碱胁迫下内生真菌对宿主布顿大麦显微结构的影响
2023-03-05户萌菲刘小珍陈水红
龙 凤, 户萌菲, 陈 胜, 刘小珍, 陈水红
(塔里木大学生命科学与技术学院/塔里木盆地生物资源保护利用兵团重点实验室, 新疆 阿拉尔 843300)
目前,大多数关于作物胁迫的研究主要集中在盐胁迫上,盐碱混合胁迫研究相对较少。然而,在自然界中,高浓度的盐和高pH值经常同时发生,它们的协同作用可能比单独使用任何一种胁迫的影响对植物生长和发育更有害[1]。植物受到碱胁迫时,通常会改变自身根茎叶形态结构和生理过程去适应环境。植物主要通过表皮的增厚、叶片肉质加厚、叶片缩小和改变浓密短柔毛的覆盖等变化来增强对环境的适应能力[2]。有研究表明,能够在高浓度碱胁迫下生长的耐盐植物,是由长期的进化过程中通过改变特定的形态组织结构来适应逆境,降低盐离子带来的伤害[3]。了解禾草植物如何改变形态结构来应对碱胁迫带来的伤害,有助于为后期在饲用牧草和盐碱地的改良方面提高理论基础。
禾草内生真菌是指在宿主禾草体内度过部分或者全部生命周期,但不引起宿主植物任何症状的一类真菌[4]。目前,关于禾草内生真菌的研究主要集中在子囊菌门(Ascomycota)麦角菌科(Clavicipitaceae)的香柱菌(Epichloё)属[5]。在盐碱胁迫下,禾草内生真菌的侵染能够提高宿主植物的对外界环境的耐受性和适应性[6-8]。布顿大麦(Hordeumbogdanii)是一种耐盐植物,它具有较强的生态适应性以及竞争优势,在作为饲用牧草和荒漠及盐碱地改良草种等方面具有广阔的应用价值[9]。目前关于布顿大麦-内生真菌共生体的研究主要集中在生长发育[10-13]、生理生化[12,14]、营养价值[15]等方面,但该共生体在非生物胁迫下细胞显微结构的响应研究较少。因此,为了探究内生真菌侵染对宿主布顿大麦在碱胁迫条件下的显微结构是否有影响,本研究运用石蜡切片技术观察内生真菌侵染对布顿大麦根、茎、叶在不同碱处理下各个显微结构的变化,为内生真菌提高宿主布顿大麦耐碱性的生理机制提供形态学基础。
1 材料与方法
1.1 试验材料
实验室前期处理得到的温宿县E+与E-布顿大麦种子[15],并鉴定种子中带有内生真菌属于Epichloё属[16]后,将种植在塔里木大学动科试验站的试验田(80°76′E,41°58′N,海拔1 514 m)繁种子,收获种子作为试验的材料。分别挑取E+,E-饱满的种子进行清水萌发,长出子叶后,移栽入4 L水培盒中,每盆19孔,每孔1个苗。每盆连接一个充氧泵,每天定时进行换气,每3 d更换一次1/2 Hoagland营养液。温室的昼/夜温度为(28±2)℃/(15±3)℃,日照时长约为12 h。待布顿大麦分蘖3~5株时,用李春杰等[17]的茎髓苯胺蓝镜检法检测布顿大麦是否含有内生真菌,不带菌的为E-,带菌为E+。
