不同裂解温度生物炭对镉胁迫下烤烟生长特性和镉积累的影响
2023-06-04杨晓锋姜永雷蔺璟煜杨自云黄晓霞程小毛
杨晓锋 姜永雷 蔺璟煜 杨自云 黄晓霞 程小毛


摘要:为探究镉(Cd)胁迫下添加不同裂解温度生物炭对烤烟生长、生理及镉积累的影响,以烤烟为试验材料,通过向镉污染的土壤中添加不同裂解温度的生物炭,研究不同裂解温度生物炭对镉污染烤烟的缓解效果。结果表明,镉胁迫抑制了烤烟的生长,而添加不同裂解温度的生物炭不同程度缓解了镉对烤烟的毒害作用,添加生物炭显著增加了烤烟叶片的干、鲜质量和最大叶面积,其中裂解温度为700 ℃生物炭处理增加最为显著,分别增加了15.6%、12.2%和67.3%。与CK相比,添加生物炭降低了烤烟超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)活性,增强了烤烟的抗性。添加不同裂解温度生物炭降低了烤烟中镉元素的含量,降低了烤烟各组织的镉富集系数,一定程度上缓解了镉元素对于烤烟的胁迫,其中裂解温度为500 ℃的生物炭效果最好。综上所述,添加不同裂解温度的生物炭可以缓解镉元素对烤烟胁迫,一定程度上改善烤烟的生长环境,提高烤烟的产量和品质,其中,添加裂解温度为500 ℃的生物炭对于缓解烤烟的镉胁迫效果较好。
关键词:烤烟;镉胁迫;生物炭;抗氧化酶活性;富集系数
中图分类号:S572.01 文献标志码:A 文章编号:1002-1302(2023)09-0227-07
基金项目:云南省科学技术厅基础研究专项(编号:202001AU070006);云南中烟工业有限责任公司重点项目(编号:2021YL03);中国烟草总公司云南省公司科技计划(编号:2019530000241011)。
作者簡介:杨晓锋(1997—),男,山东烟台人,硕士研究生,从事园艺与种业技术研究。E-mail:151890640qq.com。
通信作者:程小毛,博士,教授,从事植物生理与分子研究。E-mail:xmcheng0103@gmail.com。
重金属镉(Cd)毒性极强,能够在土壤中长期存在,且极易转移,是植物生长发育的非必需元素[1]。镉对多数植物均具有生理毒性,在镉胁迫下,植物生长发育会受到伤害,包括抑制种子萌发、减缓植物生长速率、脂质过氧化、气孔开放受抑制、破坏光合机构及阻碍矿质养分吸收、转运紊乱等,最终导致作物产量和品质降低[2-4]。
烟草(Nicotiana tabacum L.)属于茄科烟草属一年生或有限多年生草本植物,是世界范围内重要的经济作物[5]。2020年我国烟草行业实现工商税利总额12 803亿元,同比增长6.2%,财政总额12 037亿元,增长2.3%[6]。云南省是我国规模最大的烤烟生产地,云南40%土地的自然条件都很适宜烤烟生长,使得云南的烤烟产量占据全国烤烟总产量的 1/3[7]。烤烟是镉的富集植物,且极易在烤烟叶片中积累[8]。镉作为一种烤烟的非必需元素,积累过量会导致烤烟植株生长受到影响,进而使烤烟的品质和产量受到影响。研究发现,镉会通过吸入的烟气进入人体内,随着镉元素进入人体后会出现高血压、肺癌、精神分裂等一系列病症[9]。因此,如何缓解重金属镉对烤烟生长发育的毒害作用,减少镉在烤烟中的积累,是烤烟产业亟待解决的难点,也是未来烤烟研究的重要方向。
