毒方作用于空白质粒组后,CD147蛋白表达有所下降,但没有统计学意义,具体原因还有待进一步分析。
本文采取细胞、动物实验结合的方法,在整体水平上进一步探讨追毒方逆转TNBC耐药效用,并进而探讨其发生效用的分子机制,与糖基化异常相关,可能的作用靶点为糖基转移酶β3GnT8和ppGalNAc-T1/2和相关糖链结构,其靶糖蛋白包括c-JUN、CD147及相关耐药糖蛋白;涉及的信号通路:c-JUN-GT-CD147-耐药蛋白、β3GnT8-ppGalNAc-T1/2-CD147通路等途径。
