SARS-CoV-2 Delta变异株:从结构变异到强传染力
2023-02-21魏逸鸣张俊文
魏逸鸣, 张俊文
(重庆医科大学附属第一医院消化内科, 重庆 400042)
冠状病毒为一种正链RNA病毒(+ssRNA),因其特征为包膜上存在众多的刺突糖蛋白、在电子显微镜下酷似王冠状的外观而得名。严重急性呼吸系统综合征冠状病毒2(severe acute respiratory syndrome coronavirus 2, SARS-CoV-2),为目前已确定的7种能够感染人类的人类冠状病毒之一。自2019年12月开始暴发的2019冠状病毒病(coronavirus disease 2019 , CoVID-19)与SARS-CoV-2野生型毒株密切相关,至今对人类社会造成了相当大的社会经济负担,已然成为全球范围内最受关注的公共卫生事件[1]。SARS-CoV-2如同所有RNA病毒一样易于遗传进化,导致了多种与祖先菌株相比具有不同特征的变体出现,这些特征主要表现为传播力和(或)毒力的增强、通过自然感染和(或)疫苗接种获得抗体的中和作用减弱、逃避检测能力、降低治疗和(或)疫苗接种的有效性等。为此,世界卫生组织建立了一个分类系统,将SARS-CoV-2谱系中具有特定遗传标记的变体定义为感兴趣的变体(variant of Interest, VOI);而更加受到高度重视与广泛关注的新变体则独立为受关注的变体(variants of Concern, VOC)。截至2021年12月,已经有4个主要谱系被世界卫生组织确定为VOC,包括Alpha(B.1.1.7毒株)、Beta(B.1.351毒株)、Gamma(P.1毒株)和Delta(B.1.617.2毒株)。Delta变异株于2020年10月在印度首次被检测到,最初被认为是VOI,但其极快的传播速度促使世界卫生组织在2021年5月将其归类为VOC,截至2021年11月16日,Delta变异株已经传播到至少110个国家,在大多数国家Delta变异株已胜过包括其他VOC在内的所有变体[2,3]。本文就SARS-CoV-2 Delta变异株的病毒结构、动力学及临床特征研究进展作简要综述。
1 Delta变异株的结构及功能改变
1.1 SARS-CoV-2蛋白质分子结构及Delta变异株突变位点
了解病毒结构对于解决Delta变异株的发病机制极其重要。SARS-CoV-2和其他冠状病毒一样,其主要组成部分为4种结构蛋白质:刺突蛋白(S蛋白)、包膜糖蛋白(E蛋白)、核衣壳(N蛋白)和膜蛋白(M蛋白)。除此之外,还有16种非结构蛋白质和5~8种辅助蛋白质。其中,S、E、N、M基因分别编码结构蛋白质,而非结构蛋白质则由14个功能性开放阅读框(ORF)编码[4]。刺突蛋白在病毒的致病和传播中发挥至关重要的作用。大量糖基化的刺突蛋白,位于病毒粒子的外表面,通过与宿主细胞受体结合介导病毒进入,这是病毒感染的关键因素[5]。血管紧张素转化酶 2(angiotensin converting enzyme 2, ACE2)已被证实为SARS-CoV-2的受体,而刺突蛋白的另一个已知受体——二肽基肽酶 4(dipeptidyl peptidase 4, DPP4),则并不能介导SARS-CoV-2进入靶细胞[6]。刺突蛋白在天然状态下作为无活性前体存在,在感染靶细胞的过程中,靶细胞膜上的蛋白酶(例如跨膜丝氨酸蛋白酶transmembrane protease serines2, TMPRSS2和Furin蛋白酶)将刺突蛋白切割成氨基端(N-端)S1亚基和羧基端(C-端)S2 亚基。其中,S1亚基中的受体结合域(receptor binding domain, RBD)负责与ACE2受体结合,而S2亚基的主要部分HR结构域,包括HR1和HR2,主要负责病毒的融合与进入[7-9]。在病毒的融合过程中,HR1与HR2相互作用形成6-HB,这一过程实现了刺突蛋白的缩短,以此拉进其与靶细胞的距离并实现融合(Fig.1 SARS-CoV-2结构)。膜蛋白是含量最丰富的蛋白质,为病毒组装创造环境,并决定了病毒包膜的形状。包膜糖蛋白是最小的主要结构蛋白质,参与了病毒组装,在病毒出芽过程中扮演了必不可少的角色。核衣壳蛋白是形成病毒核衣壳唯一的蛋白质,其主要作用是与冠状病毒RNA基因组结合并参与病毒基因组相关过程[10]。