生物炭是一种土壤改良碳,由各种农业废弃材料如秸秆、畜禽粪便等,在高温缺氧或者限氧的条件和相对较低的温度(≤700 ℃)下热裂解产生的一种化学性质稳定且含碳量丰富的木炭[10]。生物炭不仅比表面积、孔隙度较大,化学基团丰富,同时还拥有很强的吸附能力,能够吸收土壤里的水分子、无机离子和金属离子[11-12]。土壤中加入生物炭后能显著提高土壤的含碳量和一些有效成分的活性,增强植物对营养物质的吸收使得植物能更好地生长,以至于生物炭能作为土壤碳截留并促进植物生长的一种优质材料[13]。生物炭还能改善土壤中一些微生物的生存环境,改变土壤成分的生物有效性,让植物能更好地从土壤中吸收营养成分更利于生长[14]。有研究表明,适量施用生物炭可以提高作物净光合速率、气孔导度、光合能力[15]。
多项研究表明,添加生物炭能够缓解镉对植物的胁迫。陈越等研究发现,添加生物炭能够显著降低土壤中有效态镉的含量,减少镉在菠菜中的积累[16]。刘桂华等研究发现,添加玉米生物炭能显著增加白芨块茎和整株生物量,对镉污染土壤有显著的修复效果[17]。本研究以烤烟为试验材料,采用土培法探讨不同裂解温度生物炭对镉胁迫下烤烟生长参数、光合参数、抗氧化酶活性以及烤烟镉含量的影响,以期为烤烟生产中镉污染治理以及生物炭的选择提供一定的参考和理论依据。
1 材料与方法
1.1 试验材料
选取长势相对一致、生长情况较好的半年生盆栽烤烟幼苗为试验材料,将烤烟幼苗分别栽种于塑料盆中。土壤基质由腐殖土和红土以质量比为 1 ∶ 2 的比例混合而成,每盆土质量为 5.0 kg。将烤烟幼苗都放置在温度为22~25 ℃、相对湿度为50%~70%的西南林业大学后山树木园实验基地大棚内,试验时间为2021年5—7月。在整个试验阶段期间,都保持烤烟幼苗有充足的水分,也避免烤烟幼苗有病虫害的情况。
1.2 试验设计
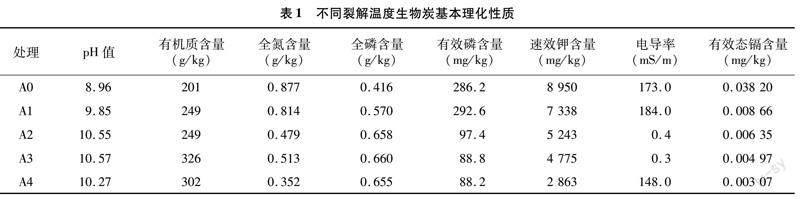
本次试验设置了镉胁迫处理组,根据加入生物炭不同分为6个不同处理组,分别是为CK、A0(牛粪原样)、A1(裂解温度350 ℃牛粪生物炭)、A2(裂解温度500 ℃牛粪生物炭)、A3(裂解温度600 ℃牛粪生物炭)、A4(裂解温度700 ℃牛粪生物炭)。不同裂解温度生物炭基本理化性质见表1。每盆种植1株烤烟,每个处理组重复3次,生物炭含量控制为5%。重金属镉元素由氯化镉(CdCl2)提供。土壤中的镉元素含量为10 mg/kg。
1.3 测定项目与方法
1.3.1 生物炭的表征测定 采用电镜扫描仪进行观察分析,放大倍数为500倍。
1.3.2 光合色素、气体交换参数测定 光合色素采用丙酮-乙醇(等体积比)混合液浸提法[18]进行测定。选取无病虫害、长势一致的第3至第5成熟功能叶片,用LI-6400便携式光合仪测定叶片的气体交换参数。
1.3.3 其他生理指标 超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)法[19]测定;过氧化物酶 (POD)活性采用愈创木酚法[20]测定;过氧化氢酶(CAT)活性采用紫外吸收法[21]测定;抗坏血酸过氧化物酶(APX)活性参照Knorzer等的方法[22]测定。
1.3.4 生长指标测定 在收样时记录每个处理组的株高、最大叶面积、叶片数。收取所有的烤烟植株,用去离子清洗干净,用直尺测量其根长。将根、叶分开分别测量其鲜质量,之后放入烘箱中105 ℃杀青30 min后,转65 ℃烘48 h至恒质量,测量其干质量。
