外源钙与甜菜碱对干旱胁迫下丝瓜幼苗的缓解效应
2023-05-23任晓雪王红军杨洪霞
任晓雪 王红军 杨洪霞

摘要:为探讨外源钙与甜菜碱提高丝瓜幼苗干旱胁迫抗逆性的作用机理,以丝瓜品种绿钻88为试材,在水培条件下借助PEG-6000模拟干旱胁迫环境,研究其对干旱胁迫下丝瓜幼苗的形态建成、光合色素、光合能力、保护酶活性、膜透性和活性氧积累的影响。结果表明:干旱胁迫能够抑制丝瓜幼苗的形态建成,外源钙与甜菜碱可在一定程度上缓解干旱胁迫对丝瓜幼苗生长的抑制作用,且以二者配施效果更佳。干旱胁迫能够显著降低丝瓜幼苗的叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素含量和光合特性,且降低程度随胁迫时间的后移而加剧,而外源钙与甜菜碱可在一定水平上缓解这种抑制作用,且以二者配施效果更佳。外源钙与甜菜碱能够提高干旱胁迫下丝瓜幼苗的叶绿素a/b值,且以二者配施的PCG处理下值最大。干旱胁迫能够显著降低丝瓜幼苗的保护酶活性,降低幅度随胁迫时间的后延而提高,而外源钙和甜菜碱能够减少干旱胁迫下保护酶活性的降低幅度,且这种减缓作用随胁迫时间的后延而更加显著,同时以二者配施效果更佳。干旱胁迫显著增加了丝瓜幼苗叶片MDA、O-2·、H2O2含量,提高叶片相对电导率,且随胁迫时间的后移呈升高趋势,引发氧化胁迫,而外源钙、甜菜碱能够缓解干旱逆境对丝瓜幼苗叶片细胞膜的氧化伤害,且以二者配施效果更佳。钙离子通道抑制剂LaCl3的施用表明,缺钙能够加重干旱胁迫对丝瓜幼苗形态建成和生理特性的抑制作用,钙对甜菜碱在增强植株抗旱性方面有正向促进作用。可见,外源钙与甜菜碱配施能够在一定水平上有效缓解干旱胁迫对丝瓜幼苗在生长、光合色素含量、光合能力、保护酶活性方面的抑制作用,减少干旱胁迫下丝瓜幼苗活性氧的积累,降低细胞膜膜脂过氧化水平,从而增强丝瓜幼苗的抗逆性。
关键词:外源钙;甜菜碱;干旱胁迫;丝瓜;缓解效应
中图分类号:S642.401 文献标志码:A
文章编号:1002-1302(2023)08-0131-08
基金项目:河南省“四优四化”科技支撑行动计划(编号:SYSH20200503004);中原学者工作站资助项目(编号:214400510023);2022年度河南省重点研发专项(编号:221111110100)。
作者简介:任晓雪(1982—),女,河南商丘人,助理研究员,主要从事植物逆境生理生态研究。E-mail:yguangwode@163.com。
通信作者:杨洪霞,高级农艺师,主要从事植物逆境生理生态研究。E-mail:yguangwode@163.com。
干旱作为一种极端天气时常出现,严重制约作物的正常生长和产量增加,是诸多农业发展所面临的主要瓶颈之一。全世界有约1/3的陆地面积为干旱或半干旱地区,而我国则有约51%的国土面积为干旱或半干旱地区[1]。植物在干旱条件下会表现出一系列表型和生理生化层面的变化,表型层面表现为生长变慢、生物量降低、叶片卷曲黄化[2],生理生化层面表现为叶绿素合成受阻、光系统Ⅱ(PSⅡ)反应中心对光能捕获和电子传递效率降低[3]、光合能力减弱、活性氧(ROS)代谢平衡紊乱[4]、膜结构损伤,最终导致作物产量和品质的下降。丝瓜富含维生素和矿物质,具有较高的食用和药用价值,具有广阔的消费市场。丝瓜的根系相对发达,但仍会遭受水分胁迫的危害。在育苗期,人为管理不当、断水断电等原因造成的水分脅迫,会严重威胁穴盘内丝瓜幼苗的生长。若移栽后遭遇水分胁迫,植株会因生长势的降低而变得瘦弱矮小。结果期受到水分胁迫,则会使丝瓜果实过早老化、纤维增加、品质降低、商品果减少。因此,研究丝瓜在干旱逆境下的响应机理,提高丝瓜抵御干旱逆境的能力,对减少生产损失有重要意义,对提高其他作物的抗逆性也有一定的参考价值。