Fig.1 The Structure of SARS-CoV-2 (A) Relative position of SARS-CoV-2 structural protein genes. (B) Structure and domains of spike protein① and proteases on the target cell membrane—Furin (S1/S2) and TMPRSS2 (S2′) cleavage sites, in which NTD means N-terminal domain and RBM means receptor-binding motif. (C) After the spike protein binds to the ACE2 receptor, the spike protein is cleaved into S1 and S2 subunits, in which the HR1 and HR2 subunits in S2 form a 6-helix bundle (6-HB) to promote the interaction of SARS-CoV-2 with Target cell membrane fusion
SARS-CoV-2结构的复制从附着和进入靶细胞开始。刺突蛋白与靶细胞ACE2受体结合后,蛋白酶裂解刺突蛋白促使病毒进入,随之病毒RNA被释放并翻译多种蛋白质组装复制酶-转录酶复合物,RNA得以复制。待结构蛋白质完整组装后,病毒粒子以囊泡形式运输到细胞表面,并通过胞吐作用释放。这便是SARS-CoV-2在靶细胞中的整个生命周期。
Nextstrain基因树以浅显易懂的预估变异株出现时间年份加字母排序命名各变异株,同时加注世界卫生组织官方命名。Delta变异株(21A)在20A基础上进一步突变,与另一VOC Kappa变异株同源(Fig.2)。在基因树上可追溯的Delta变异株与SARS-CoV-2野生株相比,总计有23个突变。其中,18个为非同义突变(见Table 1 SARS-CoV-2 Delta亚型的非同义突变(B.1.617.2)),这并不代表Delta变异株中的每个序列都拥有所有的突变[11]。Delta变异株有9个突变位点与刺突蛋白相关,也被认为是其成为最具传播性变异株的关键突变。在刺突蛋白上,受体结合域中的突变被普遍认为与病毒传染性、人与人之间的传播、免疫逃逸紧密挂钩,因此显得尤为敏感。在Delta变异株中,包括L452R和T478K。
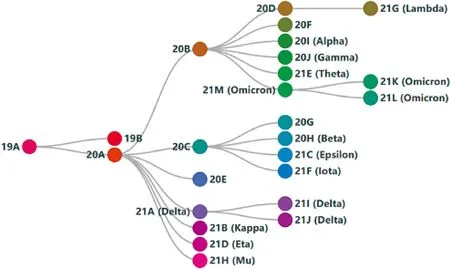
Fig.2 Phylogenetic relationships of Nextstrain clades of SARS-CoV-2 The origin and developmental relationship of the variants/clades of SARS-CoV-2. Variant names use the Nextstrain SARS-CoV-2 clade naming strategy, which means "year-letter" accorded by the official WHO designation [11]

Table 1 Non-synonymous mutation of SARS-CoV-2 Delta variant (B.1.617.2)
1.2 Delta变异株突变位点对中和抗体和血清的影响
L452R突变用精氨酸替代了452位的亮氨酸,在全球范围的刺突蛋白受体结合域流行突变频率中高居第二[12]。亲水性452R突变消除了L452残基的疏水作用,使得突变复合物具有较低的最小能量,这可能有助于与水分子的相互作用以及复合物的整体稳定性,进而允许刺突蛋白以更高的亲和力附着在ACE2受体上[13]。这样的特点使得L452R突变面对多种抗体均表现出了中和效力降低。例如,LY-CoV555/bamlanivimab单克隆抗体,原本L452残基与其结合界面为疏水区,基因突变的发生导致了抗体敏感性的降低[14]。针对LY-CoV555的所有逃避抗体结合的受体结合域突变组合中,L452R表现出了第2高频率的抗体逃逸,也从反面印证了这一突变特征[15]。目前,bamlanivimab EUA的情况说明书中已明确提示,L452R突变使bamlanivimab中和作用降低了1 000倍以上,这意味着bamlanivimab的抗体应用应避免单药治疗,尤其对于Delta变异株广泛传播的地区。