1.3.5 镉含量测定 镉含量采用原子吸收分光光度计(Hitachi 180-80)测定[23]。将烤烟根、茎、叶样品,研磨成粉末,称取1.000 g,放入烧杯中,加入9 mL浓硝酸和3 mL高氯酸后,摇匀后过夜。第2天进行加热消煮,待样品消煮至白色结晶,冷却后加入5%的硝酸定容至25mL待测定。土壤样品中镉全量采用同样的方法测定。
1.3.6 烤烟镉富集系数、转移系数 富集系数=植物各部分镉含量/土壤中镉含量;转移系数=地上部分镉含量/根中镉含量[24]
1.4 数据处理与分析
采用SPSS 19.0軟件进行一元方差分析(one-way ANOVA)及相关性分析,平均数间的多重比较采用Duncans检验方法,α=0.05;分别采用Microsoft Excel 2010和Origin 2019绘制三线表和柱状图。
2 结果与分析
2.1 不同裂解温度生物炭的表面特征
扫描电镜(SEM)是一种常用的物质结构和形态特征的表征手段。本试验分别选取了牛粪原样以及裂解温度(350、500、600、700 ℃)下生物炭的SEM图,放大倍数为500倍。由图1可知,不同裂解温度的生物炭其表面特征有很大差别。从图1-a可以看出,牛粪原样主要呈显著大颗粒堆积形态,随着裂解温度的升高,350 ℃时大颗粒开始有一部分被分解,零星出现了一些微孔结构。当裂解温度升高到500 ℃时,一部分大颗粒被分解,出现了一部分中孔结构。当裂解温度升高到600 ℃时,形成排列较为规则的孔隙结构。当裂解温度为700 ℃时,孔隙排列更为规则且紧密,孔隙结构变大。
2.2 不同裂解温度生物炭对镉胁迫下烤烟生长参数的影响
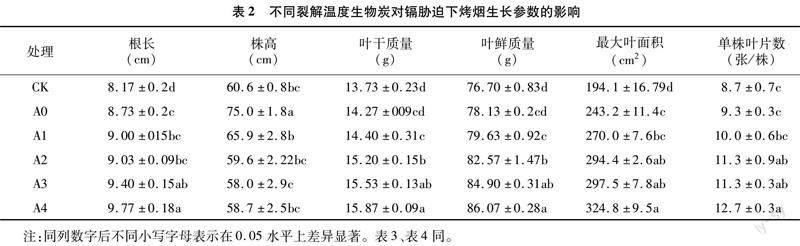
由表2可知,与CK相比,添加生物炭能显著增加烤烟的根长,A4处理增加最为显著,增加了19.6%。对于株高而言,与CK相比,A0处理的株高显著增加,A1、A2、A3、A4处理较CK变化不显著。对于叶片干、鲜质量而言,与CK相比,A0处理组变化不显著,而添加生物炭均显著增加了烤烟的干、鲜质量,其中A4处理组变化最显著,干、鲜质量分别增加了15.6%和12.2%。对于最大叶面积而言,与CK相比,添加生物炭均显著增加了叶面积,表现为A4>A3>A2>A1>A0。对于单株叶片数而言,与CK相比,A2、A3、A4处理单株叶片数显著增加,A0、A1处理较CK变化不显著。以上说明添加生物炭有利于烤烟的生长。
2.3 不同裂解温度生物炭对镉胁迫下烤烟气体交换参数的影响
由表3可知,相较于CK,添加不同裂解温度生物炭的烤烟叶片净光合速率(Pn)A1、A3处理显著增加,分别增加了11.4%、14.2%,A0、A2、A4处理组变化不显著。而胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr)均无显著变化。
2.4 不同裂解温度生物炭对镉胁迫下烤烟光合色素含量的影响
由表4可知,与CK相比,A0、A2、A3、A4处理的烤烟叶片类胡萝卜素含量分别减少了13.4%、28.1%、23.7%、33.0%,A1处理类胡萝卜含量增加了0.6%。除A1处理外,叶绿素a含量和叶绿素b含量与对照组相比均显著降低(P<0.05)。各处理组总叶绿素含量与对照组相比表现出下降趋势,分别下降了15.6%、1.8%、30.0%、26.6%、37.0%。以上结果说明在镉胁迫下添加不同裂解温度生物炭的烤烟叶绿素含量均低于对照组。