1.2 碱胁迫处理
混合碱(Na2CO3∶NaHCO3=1∶1)处理,浓度梯度为50 mmol·L-1,100 mmol·L-1,每个处理重复4次,为了减少碱冲击效应,每12 h递增50 mmol·L-1碱(将前一次的碱溶液换成增加后的溶液),直到所有处理达到预定浓度,未加混合碱的处理作为试验的对照,每7 d更换一次对应浓度的混合碱溶液,至21 d取样,进行后期指标测定。
1.3 显微切片制作与观察
将各碱处理E+和E-布顿大麦,分别根(分生区)、茎(根颈)、叶(中央)剪成约1 cm小段,放装有70% FAA(70%酒精∶冰醋酸∶甲醛=90∶5∶5)青霉素小瓶中,然后用注射器抽取瓶中空气使样品沉入瓶底固定24 h。参照黄倩[18]的石蜡切片制作方法,固定完成后将材料取出进行脱水、透明、浸蜡、包埋、切片(厚度为10 μm)、粘片、脱蜡、染色(番红-固绿双重染色)、封片(加拿大树胶,solarbio公司CAS:8007-47-4),最后用研究级正置生物显微镜(日本,尼康Nikon Eclipse Ci-L)观察,利用NIS-Elements软件进行拍照并测量根的维管束面积、表皮厚度、内皮层厚度和导管直径,测量茎的维管束面积、气腔直径和表皮厚度,测量叶片的维管束面积、泡状细胞面积、气孔直径和导管直径。每个样品测量3个样本,每个样本测量8个视野,以平均值表示。
1.4 统计分析
采用SPSS统计软件(IBM SPSS Statistics 26)进行数据分析,图中数据为平均值±标准误差。采用双因素方差分析方法检测碱处理和内生真菌对布顿大麦根茎叶显微结构的影响,采用单因素ANOVA方差分析法检测不同碱浓度处理对E+和E-布顿大麦根茎叶显微结构的影响。采用独立样本T检验来检测同一碱浓度下E+和E-根茎叶显微结构的影响。统计显著性差异定义在95%置信水平。
2 结果与分析
2.1 碱胁迫对布顿大麦植株根的显微结构影响