T478K突变,用正氨基酸(赖氨酸)取代了不带电荷的氨基酸(苏氨酸),这可能显著改变了蛋白质的静电表面,使其较突变前拥有了更高的净电荷、偶极矩,产生较低的溶剂化和表面自由能,从而降低蛋白质的稳定性,使刺突蛋白得以更容易与ACE2受体相结合[16]。结果显示,L452R+T478K的基因突变组合的确增加了受体结合域与ACE2受体之间的结合亲和力[17]。通过与Kappa变异株比较得出,T478K突变极有可能影响imdevimab单克隆抗体的中和效力,其机制尚不明确[18]。
除了直接影响受体结合域,基因突变还能通过影响刺突蛋白的切割来加快病毒传播进程,例如P681R突变。P681R突变用精氨酸取代了681位的脯氨酸,这可能增加了靶细胞膜上蛋白酶额外接触,促进S1~S2亚基切割,允许病毒更好地融合和整合到靶细胞中[13]。
D614G突变用甘氨酸取代了614位的天冬氨酸,这增加了刺突蛋白RBD的空间灵活性,使得刺突蛋白有更高的可能性与ACE2结合与融合[19]。在体外研究中,D614G突变已被证明提高了病毒传染性[20,21]。
与先前提及的单克隆抗体的作用机制不同,BNT162b2是一种通过脂质纳米配方将SARS-CoV-2完整的刺突蛋白固定在病毒融合前构象的mRNA疫苗[22]。已有研究证实,接种BNT162b2后的人类血清对Delta变异株相比于SARS-CoV-2野生株依然有超过80%的中和效力[23,24],说明Delta变异株的强感染力并不是通过刺突蛋白的显著改构而产生抗原性的变化。与BNT162b2类似,ZF2001是一种以刺突蛋白RBD为抗原的蛋白质亚单位疫苗,其对Delta变异株同样保有中和活性[25]。事实上,相比于B.1.1.7毒株的活病毒颗粒,Delta变异株中检测到更多的切割刺突[26]。这进一步证实,其刺突蛋白基因上的关键突变所引起的蛋白酶切割增加效应增强了Delta变异株的感染力。
1.3 Delta变异株的蛋白质表征
在体外研究中证实,当刺突蛋白在细胞表面以饱和水平表达时,Delta变异株可以比其他变体更快地融合细胞膜和感染靶细胞,尤其会更有效地与产生低水平ACE2受体的靶细胞融合,这使得Delta变异株能在相对短暂的暴露后传播,并迅速感染更多的宿主细胞,从而导致潜伏期缩短和感染期间病毒载量变大。但是,研究表明,完整的全长Delta变异株的刺突蛋白受氟林蛋白酶(靶细胞膜上蛋白酶)切割的效率并无显著增加,且与ACE2受体的亲和力亦无明显升高[27],这与单独研究Delta变异株突变位点相关变化截然相反。
2 病毒动力学
Delta变异株有人群快速传播的特点,这个特点以更加具体的方式反映在单个感染者身上。与SARS-CoV-2野生株相比,Delta变异株感染患者的峰值病毒载量明显更高,且病毒的脱落时间更长[28,29]。在相对小而密闭的空间中,Delta变异株表现出了更强的流通性。在俄克拉荷马州,一项针对体操设施相关的Delta变异株暴发案例提示,室内运动场所具有高风险性,并且家庭中的二次传播率有显著升高[30]。与Alpha变异株相比,Delta变异株相关的家庭传播几率增加了70%[31]。
2021年5月至6月,在中国广东省集中暴发了COVID-19 Delta变异株感染,尽管在广东省的积极干涉措施下最终得以控制,其快速传播力仍导致共167例病例感染。相较于2019年12月中国湖北省首次暴发的COVID-19 野生株感染,广东省COVID-19 Delta变异株感染平均潜伏期由5.2 d缩短至4.4 d,平均感染间隔时间更是由5.7 d缩短至2.9 d[32]。这一阶段疫情中,Delta变异株感染患者病毒载量在血清学检测阳性初期有显著增高[33]。
3 临床特征
3.1 SARS-CoV-2野生株感染COVID-19患者的临床特征
据统计,有17.9%至33.3%的COVID-19 SARS-CoV-2 野生株感染患者为无症状感染者,而在有症状的患者中,绝大多数的临床表现为发烧、咳嗽和气短,其余较少出现的症状包括喉咙痛、嗅觉障碍、味觉障碍、厌食、恶心、不适、肌痛和腹泻等[34,35]。在美国一项针对373 883例确诊的有症状的COVID-19病例研究中,70%的患者有发烧、咳嗽和呼吸急促的表现。除此外,第2、第3高频率症状,肌痛与头痛,分别仅占36%和34%[36]。在中国,COVID-19 SARS-CoV-2 野生株感染患者的临床特征与世界其他国家差异不大。对中国8 697名COVID-19患者临床病理特征的研究表明,发烧(78.4%)和咳嗽(58.3%)仍为最普遍的症状。这项研究中的无症状患者仅占5.4%[37]。