钙是植物生长发育所必需的中量元素,同时是偶联胞外刺激和胞内生理生化反应的第二信使[5],对植物生长发育和生理代谢活动有重要调控作用[6]。在逆境条件下,钙参与逆境信号的响应与表达,对稳定细胞膜结构、阻止膜脂过氧化反应、提高抗氧化保护酶系统的酶活性、改善光合性能等方面有重要作用[7]。研究表明,叶片喷施或根灌外源钙均能够有效改善干旱胁迫下黄芩幼苗叶绿素的含量和保护酶的活性[8]。钙可促进水分胁迫下小麦幼苗的生长和养分吸收,缓解水分胁迫的抑制效应[9]。甜菜碱(glycine betaine,GB)广泛存在于动植物和微生物体内,是一种季胺型水溶性生物碱,能够减缓高温、干旱、盐害等逆境胁迫对植物造成的伤害[10]。在逆境条件下,甜菜碱可通过降低细胞渗透压、清除机体活性氧、维持生物膜结构的稳定性和生物体内大分子结构的完整性等方式,增强植株的抗逆性[11]。研究表明,甜菜碱能够维持干旱胁迫下茄幼苗的生长,并使MDA含量保持较低水平[12]。甜菜碱可提高干旱胁迫下甜茶的相对含水量和渗透调节物质含量,调节抗氧化酶保护系统的酶活性[13]。可见,外源钙和甜菜碱对提高作物抗旱性均有一定正向作用,而有关外源钙、甜菜碱单施或配施在丝瓜幼苗抗旱性方面的研究尚未见报道。为此,本试验以丝瓜品种绿钻88为试材,在水培条件下借助PEG-6000模拟干旱胁迫环境,研究外源钙、甜菜碱对干旱胁迫下丝瓜幼苗形态建成、光合色素、光合能力、保护酶活性、膜透性、活性氧积累的影响,探讨外源钙与甜菜碱提高丝瓜幼苗干旱胁迫抗逆性的作用机理,以期为丝瓜生产中缓解干旱逆境伤害和进行抗旱节水栽培提供理论依据。
1 材料与方法
1.1 供试材料
供试丝瓜品种为从瑞克斯旺公司引进的绿钻88。
1.2 试验设计
试验于2022年2月17日至5月20日,在河南省商丘市农林科学院双八试验站(115°64′E,34°50′N)智能温室内进行。将丝瓜种子放入55 ℃水中,用玻璃棒不断搅拌至常温,浸泡12 h,再用10%磷酸三钠溶液浸泡15~20 min消毒,然后用清水冲洗2~3遍,放置于催芽箱内进行催芽,待2/3的种子露白即可播种。将种子芽尖向下播种于32孔穴盘内,每穴1粒,上覆1~1.5 cm的基质。基质装盘前用800~1 000倍液的72.2%霜霉威水剂均匀喷洒1遍,以预防猝倒病、立枯病等苗期病害。待幼苗长至2叶1心时,选取长势基本一致的幼苗,洗去根部基质,移栽到装有1/2 Hoagland 营养液的栽培筐内,每筐栽培6株。栽培筐长×宽×高为40 cm×25 cm×15 cm。移栽至栽培筐2 d后,开始进行干旱胁迫试验。
在前期预备试验的基础上,筛选CaCl2浓度 14 mmol/L、甜菜碱(glycine betaine,GB)浓度1.2 mmol/L,选用10% PEG-6000模拟干旱胁迫。试验共设置6个处理:对照(CK):1/2 Hoagland 营养液+0叶面处理;P:10% PEG-6000+叶面喷施纯净水;PC:10% PEG-6000+14 mmol/L CaCl2;PG:10% PEG-6000+1.2 mmol/L甜菜碱;PCG:10% PEG-6000+14 mmol/L CaCl2+1.2 mmol/L甜菜碱;PLG:10% PEG-6000+1.0 mmol/L LaCl3(钙离子通道抑制剂)+1.2 mmol/L甜菜碱。每处理重复3次,每3框记作1次重复,共54框。在干旱试验开始前连续喷施3 d CaCl2,每天喷施1次,P处理在干旱试验开始前连续喷施3 d纯净水,甜菜碱、LaCl3(钙离子通道抑制剂)在干旱试验开始前 1 d 喷施,喷施时间均为上午08:00—09:00。在干旱胁迫试验的第3天测定丝瓜幼苗生理指标,在第10天测定形态指标和生理指标。
1.3 测定指标与方法