由图1可知,布顿大麦根部结构主要由维管束、内皮层和表皮组成。碱胁迫对布顿大麦根的细胞结构影响较为明显。随着碱浓度的升高,根部细胞外皮层从50 mmol·L-1开始出现皱缩和破裂,到100 mmol·L-1处根部细胞外表皮层皱缩加深。另外,E+植株根中的导管数明显高于E-植株。由表1可知在0 mmol·L-1时维管束面积E+显著高于E-(P<0.05);碱处理后布顿大麦的维管束面积在缩小,在50 mmol·L-1处E+与E-的维管束面积差异不显著,但在100 mmol·L-1处E+的维管束面积显著高于E-(P<0.05)。碱胁迫导致对根的导管直径增加,E+的导管直径长于E-。碱胁迫对根的表皮厚度的影响,在无碱处理下表皮厚度E+与E-无显著差异,但在碱胁迫下E+的表皮厚度显著高于E-;同时,布顿大麦内皮层随着碱浓度的升高植株根内皮层在不断加厚,在50 mmol·L-1处E+显著高于E-的(P<0.05)。双因素方差分析后,发现内生真菌和碱处理显著影响维管束面积、导管直径、表皮厚度和内皮层厚度;除了导管直径以外,上述指标的内生真菌与碱处理两者之间存在交互作用(P<0.05)。
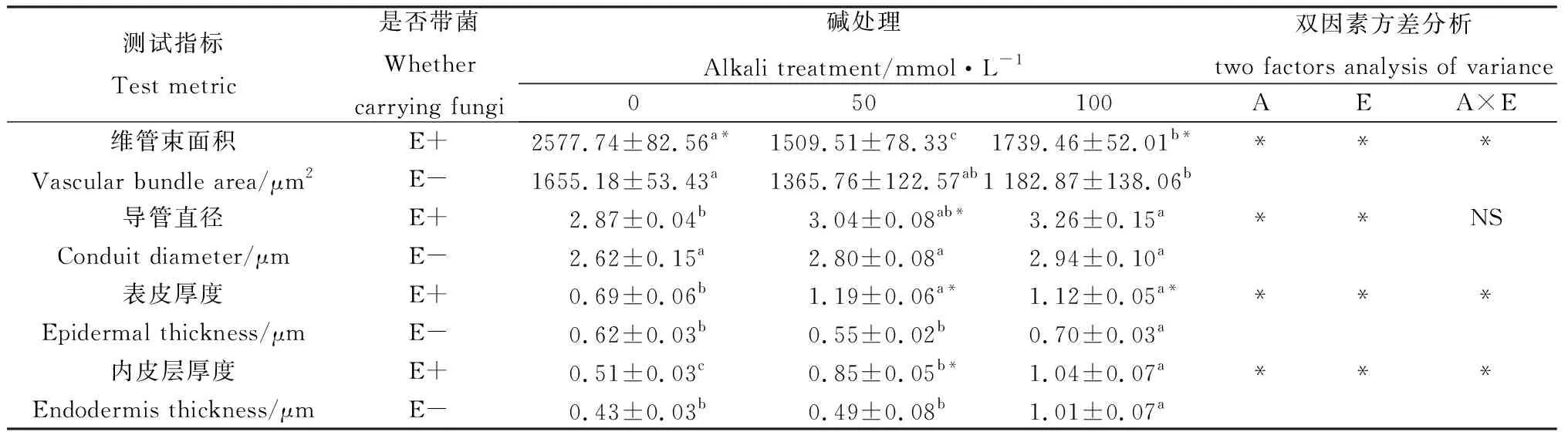
表1 碱胁迫下内生真菌对布顿大麦根结构的影响Table 1 Effects of endophytic fungi on root structure of Hordeum bogdanii under alkali stress
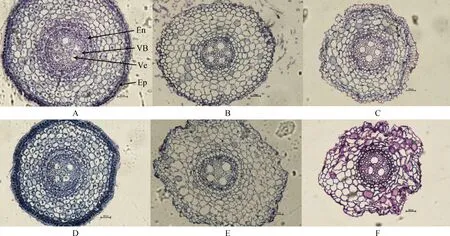
图1 不同碱浓度对E+和E-布顿大麦根的横切显微结构的影响Fig.1 Effects of different alkali concentrations on the transverse microstructure of E+ and E- Hordeum bogdanii roots注:A,B,C分别为E+ 0 mmol·L-1,E+ 50 mmol·L-1,E+ 100 mmol·L-1;D,E,F分别为E- 0 mmol·L-1,E-50 mmol·L-1,E- 100 mmol·L-1;En,内皮层;VB,维管束;Ve,导管;EP,表皮;标尺为200 μm,放大倍数为20×10Note:A,B and C are E+ 0 mmol·L-1,E+ 50 mmol·L-1 and E+100 mmol·L-1 respectively;D. E and F are E- 0 mmol·L-1,E- 50 mmol·L-1 and E- 100 mmol·L-1 respectively;En,endothelial layer;VB,vascular bundle;Ve,catheter;Ep,epidermis;Scale is 200 μm,the magnification is 20×10