2.5 不同裂解温度生物炭对烤烟抗氧化酶活性的影响
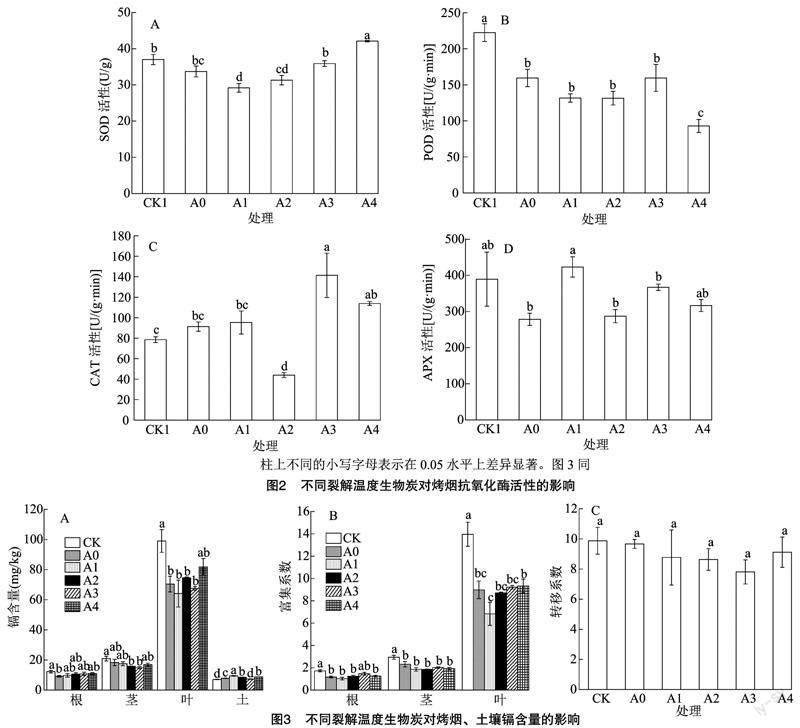
由图2可以发现,与CK相比,添加不同裂解温度生物炭的烤烟超氧化物歧化酶(SOD)活性有显著变化,除A4呈显著升高外,其他处理均呈下降趋势,其中A1处理下降最为显著,下降了21.1%(图2-A);过氧化物酶(POD)活性与CK相比都显著下降,分别降低了28.3%、、40.8%、40.9%、28.3%和58.2%,且A0、A1、A2、A3处理之间差异不显著(图2-B);A2处理组过氧化氢酶(CAT)活性与CK相比显著降低,降低了44%,其他处理组与CK相比均有所上升,其中A3和A4与CK差异显著(P<0.05)(图2-C);抗坏血酸过氧化物酶(APX)除了A1活性表现升高外,其他处理组酶活性与CK相比均呈现下降趋势,A0、A2、A3、A4组的APX含量分别下降了28.5%、26.3%、5.7%、18.7%(图2-D)。
2.6 不同裂解温度生物炭对土壤和烤烟各部位镉含量的影响
由图3-A可知,烤烟各部位镉含量表现为叶>茎>根。各处理组烤烟根部的镉含量与CK相比均降低,但各处理组之间无显著差异;各处理组茎镉含量与CK相比均表现出降低趋势,其中A2和A3与CK之间呈现显著差异,A0、A1和A4处理与CK无显著差异;各处理组烤烟叶片的镉含量与CK相比均表现出降低趋势,其中A0、A1、A2和A3各处理组之间无显著差异,但与CK之间表现出显著差异。各处理组植烟土壤镉含量与CK相比均有一定程度增加。由图3-B可知,与CK相比,添加生
物炭顯著降低了烤烟的富集系数,其中A1组的富集系数相比较而言最低,根、茎、叶的富集系数分别降低了40.1%、58.1%、51.1%。由图3-C可知,转移系数CK与各处理组之间差异不显著。
2.7 镉含量与叶片抗氧化酶活性和叶绿素含量的相关性分析
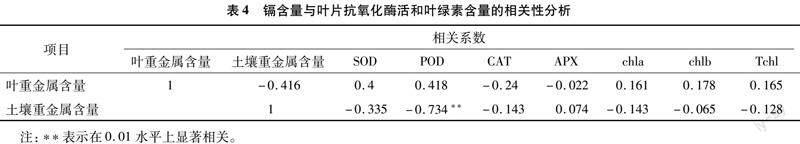
由表4可知,叶片重金属含量与抗氧化酶活性和叶绿素含量之间的相关性不显著,而土壤重金属含量与POD活性呈极显著负相关关系(P<0.01),与其他抗氧化酶活性和叶绿素的相关性不显著。
3 讨论与结论
生长参数是反映植物生长发育状况和抗逆性的重要指标,在镉胁迫下植物生长参数会大幅降低[25]。根系是植物生长过程中供应水分和营养物质的主要器官,能调节植物地上部分的生长,根系的形态特征对整个植物的生长尤为重要[26]。本研究结果表明,与对照组相比添加不同裂解温度的生物炭均能促进烤烟根长的生长,表现为A4>A3>A2>A1处理。说明添加生物炭缓解了镉胁迫对于烤烟根系生长的抑制,而烤烟叶的干质量、鲜质量、最大叶面积和叶片数也都显著增加,进一步说明在镉胁迫下添加生物炭对烤烟植株生长具有促进作用。其原因主要为添加生物炭使得土壤中的镉被生物炭吸附,土壤中的有效态镉含量下降,烤烟受到的胁迫减少,生长发育得到缓解[27]。此外,生物炭还可以通过改善土壤理化性质、减少土壤养分流失、促进植物养分的增加来促进烤烟植株的生长发育,从而进一步缓解镉胁迫对烤烟植株生长的抑制作用[28]。张璐研究发现,添加不同种类的生物炭均可提高烤烟的根系活力,缓解镉胁迫对根系生长的抑制作用,增加烤烟植株对镉胁迫的耐受性[29]。