用直尺测量株高和根长,游标卡尺测量茎粗,叶面积仪测量叶面积,LA-S2400根系扫描仪测量根尖数,千分之一电子天平称量干鲜质量。采用乙醇丙酮浸提法[14]测定叶绿素含量。使用便携式光合仪Li-6400(美国,LI-COR公司生产)测定第2张真叶净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。参照王学奎等的方法[15]测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性。采用沈文飚等的方法[16]测定抗坏血酸过氧化物酶(APX)活性。参照赵世杰等的方法[17]测定MDA含量、相对电导率。采用郭允娜等的方法测定超氧阴离子(O-2·)和过氧化氢(H2O2)含量[18]。
1.4 数据处理
使用Microsoft Excel 2007进行数据处理和作图,利用SPSS 26.0软件对数据进行方差分析及差异性检验(Duncans法,α=0.05)。
2 结果与分析
2.1 外源钙与甜菜碱对干旱胁迫下丝瓜幼苗形态建成的影响
由表1可知,与CK相比,干旱胁迫能够显著降低西瓜幼苗的株高、茎粗、叶面积、根长、根尖数、鲜质量、干质量,这说明干旱胁迫能够抑制丝瓜幼苗形态建成。与单纯的干旱胁迫P处理相比,PC、PG处理显著提高了丝瓜幼苗的株高、茎粗、叶面积、根长、根尖数、鲜质量和干质量,这表明外源钙和甜菜碱叶面喷施均能够有效缓解干旱逆境胁迫对丝瓜幼苗生长的抑制作用。PCG处理下丝瓜幼苗各形态指标均高于PC、PG处理,其中在株高、根尖数、鲜质量指标上与PC、PG处理差异显著(P<0.05),PCG较PC处理分别增加9.98%、12.47%、8.25%,PCG较PG处理分别增加8.09%、23.62%、9.95%。添加有钙离子通道抑制剂和甜菜碱的PLG處理,其各指标均显著低于PG处理,且在株高、叶面积、根长、鲜质量、干质量指标上与P处理差异显著(P<0.05)。说明在钙缺乏的前提下,甜菜碱对增强植株抗旱性的促进作用会受到抑制。
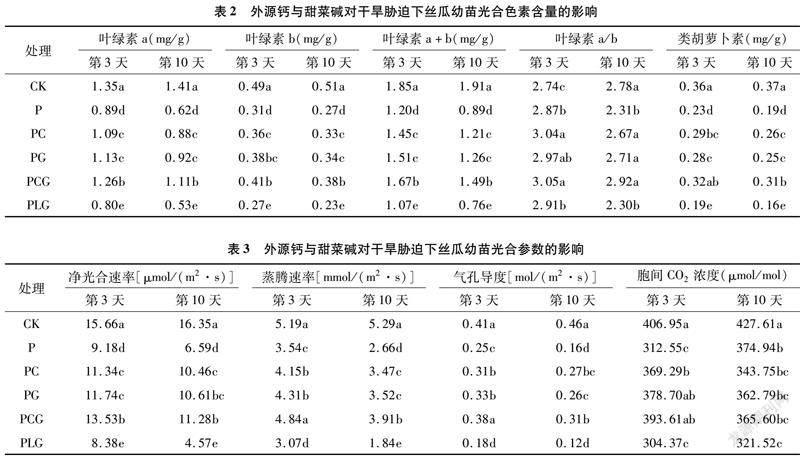
2.2 外源钙与甜菜碱对干旱胁迫下丝瓜幼苗光合色素含量的影响
由表2可知,干旱胁迫能够显著降低丝瓜幼苗叶片的叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素含量,其含量随胁迫时间的延长呈下降趋势。在干旱胁迫3 d时,PC、PG、PCG处理的丝瓜幼苗叶片的叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素含量均高于P处理;其中以PCG处理下值最大,较P处理分别增加41.57%、32.26%、39.17%、39.13%。表明叶面喷施外源钙、甜菜碱能够在一定水平上阻止干旱胁迫下丝瓜叶片光合色素含量的降低幅度,且外源钙与甜菜碱复配效果更佳。PC、PG、PCG处理的丝瓜幼苗各指标在第10天时的变化规律与第3天时一致,且均低于第3天,但PCG处理的叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素降幅均低于PC、PG处理,分别比第3天降低11.90%、7.32%、10.78%、3.13%,表明PCG处理更有利于延缓长时间干旱胁迫环境下光合色素含量的降低速率。PLG处理下各色素指标均低于PG、PLG处理,这同时也说明钙的添加能够促进甜菜碱对干旱胁迫的缓解效应。另外,在5个干旱处理下,以PCG处理下的叶绿素a/b值最大。
2.3 外源钙与甜菜碱对干旱胁迫下丝瓜幼苗光合参数的影响