2.2 碱胁迫对布顿大麦植株茎的显微结构影响
不同浓度碱胁迫对E+和E-布顿大麦茎的显微结构的影响见图2和表2。由表2可知,碱胁迫对布顿大麦E+和E-维管束面积无显著影响,并且E+与E-在维管束面积上无显著差异。碱胁迫缩小了E+植株的气腔直径,对E-无显著影响;在0 mmol·L-1和100 mmol·L-1碱胁迫发现E+的气腔直径显著长于E-(P<0.05)。随着碱浓度的增加茎的表皮厚度在逐渐增加。由表2双因素方差分析可知,碱胁迫和内生真菌显著影响表皮厚度,但两者之间不存在交互作用。
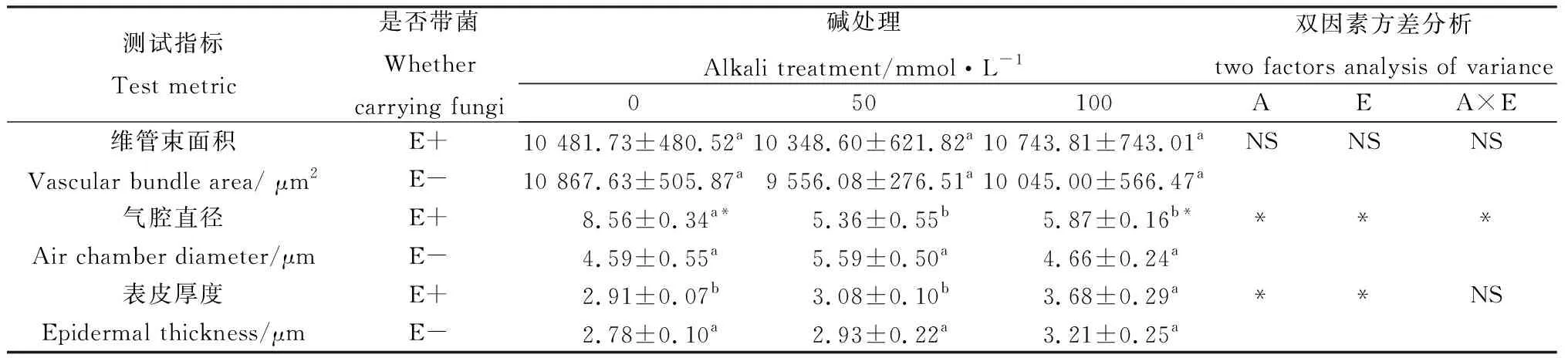
表2 碱胁迫下内生真菌对布顿大麦茎结构的影响Table 2 Effects of stem structure of Hordeum bogdanii by endophytic fungi under alkali stress
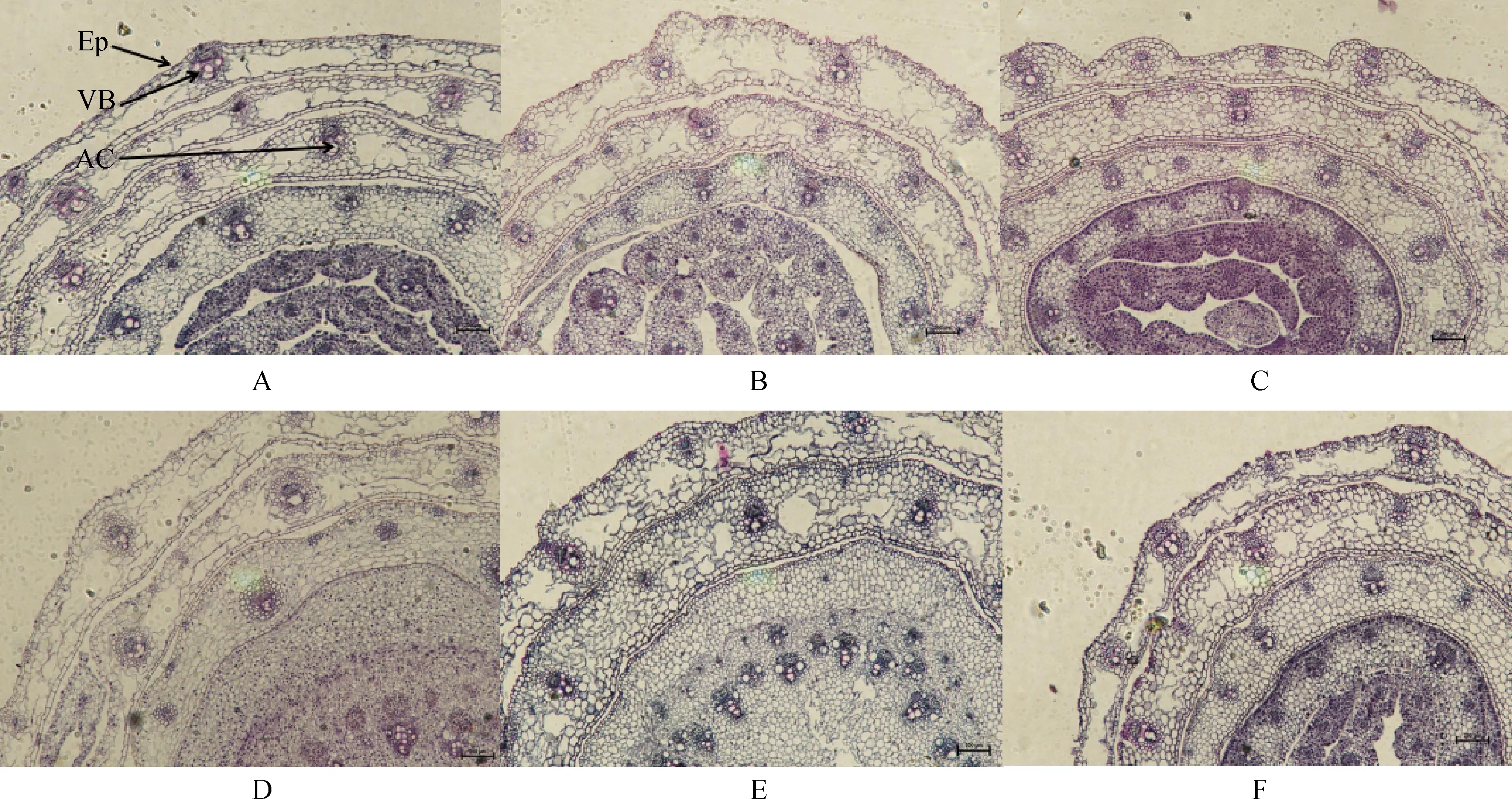
图2 不同碱浓度对E+和E-布顿大麦茎的横切显微结构的影响Fig.2 Effects of different alkali concentrations on the transverse microstructure of E+ and E- Hordeum bogdanii stems注:A,B,C分别为E+ 0 mmol·L-1,E+ 50 mmol·L-1,E+ 100 mmol·L-1;D,E,F分别为E- 0 mmol·L-1,E- 50 mmol·L-1,E- 100 mmol·L-1;En,内皮层;VB,维管束;AC,气腔;EP,表皮;标尺为200 μm,放大倍数为10×10Note:A,B and C are E+ 0 mmol·L-1,E+ 50 mmol·L-1 and E+ 100 mmol·L-1 respectively;D,E and F are E- 0 mmol·L-1,E- 50 mmol·L-1 and E- 100 mmol·L-1 respectively;En,endothelial layer;VB,vascular bundle;AC,air chamber;Ep,epidermis;Scale is 200 μm,the magnification is 10 × 10