气体交换参数是反映植物光合作用的指标之一。本研究结果表明,镉胁迫下添加生物炭显著提高了烤烟的净光合速率,其效果表现为A3>A1>A2>A0>A4处理。刘跃东等发现镉通过烤烟根系吸收进入植株体内后,一方面镉会直接毒害烤烟根系,另一方面镉会导致烤烟的营养平衡遭到破坏,引起代谢紊乱,减少叶片的同化面积,造成光合速率的下降,进而抑制烤烟的生长[30];同时,镉还会引起气孔水分丧失,导致气孔开度减少甚至关闭,CO2无法进入,固碳过程受阻,进而影响光合速率。本研究中净光合速率增加可能是由于生物炭降低了土壤镉元素的有效态,使得烤烟中富集的镉元素含量下降,烤烟生长受到的胁迫变小,使得净光合速率增加。
叶绿素和类胡萝卜素均有吸收光能的作用,且叶绿素是植物光合作用的关键色素之一。当植物遭受镉毒害时体内的叶绿素含量就会降低[31],在镉胁迫下大多数植物的叶绿素合成会受到抑制[32],由本研究结果可知,添加生物炭处理组的叶绿素含量较对照组减少,说明添加生物炭抑制了叶绿素的合成,可能是施用生物炭过量不利于植物生长和光合作用的提高,施加生物炭导致植物光合性能表现不同,进而导致烤烟叶片内的叶绿素含量下降,这与马丽等的研究结果[33]相同。
植物在逆境下会产生大量的活性氧(ROS),若不及时清除,会造成ROS的失衡。植物通过增加抗氧化酶活性等方式,提高清除ROS的能力,以抵御胁迫对植物细胞膜的伤害。SOD酶是重要的活性氧清除酶,能够反映出植物对于生物与非生物胁迫的抗性。SOD酶活性高低直接决定了植物的抗逆性[34-35]。本研究结果表明,镉处理增强了烤烟植株的抗氧化酶活性,而添加生物炭后SOD酶活性除A4处理增加了13.8%外,其余处理均有降低,POD酶活性都显著降低,主要是因为生物炭降低了土壤中有效镉含量,植物对镉的吸收减少[36],因此抗氧化酶活性有所下降。简敏菲等也发现施加生物炭后,镉胁迫下紫花地丁中的抗氧化酶活性也呈一定的下降趋势[37],这表明添加生物炭能够激活烤烟的应激机制,以抵抗镉污染胁迫,并且能够缓解镉对于烤烟的脂膜氧化过程,进而缓解了镉对于烤烟的胁迫作用,降低了抗氧化酶的活性。
施用生物炭能够降低烤烟对镉的富集和转运系数,减少对镉的吸收和积累,降低了烤烟中各部位的镉含量。这可能是因为生物炭在高温裂解状态下表面产生许多孔隙并且带有大量负电荷,会与土壤中的镉离子产生静电作用,进而起到吸附作用[38];其次,生物炭表面分布着很多含氧官能团 (如羧基和羟基),这些官能团能与Cd2+结合形成金属络合物,从而降低镉的迁移性和生物有效性[39]。但是不同裂解温度的生物炭其效果是不同的,本研究结果得出500 ℃的生物炭效果最好。
本研究结果表明,添加生物炭减缓了镉对烤烟根系的胁迫作用,促进了根系的生长,提高了根系的活力,增加了烤烟对镉胁迫的耐受性,提高了镉胁迫下烤烟的净光合速率。生物炭促进了镉胁迫下烤烟生物量的增加,其中裂解温度为700 ℃的生物炭作用最明显。添加还生物炭减少了烤烟的富集系数和转移系数,其中裂解温度500 ℃效果最好,可作为烤烟安全生产以及土壤镉修复的的有效措施。
参考文献:
[1]罗 勇,焦桂珍,刘胜波,等. 不同浓度镉对烟草幼苗生长发育及生长素相关基因表达的影响[J]. 中国农业科技导报,2021,23(1):58-65.