由表3可知,5个干旱处理的净光合速率、蒸腾速率、气孔导度、胞间CO2浓度均低于对照,其中净光合速率以P、PLG处理的降幅相对较大,PC、PG、PCG降幅相对较小,说明外源钙、甜菜碱单施或配施均能减轻干旱胁迫对丝瓜幼苗光合能力的抑制作用。PC、PG处理的净光合速率均高于P处理,虽然PG高于PC,但二者差异不显著;PCG处理在胁迫第3天时显著高于PC、PG处理,在胁迫第10天显著高于PC处理,高于PG处理但与其差异不显著,表现出一定的补偿效应,这说明外源钙和甜菜碱复配较单一外源物质效果更佳,且随着时间的推移,能够更好地延缓丝瓜幼苗光合能力的降低速率。另外,在胁迫第3天和第10 天,PCG处理的净光合速率较P处理分别增加47.39%、71.17%,与P处理差异显著(P<0.05)。PLG处理的净光合速率不仅低于PG处理,还低于P处理,说明钙的缺乏能够显著抑制丝瓜幼苗光合能力的提高。从胁迫第3天至第10天,PC、PG、PCG处理的净光合速率、蒸腾速率、气孔导度均表现为下降趋势,而胞间CO2浓度亦表现为下降趋势;在P、PLG处理下,净光合速率、蒸腾速率、气孔导度表现为下降趋势,而胞间CO2浓度却呈现升高趋势。
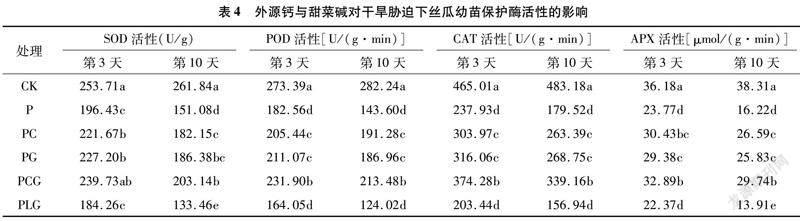
2.4 外源钙与甜菜碱对干旱胁迫下丝瓜幼苗保护酶活性的影响
由表4可以得出,单纯的干旱胁迫P处理显著降低了丝瓜幼苗叶片抗氧化保护酶的活性,其中SOD、POD、CAT、APX活性在胁迫第3天较CK分别降低22.58%、33.22%、48.83%、34.30%,胁迫第10天较CK分别降低42.30%、49.12%、62.85%、57.66%。外源钙、甜菜碱单施或复配施用均能够有效缓解干旱胁迫对丝瓜幼苗叶片保护酶活性的抑制作用,PC、PG、PCG处理在干旱胁迫第3天和第10天,其保护酶活性均显著高于P处理,且以PCG处理的保护酶活性相对较高。PCG处理的SOD、POD、CAT、APX活性在胁迫第3天较P处理分别增加22.04%、27.03%、57.31%、38.37%,而在第10天较P处理分别增加34.46%、48.66%、88.93%、83.35%,增加幅度明显高于第3天,表明随着时间的后移,外源钙和甜菜碱复配对干旱胁迫下保护酶活性降低幅度的缓解作用更加显著。在胁迫第3天和第10天,PLG处理保护酶活性均低于PG、PCG,说明外源钙对甜菜碱缓解干旱胁迫有正向促进作用。
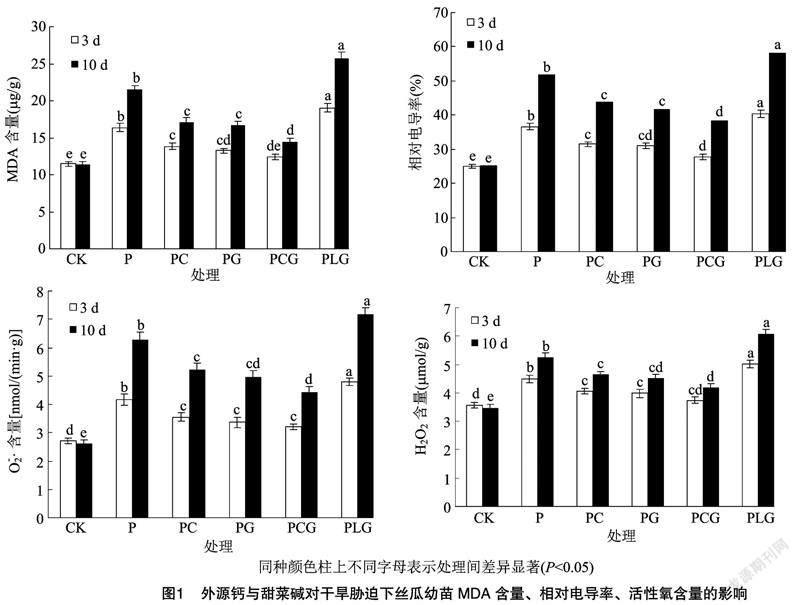
2.5 外源钙与甜菜碱对干旱胁迫下丝瓜幼苗MDA、相对电导率和活性氧含量的影响
由图1可以看出,P处理下的MDA含量、相对电导率、O-2·含量、H2O2含量均显著高于对照,且随着干旱时间的后移呈升高趋势,表明干旱胁迫对丝瓜幼苗细胞造成一定程度的伤害,且伤害程度随胁迫时间的增加而加重。与P处理相比,PC、PG、PCG处理下MDA含量、相对电导率、O-2·含量、H2O2含量均显著降低,且以PCG处理的降低幅度最大,说明外源钙、甜菜碱能够缓解干旱逆境对丝瓜幼苗的伤害,且以二者复配效果更佳。而PC、PG、PCG处理的MDA含量、相对电导率、O-2·含量、H2O2含量仍高于对照,说明外源钙、甜菜碱对干旱胁迫的缓解效应存在一定的限度。PLG处理的MDA含量、相对电导率、O-2·含量、H2O2含量显著高于PC、PCG处理,说明钙对甜菜碱缓解干旱胁迫方面有促进作用。
3 讨论与结论
干旱是影响植物生长发育的主要非生物胁迫因子之一。干旱胁迫下植株生长速度放缓、株形矮小、叶面积减小、生物量积累减少、产量降低。本研究结果表明,与对照CK相比,干旱胁迫能够显著降低丝瓜幼苗的株高、茎粗、叶面积、根长、根尖数、鲜质量、干质量。这是因为,植物生长依赖细胞的分裂分化,但在干旱胁迫下,细胞失水膨压降低,分生组织细胞有丝分裂速度变慢、细胞生长受到抑制,使得植物表现为生长减弱[19]。钙是植物代谢发育的主要调控者,对生长发育的各个时期均有调控作用。研究也表明,外源钙的施用能够缓解干旱、冷害等逆境胁迫对植株的伤害[20]。高向阳等研究发现,在0~14 mmol/L范围内,Ca2+浓度的增加能够促进植株根系生长及干物质积累[21]。山仑等研究得出,添加外源钙能促进小麦根茎叶的生长,并使小麦增长速度保持在较高水平[22]。