2.3 碱胁迫对布顿大麦植株叶的显微结构影响
从图3叶片的显微结构图可知,布顿大麦的上下表皮细胞大小不规则,上下表皮都存在气孔,泡状细胞只存在于上表皮。随着碱浓度的升高,布顿大麦叶的维管束面积逐渐降低,泡状细胞的细胞面积增大,同一碱浓度处理下E+的维管束面积和泡状细胞面积高于E-,并且维管束面积达到显著水平(P<0.05)。碱胁迫缩小了E-的导管直径,但对E+的导管直径无显著影响,在100 mmol·L-1碱浓度下E+的导管直径显著大于E-(P<0.05)。在叶片气孔直径中发现50 mmol·L-1碱浓度处理下E+的显著大于E-(P<0.05)。通过双因素方差分析发现碱胁迫显著影响布顿大麦叶的维管束面积、泡状细胞面积、导管直径以及气孔直径;内生真菌显著影响叶片的维管束面积和气孔直径(P<0.05),但内生真菌和碱胁迫不存在交互作用(表3)。
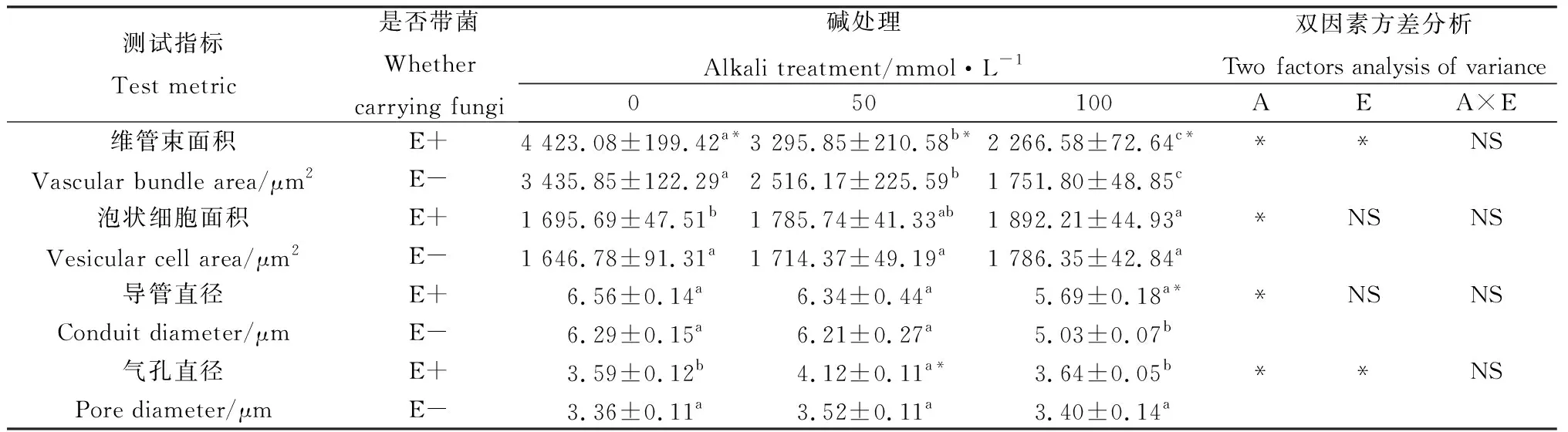
表3 碱胁迫下内生真菌对布顿大麦叶结构的影响Table 3 Effects of leaf structure of Hordeum bogdanii by endophytic fungi under alkali stress
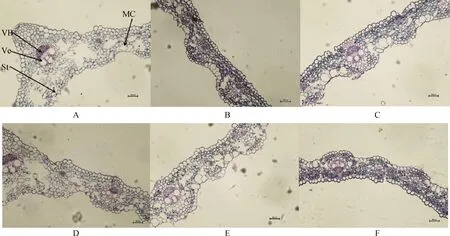
图3 不同碱浓度对E+和E-布顿大麦叶的横切显微结构的影响Fig.3 Effects of different alkali concentrations on the transverse microstructure of E+ and E- Hordeum bogdanii leaves注:A,B,C分别为E+ 0 mmol·L-1,E+ 50 mmol·L-1,E+ 100 mmol·L-1;D,E,F分别为E- 0 mmol·L-1,E-50 mmol·L-1,E- 100 mmol·L-1;VB,维管束,Ve,导管;St,气孔;MC,泡状细胞;标尺为200 μm,放大倍数为20×10Note:A,B and C are E+ 0 mmol·L-1,E+ 50 mmol·L-1 and E+ 100 mmol·L-1 respectively;D,E and F are E- 0 mmol·L-1,E- 50 mmol·L-1 and E- 100 mmol·L-1 respectively;VB,vascular bundle;Ve,catheter;St,air hole;MC,vesicular cells;Scale is 200 μm,the magnification is 20×10