[2]Dias M C,Monteiro C,Moutinho-Pereira J,et al. Cadmium toxicity affects photosynthesis and plant growth at different levels[J]. Acta Physiologiae Plantarum,2013,35(4):1281-1289.
[3]Pinto F R,Mourato M P,Sales J R,et al. Oxidative stress response in spinach plants induced by cadmium[J]. Journal of Plant Nutrition,2017,40(2):268-276.
[4]张 晓,张环纬,陈 彪,等. 外源硅及水杨酸对镉胁迫下烟草幼苗生长和生理特性的影响[J]. 中国农业科技导报,2019,21(3):133-140.
[5]张建云,吴胜春,王敏艳,等. 烟秆炭修复重金属污染土壤的效应及对烟草生长的影响[J]. 浙江农林大学学报,2018,35(4):674-683.
[6]何 欣. 中国烟草概况[N]. 经济日报,2021-02-02(5).
[7]马思敏洁,刘殊捷,张轶源,等. 云南省烟草产业与经济发展的研究[J]. 财富时代,2019(8):154.
[8]Kuthanová A,Gemperlová L,Zelenková S,et al. Cytological changes and alterations in polyamine contents induced by cadmium in tobacco BY-2 cells[J]. Plant Physiology and Biochemistry,2004,42(2):149-156.
[9]蒋喜艳,张述习,尹西翔,等. 土壤-作物系统重金属污染及防治研究进展[J]. 生态毒理学报,2021,16(6):150-160.
[10]Smernik R J,Kookana R S,Skjemstad J O. NMR characterization of 13C-benzene sorbed to natural and prepared charcoals[J]. Environmental Science & Technology,2006,40(6):1764-1769.
[11]Jaiswal A K,Elad Y,Graber E R,et al. Rhizoctonia solani suppression and plant growth promotion in cucumber as affected by biochar pyrolysis temperature,feedstock and concentration[J]. Soil Biology and Biochemistry,2014,69:110-118.
[12]Ok Y S,Chang S,Gao B,et al. SMART biochar technology—a shifting paradigm towards advanced materials and healthcare research[J]. Environmental Technology & Innovation,2015,4:206-209.
[13]王艳红,李盟军,唐明灯,等. 稻壳基生物炭对生菜Cd吸收及土壤养分的影响[J]. 中国生态农业学报,2015,23(2):207-214.
[14]梁仲哲,齐绍武,淡俊豪,等. 生物炭对镉胁迫下烟草镉含量动态变化及土壤理化性质的影响[J]. 江苏农业科学,2018,46(1):56-59.
[15]阚正荣,马守田,祁剑英,等. 施用生物炭对冬小麦光合潜力和籽粒产量的影响[J]. 麦类作物学报,2019,39(6):719-727.
[16]陈 越,闫 晶,闫 寒,等. 钝化剂对镉污染土壤中烟草生长发育的影响[J]. 吉林农业大学学报,2019,41(6):719-726.
[17]刘桂华,吴科堰,秦 松,等. 不同钝化材料对白及吸收累积镉的影响[J]. 西南农业学报,2022,35(6):1401-1406.