本试验结果表明,在添加外源钙的PC处理下,丝瓜幼苗各形态指标虽低于CK,但均显著高于单纯的干旱胁迫P处理,说明喷施外源钙能够有效消弱干旱对丝瓜幼苗生长的抑制作用,这应该与钙是细胞信号转导的第二信使有关,钙与钙调蛋白(CaM)结合后,将感受到的外界干旱胁迫信号传递下去;同时,钙参与信号的响应和表达,降低逆境胁迫给细胞膜带来的伤害,进而增加植株的抗旱能力[23]。另外,钙是构成细胞壁的重要元素,外源钙的添加能够促进细胞壁的形成,促进细胞分裂生长。甜菜碱是重要的渗透调节物质,并在生物界广泛存在,而外源甜菜碱易被植株吸收并能够在植物体内长时间存在。研究也表明,甜菜碱能够提高植物的抗逆性[24]。本试验结果表明,添加有甜菜碱的PG处理丝瓜幼苗各形态指标均高于P处理,这应该是因为渗透胁迫引起的细胞膨压降低,对植株生长不利,而外源甜菜碱的添加能够在一定水平上通过渗透调节提高植株生长所需要的膨压,促进植株生长[25]。PCG处理下丝瓜幼苗各形态指标均高于单施外源钙或甜菜碱的干旱处理,这说明外源钙与甜菜碱配施效果更佳。PCG处理各指标均高于单施甜菜碱的PG处理,而含有钙离子通道抑制剂和甜菜碱的PLG处理各指标均低于PG处理,说明钙的缺失会制约甜菜碱在增强植株抗旱性方面的作用,而外源钙的添加能够提高甜菜碱对丝瓜幼苗干旱胁迫的缓解作用,同时也从侧面说明外源钙与甜菜碱配施更有利于提高植株的抗旱能力。
叶绿素是绿色植物进行光合作用的主要场所,其含量与叶片光合速率、碳同化能力密切相关,但叶绿素对干旱逆境反应敏感,因此是表征植株抗逆性的重要指标[26]。本试验结果表明,在单纯的干旱胁迫下,叶片叶绿素含量较对照均显著降低,这是因为叶绿素在缺水条件下合成受到抑制,同时加速已有叶绿素的分解[27]。在干旱条件下,添加外源钙或甜菜碱能够提高光合色素含量,试验结果与郭郁频等在苜蓿[28]、范春丽等在石榴[29]上的研究结论一致。一方面是因为,Ca2+对叶绿体双层膜的稳定有保护作用,甜菜碱能够减轻逆境胁迫对叶绿体类囊体膜的伤害,对光合色素有一定的保护作用[30];另一方面是因为,活性氧(ROS)不但能够对细胞结构造成伤害,还可能参与对叶绿素4-吡咯环结构的破坏[31]。而在本试验条件下,外源钙或甜菜碱的添加降低了干旱胁迫处理超氧阴离子等ROS含量,这在一定水平上减少了ROS对叶绿素的破坏。外源钙和甜菜碱复配后,较外源钙和甜菜碱单施更能够提高干旱胁迫下丝瓜幼苗叶绿素的含量,说明二者配施效果更佳。PC、PG、PCG处理下,叶绿素a/b值均高于P处理,且以PCG处理下值最大,这应该是因为超氧阴离子能够对叶绿素造成伤害,并特异性地对叶绿素a造成破坏[32],而PCG处理下超氧阴离子含量较其他干旱处理低,叶绿素a受到的破坏程度相对较轻。
光合作用是植物进行物质代谢和能量代谢的前提,光合能力的高低除受自身遗传因素影响外,还受干旱、高温、低温等外界环境因素的制约,因此常被作为衡量植株抗逆性的重要参考指标。植物在遭受干旱逆境胁迫时,气孔阻力增加,CO2交换速率和光合酶活性下降,净光合速率降低,干物质积累减少,植物生长受到抑制[33]。在干旱胁迫下,植物光合速率的降低受气孔因素和非气孔因素的影响。本试验条件下,P处理和PLG处理在干旱胁迫第3天时,净光合速率、气孔导度、胞间CO2浓度均显著低于对照,表现为净光合速率随气孔导度、胞间CO2浓度的降低而降低,说明在胁迫第3天,P、PLG处理光合速率的降低主要是由气孔因素限制引起。与胁迫第3天相比,胁迫第10天的P、PLG处理净光合速率、气孔导度虽有所降低,但胞间CO2浓度却呈不降反升的趋势,这说明在干旱胁迫的后期,P、PLG处理净光合速率的降低已由气孔因素限制为主转变为非气孔因素限制为主,试验结果与郭爱霞等在苹果砧木上的研究结论[34]一致。这应该是因为,干旱胁迫时间的延长导致活性氧大量积累,使细胞膜受到破坏,最终引起光合细胞机构与功能的损伤。