3.2 SARS-CoV-2 Delta变异株感染COVID-19患者的临床特征
目前,已知的SARS-CoV-2 Delta变异株感染常见症状包括发烧、咳嗽、呼吸急促、呕吐、腹泻、喉咙痛和头痛,诸如肌痛、味觉丧失、嗅觉丧失、疲劳和流涕等症状亦有报道。其中,发烧与咳嗽依然占据主要比例,整体临床表现不具备特异性。一项针对中国广州的159例确诊Delta变异株感染患者的研究中,中度症状的感染患者以61.6%的比例占大多数;轻症患者占25.8%;重症及危急症患者占12.5%。其中,呼吸道症状与野生株感染患者相差不大,但腹泻、呕吐等消化道症状明显少于SARS-CoV-2野生株感染患者,同时老年患者更多[28]。在苏格兰,SARS-CoV-2 Delta变异株感染患者有年轻化的特点[38]。
4 问题与展望
SARS-CoV-2 Delta变异株作为目前印度新型冠状病毒肺炎的主要致病株,已经表现出了惊人的传播力,并对欧美国家造成了大面积的感染。中国作为目前全世界抗疫防疫最为严格与成功的国家,亦需要对其进行全面深入的了解,以做到提前预防、尽早发现、妥善治疗。事实上,中国针对新型冠状病毒肺炎的防控措施已被证明是有效的,这些包括戴口罩、保持社交距离、限制社交活动和社区区域封锁等措施也有助于遏制目前SARS-CoV-2 Delta变异株的爆发。
尽管快速传播已是SARS-CoV-2 Delta变异株受世界公认的特点,但其发生机制仍有争议。追溯其基因组的突变位点,位于RBD的L452R和T478K突变被认为是影响其传播力的根本因素,因为很有可能通过改变RBM对ACE2受体的亲和力来增加病毒感染效率。P681R可能增加靶细胞膜蛋白对S1~S2亚基的切割效率。D614G突变则通过提升刺突蛋白RBD空间灵活性来增加与ACE2受体的结合可能。总而言之,Delta变异株的刺突蛋白基因突变都指向Delta变异株通过增加与靶细胞膜的亲和力而非改变抗原性来实现感染力的增强。然而,根据Zhang等[27]对全长刺突蛋白的研究,SARS-CoV-2 Delta变异株对ACE2低表达水平细胞有更明显的传播增强效果。反之,对ACE2受体亲和力以及靶细胞膜蛋白的切割效率均无明显变化。目前仍需要更多的研究证据来解释SARS-CoV-2 Delta变异株的快速传播特点。
SARS-CoV-2 Delta变异株被列为受关注变体的另一个重要原因是其对中和抗体和血清的免疫逃避。基于L452R突变,单克隆抗体bamlanivimab中和效力降低了1 000倍以上。单克隆抗体imdevimab对于Delta变异株的中和效力同样有所降低。然而,拥有完整SARS-CoV-2野生型刺突蛋白结构的mRNA疫苗BNT162b2,以及蛋白质亚单位疫苗ZF2001仍对Delta变异株保有可观的中和效力,这对未来的疫苗接种提供了方向。目前,中国广泛接种的主流疫苗类型为灭活疫苗。研究表明,灭活SARS-CoV-2疫苗对Delta变异株感染估计疫苗有效性较野生株降低了20%,不过根据世界卫生组织设定的50% 的疫苗有效性最低阈值,接种灭活疫苗对Delta变异株依然有效[39]。
SARS-CoV-2 Delta变异株感染所致症状仍以咳嗽、发热为主要表现。这表明,其易感器官及致病作用并未发生本质改变。但是,仍有值得注意的临床特征。SARS-CoV-2 Delta变异株感染表现出消化道症状明显减少的特征。这提示,COVID-19的胃肠道表现可能并不以ACE2介导的肠粘膜病毒细胞毒性为主要的发病机制。多个研究表明,相比于SARS-CoV-2野生株,Delta变异株更容易感染青少年及老年人。其中,老年人的易感性与其免疫衰老有关,尤其是炎症细胞因子的产生增加[40]。
在全球新型冠状病毒危机的大环境下,Delta变异株以其广泛的传播能力成为了全世界都无法忽视的SARS-CoV-2变体。但目前看来,它可能并不是病毒遗传变异的终点。2021年11月9日,在南非首次被检测到的Omicron变体(B.1.1.529菌株)已以毫不逊色于Delta变异株的传播速度散布于多个国家和地区。面对SARS-CoV-2 Delta病毒变异株,未来仍需更多研究成果证实其作用机制以指导治疗与预防。