[18]徐 敏,刘 君,阿衣古力·阿布都瓦依提.植物生理实验教学中叶绿素提取方法比较[J]. 实验科学与技术,2018,16(4):129-133.
[19]Stewart R R,Bewley J D. Lipid peroxidation associated with accelerated aging of soybean axes[J]. Plant Physiology,1980,65(2):245-248.
[20]何 冰,叶海波,杨肖娥. 铅胁迫下不同生态型东南景天叶片抗氧化酶活性及叶绿素含量比较[J]. 农业环境科学学报,2003,22(3):274-278.
[21]邹 琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2000.
[22]Knrzer O C,Burner J,Boger P. Alterations in the antioxidative system of suspension-cultured soybean cells (Glycine max) induced by oxidative stress[J]. Physiologia Plantarum,1996,97(2):388-396.
[23]Dai H P,Wei S H,Skuza L,et al. Phytoremediation of two ecotypes cadmium hyperaccumulator Bidens pilosa L. sourced from clean soils[J]. Chemosphere,2021,273:129652.
[24]王天顺,陈 伟,杨玉霞,等. 螯合剂处理下钻叶紫苑对土壤重金属的吸收富集效应[J]. 南方农業学报,2020,51(2):291-298.
[25]Li H,Li X,Xiang L,et al. Phytoremediation of soil co-contaminated with Cd and BDE-209 using hyperaccumulator enhanced by AM fungi and surfactant[J]. Science of the Total Environment,2018,613/614:447-455.
[26]邢 瑶,马兴华.氮素形态对植物生长影响的研究进展[J]. 中国农业科技导报,2015,17(2):109-117.
[27]黄玉芬,杜 毅,王钰静,等. 桑树枝和肉桂枝生物炭吸附及钝化土壤镉效果比较[J]. 广东农业科学,2022,49(3):96-105.
[28]吴春蕾. 生物炭修复技术在铅污染土壤及植物生长中的应用研究[J]. 聊城大学学报(自然科学版),2017,30(1):66-71,77.
[29]张 璐. 不同高碳物料对土壤理化性状、微生物多样性和烟株生长发育的影响[D]. 郑州:河南农业大学,2018.
[30]刘跃东,刘 祥,林 伟,等. 甲霜灵与镉复合污染对烟草生长发育及光合性能的影响[J]. 中国烟草科学,2019,40(1):41-48.
[31]Yue Z. Cadmium accumulation and antioxidative defenses in leaves of Triticum aestivum L.and Zea mays L.[J]. African Journal of Biotechnology,2011,10(15):2936-2943.
[32]Ozyigit I I,Vardar F,Yasar U,et al. Long-term effects of aluminum and cadmium on growth,leaf anatomy,and photosynthetic pigments of cotton[J]. Communications in Soil Science and Plant Analysis,2013,44(21):3076-3091.
[33]马 丽,郭学良,齐红志,等. 生物炭对连作草莓光合特性及光响应的影响[J]. 西北林學院学报,2020,35(2):72-78.
[34]王发展,金伊楠,李子玮,等. 干旱胁迫下外源ALA对烤烟幼苗光合特性和抗氧化能力的影响[J]. 中国烟草科学,2020,41(1):22-29.
[35]李雪君,吴 疆,吴家静,等. 烤烟新品种渠首1号对低氮胁迫的响应[J]. 江苏农业科学,2022,50(14):80-85.
[36]悦飞雪,李继伟,王艳芳,等. 施用秸秆生物炭和鸡粪对镉胁迫下玉米生长及镉吸收的影响[J]. 农业环境科学学报,2018,37(10):2118-2126.
[37]简敏菲,何旭芬,彭雨露,等. 生物炭对镉污染土壤中紫花地丁的生长及生理生态的影响[J]. 江西师范大学学报(自然科学版),2022,46(1):99-106.
[38]孙家婉,张振华,赵玉萍,等. 生物炭改性及其在农田土壤重金属修复中的应用研究进展[J]. 江苏农业科学,2022,50(10):9-15.
[39]Zeng G M,Wu H P,Liang J,et al. Efficiency of biochar and compost (or composting) combined amendments for reducing Cd,Cu,Zn and Pb bioavailability,mobility and ecological risk in wetland soil[J]. RSC Advances,2015,5(44):34541-34548.
