CRISPR/Cas9系统在临床医学研究的应用与改进
2023-02-21杨育宾黄卫人
杨育宾, 黄卫人, 陈 巍
(1)汕头大学医学院药学系,广东, 汕头 515041;2)深圳市第二人民医院医学合成生物学临床应用关键技术国家地方联合工程实验室,广东, 深圳 518036)
锌指核酸酶(zinc finger nuclease,ZFN)和转录激活因子样效应核酸酶(transcription activator-like effect nuclease,TALEN)是两类可编程的作用于基因组编辑的核酸酶。ZFN[1]和TALEN[2]是由序列非特异性DNA核酸酶结构域融合序列特异性DNA识别结构域产生的模块化蛋白质。特异性的DNA识别结构域的构建是极费时间与资金的[3]。近年来,新型的CRISPR/Cas9系统由于组分简单、靶向性高、对靶基因的编辑效率高等特点,逐渐成为临床医学研究中应用最为广泛的基因编辑工具。通过对sgRNA的简单编码,CRISPR/Cas9系统可实现对不同靶标的编辑,极大程度拓展了应用范围。但单一的CRISPR系统只可实现对靶基因的敲除,难以满足实际研究中对基因双向调控的需求。对CRISPR元件的功能进行改造,拓展使用范围和减少应用限制已成为日益关注的重点。本文对CRISPR/Cas9基因编辑技术的应用与其系统元件改造后在基因编辑、活体细胞成像、降低应用障碍等方面的研究进展和前景做简要阐述。
1 CRISPR系统基因编辑的原理
细菌在漫长的自然演化过程中为适应噬菌体类型的多态性,广泛产生了多种噬菌体抵抗机制。其中,CRISPR/Cas系统(clustered regularly interspaced short palindromic repeats/CRISPR associated proteins,CRISPR/Cas)是细菌具有的获得性免疫防御系统,可帮助细菌抵抗外源核酸以及噬菌体的入侵[4]。CRISPR的结构中包含了短的部分回文结构的重复DNA序列。重复序列并非连续而是存在一定的间隔,最终形成了交替重复元素的直接重复序列(21~48 bp)和可变序列(26~72 bp,即间隔体)的基因座结构[5]。CRISPR/Cas系统根据效应器模块设计的原则可分为两类,第一类为多亚基效应复合物,第二类为单蛋白质效应模块[6]。一般而言,不同的CRISPR/Cas系统的作用机制基本类似。获得性免疫防御系统的激活一般分3步进行:(1)适应。在该过程中,入侵的外源核酸或噬菌体DNA的原间隔邻近序列被识别,识别后在CRISPR阵列前端(富含AT和启动子的前导序列以及重复序列间)将新的间隔序列进行位点特异性的整合;(2)crRNA的成熟。当相同的外源核酸或噬菌体再度入侵时,对应的CRISPR阵列被激活,产生前体的crRNA(pre-crRNA)被加工为成熟的crRNA;(3)干扰。crRNA与对应的Cas蛋白形成Cas-crRNA复合物识别并降解外源核酸或噬菌体。本文主要关注于CRISPR/Cas9系统的相应研究。