3 讨论
植物的形态结构总是与环境相适应。逆境环境影响植物生长,并可引起植物形态结构的相应变化[19]。维管束是主要输水组织,碱胁迫导致根的维管束横截面面积减小,韧皮部和木质部的传导能力明显减弱[20]。赵海新等[21]的研究也证实了碱胁迫可促使植物根系维管束数目减少、木质化程度加强、导管数量增加等方式来提高根的运输能力。此外,陈泰祥[22]研究发现碱胁迫对野大麦的生长影响强于盐胁迫;内生真菌侵染减缓了宿主野大麦(Hordeumbrevisubulatum)在盐胁迫下根和茎的皮层厚度的降低趋势,增强了根的内皮层厚度的增高趋势;同时内生真菌也减缓了叶片厚度、维管束的降低趋势[23]。本研究与陈泰祥的研究结果相似,碱胁迫导致布顿大麦根维管束横截面积减小,木质化程度加深,导管直径变大,这些结构的变化可能是布顿大麦为了适应碱胁迫带来的损坏做出的响应。同时,植物在受到外界环境的刺激下,植物会增加表皮厚度和内皮层厚度来防止体内水分过分的蒸腾,保持水分,并起着机械支持作用[24]。内生真菌的存在对植物显微结构的改变有关,真菌侵染与叶片木质部、叶片韧皮部、茎维管束、茎木质部导管和根间木质部面积显著相关。在NaCl胁迫下,Epichlo⊇内生真菌侵染对野大麦的叶脉、茎表皮、根皮层和根内皮层厚度增加有关[25]。本研究发现,Epichlo⊇内生真菌提高布顿大麦根的维管束面积、增加导管直径、表皮厚度和内皮层厚度,说明碱胁迫条件下,Epichlo⊇内生真菌的侵染对宿主细胞起到一定的保护能力,提高了宿主植物对水分和无机盐的吸收能力,并减少水分的散失。
植物通过木质部将根部吸收的水分和矿物质往上运输到地上部各营养器官,地上部光合作用的产物则通过韧皮部向下运输至地下部,并将叶片产生的有机物质运送到根内或暂存茎内[26]。据报道,碱胁迫导致草木犀(Melilotusofficinalis)茎秆侧面的维管束大量减少和上、下表皮厚度增加[20]。本研究对E+和E-植株的茎在不同浓度碱胁迫下的显微结构进行了观察,发现茎中维管束与髓部薄壁细胞都发达。在碱胁迫下,茎通过表皮厚度、维管组织髓结构的改变来适应环境的变化。茎的表皮厚度随着碱浓度增大逐渐加厚,说明植物通过增加表皮厚度减少机体水分的散失。同时,本研究发现在0 mmol·L-1和100 mmol·L-1E+植株的气腔直径显著高于E-,因气腔主要功能是有利于器官中的细胞呼吸时的气体交换,所以我们推测Epichlo⊇内生真菌对宿主植物在碱胁迫时细胞呼吸有促进作用。