從胁迫第3天至第10天,PC、PG、PCG处理下净光合速率、气孔导度、胞间CO2浓度均呈下降趋势,且显著低于对照,说明气孔因素限制是引起净光合速率降低的主要因素;同时,还表明外源钙、甜菜碱的叶面喷施,能够延缓干旱胁迫下光合作用由气孔因素限制向非气孔因素限制转变的过程,增强植株的抗旱性。外源钙提高丝瓜幼苗的净光合速率,应该是因为钙有利于细胞膜结构的完整性,提高膜上酶的活性和叶肉细胞CO2羧化效率;同时,钙能增加气孔反应的灵敏性和自我调节能力,使叶片保持相对较高的含水量[35],有利于光合作用的提高。另外,植物遭受干旱逆境时,外源钙可通过调控 PSⅡ中心活性来优化细胞叶绿素荧光能量分配,从而改善植物的光合性能[36]。甜菜碱可通过保持PSⅡ反应中心结构的完整性和1,5-二磷酸核酮糖羧化/加氧酶(Rubisco)活性,提高叶片的光合速率[37],有利于缓解逆境条件下叶片光合能力的降低幅度。
SOD、POD、CAT、APX是抗氧化保护酶系统中的主要酶类,这些酶相互协同,使机体活性氧的产生与消除保持动态平衡,其中SOD可有效清除破坏性较强的超氧阴离子自由基,并将其转化成H2O2,POD、CAT、APX可将由超氧阴离子转化而来的H2O2分解成完全无害的水[38]。另外,APX还能催化抗坏血酸AsA的氧化,减轻自由基对机体的危害。本试验结果表明,干旱胁迫显著降低了丝瓜幼苗叶片保护酶的活性,这是因为保护酶系统对细胞内活性氧的清除存在一定的阈值,当活性氧的积累超出这一阈值时,细胞受到活性氧伤害,保护酶活性降低[39]。钙信使系统与抗氧化保护酶系统关系密切。本试验条件下,外源钙的添加提高了干旱胁迫下丝瓜叶片保护酶的活性,试验结果与谢诗在结缕草上的研究结论[40]一致。这应该是因为钙离子与CAM等受体结合后,通过对信号的放大和传递,降低活性氧的积累,增强了抗氧化酶活性[41]。甜菜碱同样提高了干旱条件下丝瓜叶片保护酶的活性,试验结果与马千全等在小麦上的研究结论[42]一致。这可能与3个方面原因有关:一是甜菜碱通过调控抗氧化酶相关基因在逆境条件下的表达,减轻抗氧化酶保护系统遭受的逆境损伤[43];二是甜菜碱在逆境条件下能够稳定保护酶的结构和功能,减轻渗透胁迫对蛋白质结构和功能的伤害,从而维持酶的活性;三是甜菜碱的亲水性可将植物组织间的水分聚集在蛋白质表面,提高了其表面的水合程度,从而减轻逆境胁迫引起的酶失活[44]。PCG处理下丝瓜幼苗叶片保护酶活性高于PC、PG处理,说明外源钙与甜菜碱具有一定的互作效应,二者配施比单一施用更有利于增强丝瓜幼苗的抗旱能力。
氧化胁迫是干旱胁迫引发的最主要的次级胁迫,它通过活性氧代谢的失衡在细胞水平上对植物造成伤害。MDA含量和相对电导率常用于反映逆境胁迫对细胞的伤害程度,是植物应激反应的重要指标。本试验结果表明,单纯的干旱胁迫显著提高了丝瓜幼苗MDA含量和相对电导率,数值随胁迫时间的延长而增大,这是因为干旱条件下,O-2·、H2O2在丝瓜幼苗细胞内大量积累,活性氧(ROS)积累达到一定阈值,使细胞膜发生膜脂过氧化反应,而MDA是该反应的最终产物之一,因此表现出干旱胁迫下MDA含量随ROS的增加而增加的现象。膜脂过氧化反应发生后,细胞膜透性增大,相对电导率随之增加。增施外源钙后,丝瓜幼苗MDA含量和相对电导率比单纯干旱胁迫处理显著降低,这应该是因为钙是植物细胞膜的重要组成成分,是生物膜的稳定剂,钙与膜磷脂的极性头部发生交联作用,使膜蛋白与磷脂结合更加紧密,这也使得膜的透性降低[45],从而降低了干旱条件下丝瓜幼苗MDA含量和相对电导率。甜菜碱的施用同样能够降低丝瓜幼苗MDA含量和相对电导率,这应该是因为甜菜碱能够抑制膜脂过氧化反应,有利于维持细胞膜结构的完整性,从而提高机体的抗逆性,进而降低了O-2·、H2O2等活性氧的过量积累[46]。外源钙与甜菜碱配施较单一施用更加降低了丝瓜幼苗MDA含量、相对电导率、O-2·含量、H2O2含量,说明二者配施对增强植物抗逆性有正向促进作用。
参考文献:
[1]李承业,王燕飞,黄 润,等. 我国甜菜抗逆性研究进展[J]. 中国糖料,2010,32(1):56-58.
[2]王 齐,孙吉雄,安 渊. 水分胁迫对结缕草种群特征和生理特性的影响[J]. 草业学报,2009,18(2):33-38.
[3]张会慧,张秀丽,许 楠,等. 外源钙对干旱胁迫下烤烟幼苗光系统Ⅱ功能的影响[J]. 应用生态学报,2011,22(5):1195-1200.
[4]安玉艳,梁宗锁. 植物应对干旱胁迫的阶段性策略[J]. 应用生态学报,2012,23(10):2907-2915.
[5]姜义宝,李建华,方丽云,等. 钙处理对苜蓿幼苗抗旱性的影响[J]. 中国草地学报,2008,30(1):117-120.
[6]周双云,蒋 晶,高龙燕,等. 不同浓度CaCl2对盐胁迫下巴西蕉幼苗生理的影响[J]. 应用与环境生物学报,2014,20(3):449-454.
[7]陈立松,刘星辉. 渗透胁迫下Ca2+对龙眼叶片光合色素及膜脂过氧化的影响[J]. 园艺学报,1998,25(1):87-88.
[8]李小玲,华智锐,何娇娇.干旱条件下外源钙对商洛黄芩生理特性的影响[J]. 江西农业学报,2019,31(4):31-35.
[9]周 芳,赵玉霞,王文岩,等. 局部根区水分胁迫下钙对冬小麦生长及养分吸收的影响[J]. 干旱地区农业研究,2015,33(1):14-19,71.
[10]张红敏,李姣姣,黑刚刚,等. 外源甜菜碱处理对干旱胁迫下半夏氮代谢及相关酶活性的影响[J]. 草业学报,2014,23(4):229-236.
[11]Robinson S P,Jones G P. Accumulation of glycinebetaine in chloroplasts provides osmotic adjustment during salt stress[J]. Functional Plant Biology,1986,13(5):659.
[12]柴文臣,閻世江. 甜菜碱对干旱胁迫下茄子幼苗生长及生理指标的影响[J]. 中国瓜菜,2021,34(8):78-83.
[13]李爱梅,张 玲,张 超,等. 黄腐酸和甜菜碱预处理对干旱胁迫下平邑甜茶生理特性及光合的影响[J]. 西北植物学报,2017,37(2):307-314.
[14]白宝璋. 植物生理学(上):理论教程[M]. 北京:中国农业科技出版社,1996:65-67.
[15]王学奎,黄见良. 植物生理生化试验原理与技术[M]. 3版.北京:高等教育出版社,2015:168-181.
[16]沈文飚,徐朗莱,叶茂炳,等. 抗坏血酸过氧化物酶活性测定的探讨[J]. 植物生理学通讯,1996,32(3):203-205.
[17]赵世杰. 植物生理学试验指导[M]. 北京:中国农业科技出版社,1998:84-85,98-99.
[18]郭允娜,李衍素,贺超兴,等. 亚适宜温光下萘乙酸钠对番茄幼苗生长与生理特性的影响[J]. 应用生态学报,2015,26(10):3053-3058.
[19]Hussain M,Malik M A,Farooq M,et al. Improving drought tolerance by exogenous application of glycinebetaine and salicylic acid in sunflower[J]. Journal of Agronomy and Crop Science,2008,194(3):193-199.
[20]Kudla J,Batistic O,Hashimoto K.Calcium signals:the lead currency of plant information processing[J]. The Plant Cell,2010,22(3):541-563.