在CRISPR/Cas9系统中,成熟的crRNA与反式激活的crRNA(tracrRNA)结合形成sgRNA。sgRNA与Cas9蛋白形成复合物。Cas9蛋白在sgRNA的导向作用下通过碱基互补配对结合到靶标处。随后,Cas9蛋白识别靶基因的PAM序列(protospacer adjacent motif, PAM),并在靶基因的PAM上游的第3个碱基开始切割靶标DNA,产生双链断裂位点(double strand breaks, DSBs)。DSBs的产生诱发机体内的DNA修复机制。这种修复机制可分为同源重组修复(homologous recombination,HR)和非同源末端连接(non-homologous end joining,NHEJ)。其中,同源重组修复可精确地插入目的片段,但需要提供外源的DNA修复模板,且该修复机制的发生频率低(Fig.1)。生物体内更倾向于发生非同源末端连接修复,该修复过程中碱基的插入或缺失是随机的,改变了靶基因原先的核苷酸序列[7]。基于该特性,CRISPR/Cas系统可用于基因编辑,建立多种细胞系及动物模型[8],或生产更为广泛的基因修改动物等[9]。
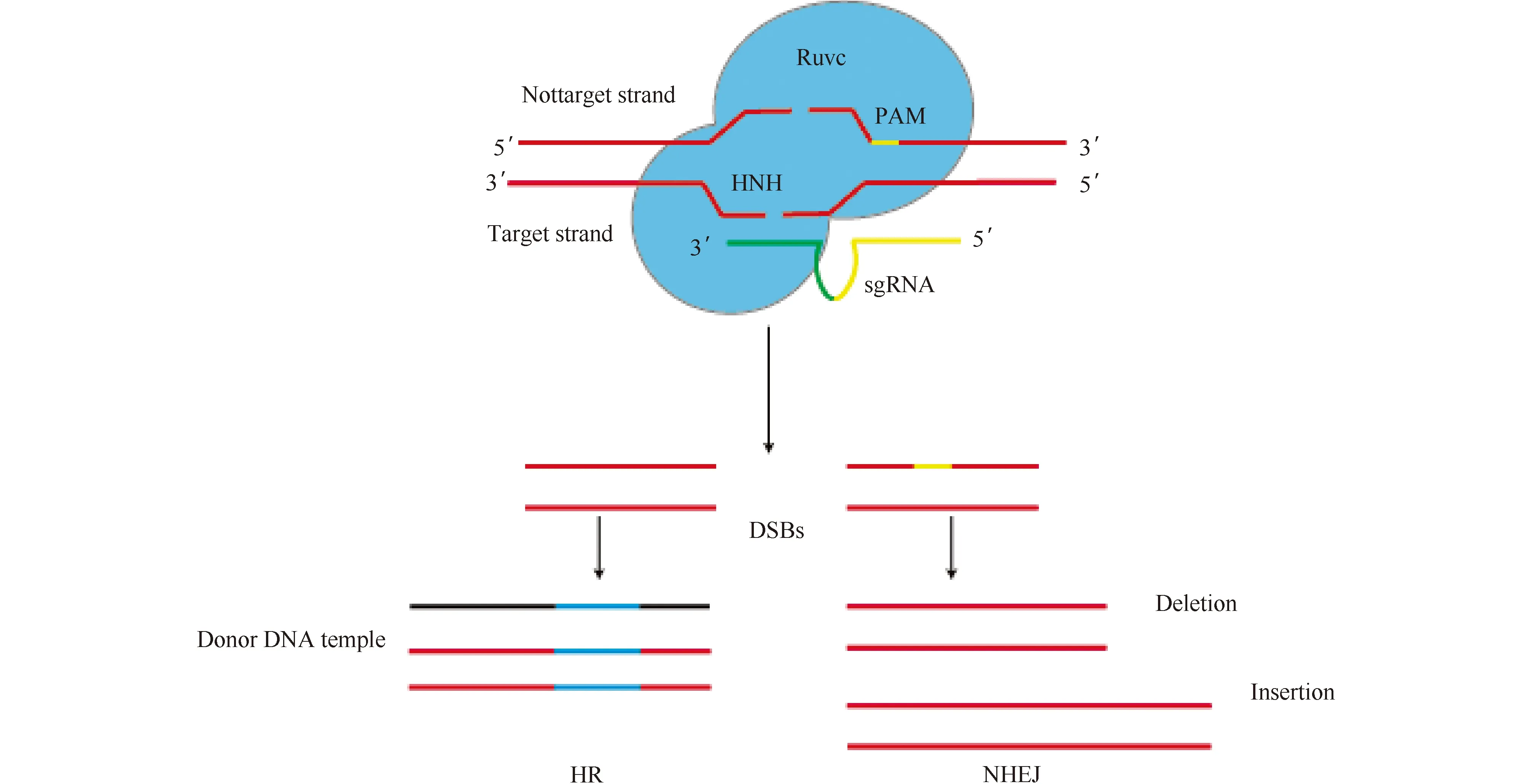
Fig.1 Schematic diagram of the cas9 system’s action principle and repair principle
2 活细胞染色体成像技术的应用与改进
生物体应对外界刺激所进行的基因表达调控会引发染色质动态空间结构的变化,这种变化反映基因组的时空调节,在控制生物体的转录、细胞分化和发育中发挥直接作用。构建可时实检测染色质的空间结构变化的工具,能极大促进对基因表达的物理调节和相互作用的认知[10],更好的了解疾病发生发展的分子机制。
早期的研究表明,通过设计特异性DNA荧光探针,实现了细胞内多个基因组位点的可视化检测,获得了一些关于基因组结构的最早见解[11]。但DNA荧光原位杂交存在着许多缺点[12]。原位杂交技术需要对细胞进行固定,再经过加热和甲酰胺变性来实现荧光探针与靶点的结合,因此该方法不适用于活体细胞成像。且变性条件会引发基因组结构损伤,导致研究结果的可靠性存疑。基于CRISPR系统的靶向性和能用sgRNA的简单重编程进行改变的特性,开始对CRISPR组分进行重编程,在不破坏基因组结构的基础上,通过输入荧光信号获得靶点可视化的输出结果,实现了对活细胞时实成像。2013年,Chen等[13]开创性地将dCas9用于基因组成像,通过将dCas9与核定位序列和eGFP标签的融合,实现在适量的sgRNA表达后动态显示活细胞中的端粒。这种基于对CRISPR组分进行重编程的思维,极大扩展了CRISPR系统的应用。但只是将dCas9与荧光蛋白质融合进行活体细胞成像,在实际应用中依旧存在着较大的障碍。首先,CRISPR系统本身具有脱靶效应,确保荧光信号代表的位点是靶点而非脱靶位点是应用该工具的前提。其次,核质中游离的dCas9荧光融合蛋白质,一些细胞中的自发荧光及核仁中与核糖体RNA非特异结合所富集的dCas9荧光融合蛋白等背景信号都会影响信噪比,干扰结果的判读。最后,靶向非重复的序列区域时,为了获得足够的检测信号,通常需要大量sgRNA进行共转染,增大成像难度。