叶是植物进行光合作用和蒸腾作用的主要器官,因结构特征的变化最能体现植物对环境的适应[27],所以研究最多的器官是叶[28-29]。叶脉在水分和养分的运输中发挥着重要的生理作用[30]。在碱胁迫下草木犀小叶的上、下表皮细胞壁增厚,减少了叶片非气孔路径的水分流失以及在200 mmol·L-1碱浓度处理后,木质部导管直径减少了47 %,增加了蒸腾阻力[20]。ARAFA等[31]发现,较高水平的盐胁迫(3 000和6 000 ppm NaCl)降低了高粱木质部和韧皮部组织厚度、叶片厚度、维管束尺寸以及后生木质部导管直径。本研究通过观察碱胁迫下E+和E-植株叶片显微结构的形态变化,发现随着碱浓度的增加叶片维管束面积变小,导致水分、无机盐和有机物质的输送能力减弱,使得叶肉细胞得不到充足的养分,所以在显微结构上表现出叶肉细胞体积变小。泡状细胞与叶的卷曲和伸开的作用有关,在水分吸收和保持细胞水势中起一定的作用,本研究发现碱胁迫使泡状细胞下陷较深,面积变大,使得在缺水时快速将叶片卷曲,减少水分的散失,这与盐胁迫下小花碱茅(P.tenuiflora)叶片泡状细胞的研究结果一致[32],但E+植物较E-植株泡状细胞下陷更深,面积变大,表明碱胁迫下Epichloё内生真菌能有效的防止布顿大麦叶肉细胞内水分的丢失。气孔直径的变化可以看出E+的先升后降,但碱处理后E+的气孔直径始终大于0 mmol·L-1时的气孔直径;E-无显著影响,说明Epichloё内生真菌具有促进布顿大麦气体交换能力。
4 结论
在碱胁迫下,内生真菌的侵染影响了布顿大麦的显微结构。碱胁迫下Epichloё内生真菌的侵染提高了根的维管束面积、增加了导管直径、表皮厚度和内皮层厚度,提高了植物对水分和无机盐的吸收能力,增加对细胞的保护和减缓水分的散失;Epichloё内生真菌增加了茎中气腔直径、表皮厚度和促进维管组织中髓的发育;同时,叶的泡状细胞面积和气孔直径变大,使得叶片的失水能力减弱;从而有效的防止布顿大麦叶肉细胞内水分的丢失,促进气体交换能力。本研究从形态结构方面证明了碱胁迫下Epichloё内生真菌提高了宿主布顿大麦的耐受性。