[21]高向阳,许志强,徐凤彩. 水分胁迫下钙对大豆叶片膜脂过氧化程度的影响[J]. 华南农业大学学报,1999,20(3):67-71.
[22]山 仑,郭礼坤,徐 萌,等. 干旱条件下钙与赤霉素混合处理种子的生理效应及增产效果[J]. 干旱地区农业研究,1994,12(1):85-91.
[23]Males J,Griffiths H. Stomatal biology of CAM plants[J]. Plant Physiology,2017,174(2):550-560.
[24]景蕊蓮,昌小平,胡荣海. 外源甜菜碱对小麦幼苗抗旱性的影响[J]. 干旱地区农业研究,1998,16(2):1-5.
[25]Okazaki Y,Tazawa M. Calcium ion and turgor regulation in plant cells[J]. The Journal of Membrane Biology,1990,114(3):189-194.
[26]倪 锋,谢鹏飞,褚荣浩,等. 利用日光诱导叶绿素荧光估算高温干旱复合胁迫下夏玉米生理生态参数[J]. 江苏农业学报,2022,38(3):587-596.
[27]薛 崧,汪沛洪,许大全,等. 水分胁迫对冬小麦CO2同化作用的影响[J]. 植物生理学报,1992,18(1):1-7.
[28]郭郁频,任永霞,刘贵河,等. 外源钙和赤霉素对干旱胁迫下苜蓿幼苗生理特性的影响[J]. 草业学报,2015,24(7):89-96.
[29]范春丽,罗 青. 干旱胁迫下外源甜菜碱对石榴光合作用、渗透调节及保护酶活性的影响[J]. 江苏农业科学,2016,44(11):229-232.
[30]赵新西,马千全,杨兴洪,等. 根施甜菜碱对干旱胁迫下小麦幼苗类囊体膜组分和功能的影响[J]. 植物生理与分子生物学学报,2005,31(2):135-142.
[31]蒋明义,杨文英,徐 江,等. 渗透胁迫下水稻幼苗中叶绿素降解的活性氧损伤作用[J]. 植物学报,1994,36(4):289-295.
[32]伍泽堂. 超氧自由基与叶片衰老时叶绿素破坏的关系(简报)[J]. 植物生理学通讯,1991,27(4):277-279.
[33]杨肖华,郭圣茂,冯美玲,等. 干旱胁迫及复水对射干光合作用和叶绿素荧光特性的影响[J]. 江西农业大学学报,2018,40(3):525-532.
[34]郭爱霞,石晓昀,王延秀,等. 干旱胁迫对3种苹果砧木叶片光合、叶绿体超微结构和抗氧化系统的影响[J]. 干旱地区农业研究,2019,37(1):178-186.
[35]任丽花,翁伯琦,方金梅. 施钙增强作物抗旱力的研究进展[J]. 亚热带农业研究,2005,1(3):19-25.
[36]李海霞,米银法,陈双臣. 外源钙对干旱胁迫下不同抗性牡丹PSⅡ功能和光能分配的影响[J]. 江苏农业科学,2022,50(16):120-127.
[37]Xing W B,Rajashekar C B. Alleviation of water stress in beans by exogenous glycine betaine[J]. Plant Science,1999,148(2):185-192.
[38]姜义宝,杨玉荣,郑秋红. 外源一氧化氮对干旱胁迫下苜蓿幼苗抗氧化酶活性和叶绿素荧光特性的影响[J]. 干旱地区农业研究,2008,26(2):65-68.
[39]何 玮,蒋 安,王 琳,等. PEG干旱胁迫对红三叶抗性生理生化指标的影响研究[J]. 中国农学通报,2013,29(5):5-10.
[40]谢 诗. 外源钙对干旱胁迫下结缕草生长的缓解作用[D]. 沈阳:辽宁大学,2013:1-49.
[41]Grlach A,Bertram K,Hudecova S,et al. Calcium and ROS:a mutual interplay[J]. Redox Biology,2015,6:260-271.
[42]马千全. 外源甜菜碱提高小麦抗旱性的研究[D]. 泰安:山东农业大学,2003.
[43]Einset J,Nielsen E,Connolly E L,et al. Membrane-trafficking RabA4c involved in the effect of glycine betaine on recovery from chilling stress in Arabidopsis[J]. Physiologia Plantarum,2007,130(4):511-518.
[44]郭启芳,马千全,孙 灿,等. 外源甜菜碱提高小麦幼苗抗盐性的研究[J]. 西北植物学报,2004,24(9):1680-1686.
[45]Abdel-Basset R. Calcium channels and membrane disorders induced by drought stress in Vicia faba plants supplemented with calcium[J]. Acta Physiologiae Plantarum,1998,20(2):149-153.
[46]刘金兰. 外源钙和甜菜碱提高小麦抗旱性研究[D]. 泰安:山东农业大学,2009:1-73.