随着研究的不断进展,上述难题也逐步得到解决。检测是否产生脱靶效应的最有效的方法是标记多个成像组件,然后检查多色共定位,从而排除脱靶误读[14, 15]。Cas9-sgRNA支架的四环和3′-末端区域已被证明非常易于修饰,可应用为添加荧光蛋白质进行多色标记的手柄[16]。降低背景信号的干扰可通过减少游离的Cas9荧光融合蛋白质和增加sgRNA的表达来实现。多西环素诱导启动子门控的dCas9 mRNA系统能人为控制Cas9荧光融合蛋白质的表达水平。而对于sgRNA,茎环基序的稳定以及多个导向表达结构的修饰会增强sgRNA表达[17]。实际上,游离的Cas9荧光融合蛋白质是导致背景信号过强的主要原因,通过荧光标记sgRNA可除去Cas9荧光融合蛋白质的干扰。基于该理念,Wang等[18]采用cy3标记了sgRNA。该研究发现,在非靶标处95%的sgRNA被体内的核酸酶降解,而靶标处形成的cas9-gRNA-DNA三元复合物保护了sgRNA,使得靶标处信号稳定且持久,信噪比高于Cas9-egfp成像系统4倍以上。非重复基因组区域的活细胞成像,通常需要转染多达26~36个不同的sgRNAs。为了降低非重复组区域的成像难度,Qin等[19]构建了含有多达16个MS2结合模块的工程化sgRNA支架,通过增加单个sgRNA的荧光标记量来放大荧光信号。该工程化的成像系统能够使用单个sgRNA对低重复序列的区域进行多色标记,而在非重复序列的区域仅需4个sgRNA即可进行成像。
3 CRSPR系统基因编辑功能的应用与改进
CRISPR基因编辑功能为临床上肿瘤分子机制的研究、癌症免疫疗法的发展以及遗传疾病等难治性疾病的治疗开创了新的方案。对比于DNA识别结构域构建困难的ZFN和TALEN核酸酶,CRISPR系统可通过简单的sgRNA重构来靶向不同的目的基因,能更加方便快捷地进行靶基因的编辑。但CRISPR系统在编辑目的基因时产生的DSBs会引发体内易错的非同源末端连接修复,从而可能引入致病突变,在应用CRISPR基因编辑功能同时,若能通过改进CRISPR系统来降低其DSBs产生,将能使之更加安全地应用于临床治疗中。
3.1 CRISPR基因编辑功能的应用
CRISPR系统是临床研究中应用最为广泛的基因编辑工具之一,CRISPR强大的基因编辑功能在筛选关键基因、癌症免疫疗法以及遗传疾病的治疗等方向都展现出巨大的价值。
CRISPR/Cas9筛选可有效地解码生物学系统的功能,是发现癌症治疗新靶点和识别基因与表型相关性的强大基因组学工具。通过构建sgRNA慢病毒文库,将之以较低的感染度感染稳定表达Cas9蛋白的细胞,而后使之暴露于设计的干扰下,即可高通量、快速地检测细胞群体中特定基因敲除的表型效应[20]。CRISPR筛选在临床研究中一般有2种用途:(1)通过测序不同转染时间段细胞中sgRNA的丰度,即可识别基因型特异性的脆弱性,用以筛选获得细胞生长必须的关键基因(Fig.2A)。这些“必需”基因可能成为潜在的药物靶点。因为,它们的功能缺失导致生存能力降低[21]。(2)CRISPR筛选还会得出在不同的转录背景下,相同基因的扰动可能带来的差异化结果,这种现象被称为合成致死性(Fig.2B)。简而言之,一个细胞能够补偿基因A或B的功能改变,但如果基因A和B同时改变,则无法存活,这种基因间相互作用的关系称为合成致死性。转化为癌症生物学,这表明特定肿瘤抑制因子的丢失或癌基因的激活,会导致以往生长非必需的基因表达成为必需。一项临床研究证明了这一概念。该研究表明,BRCA基因突变的乳腺癌对多聚ADP核糖聚合酶(poly(ADP-ribose) polymerase ,PARP)抑制剂具有更高的敏感性[22]。与此类似,Shen等[23]使用双敲除筛选方法对73个癌症基因进行配对筛选,发现了许多已知(例如BRCA-PARP)和未知的合成致死相互作用,其中一些相互作用可以在体外用对应的功能抑制药物进行组合模拟。合成致死关系的探究为精准基因治疗提供了理论基础,也为精准药物治疗提供了参考建议。

Fig.2 Diagram of CRISPR screening (A) CRISPR screening identifies key genes;(B) Diagram of synthetic lethality
癌症免疫疗法是临床上最为基础且常用的癌症治疗方法之一。近年来,随着CRISPR基因编辑系统的发展,癌症免疫疗法更侧重于T细胞受体编辑疗法(genetically modified T-cell receptor therapy,TCR)和嵌合抗原受体T细胞疗法(chimeric antigen receptor T cell therapy,CAR-T)。在机体内,癌症细胞通过过度表达免疫检查点分子来逃逸免疫细胞的杀伤作用。已有研究表明,通过对T细胞表面的免疫检查点分子受体进行编辑,能发挥良好的肿瘤杀伤作用甚至逆转肿瘤细胞的免疫耐受性[24]。但TCR疗法在编辑体内分子受体时,需克服内源性已有的TCR分子与外源编辑的TCR分子间配对形成异二聚体、内源性TCR分子竞争结合CD3分子干扰受编辑的TCR表达等难题。解决上述问题最有效的方法是引入TCR分子的同时敲除内源性的TCR,即在引入TCR分子的同时,转染靶向内源性TCR-β链的CRISPR慢病毒[25]。而在CAR-T疗法中,CRISPR系统可以将特异性的CAR靶向T细胞受体,使T细胞表达特异性的CAR。编辑后的CAR-T细胞在体外进行增殖培养,最后将之回注于患者体内,增强患者机体对肿瘤细胞的免疫应答[26, 27]。而在遗传疾病的治疗中,CRISPR系统较常用于异常分子的敲除与突变区域的切割。例如,在β-地中海贫血和镰刀状细胞贫血症2种遗传病中,编码血红蛋白的基因单碱基突变使得β-珠蛋白的表达缺失,进而导致贫血。之前的研究显示,γ-珠蛋白与β-珠蛋白的功能相似,而BCL11A转录因子可以抑制γ-珠蛋白的表达。这表明,BCL11A分子的敲除或能逆转珠蛋白表达缺失诱发的贫血。Frangoul等[28]研究验证了该理论的可行性。其研究结果表明,在BCL11A分子敲除后,患者不再需要通过依赖传统的输血来治疗β-地中海贫血和镰刀状细胞贫血症。
3.2 基因编辑功能在临床应用研究中的改造
由CRISPR系统的工作原理可知,Cas9的基因编辑系统在插入和缺失目的基因时通常会产生DSBs。DSBs引发了生物体内的DNA修复机制,且该修复机制更倾向于非同源末端连接修复,导致碱基的插入与缺失是随机的。不可忽视的是,绝大多数致病性等位基因来自特定的插入、删除或碱基替换。由此可知,在使用Cas9系统时可能会引入致病突变[29]。能否对CRISPR元件进行重编程,使之更精确地操纵基因是近年来临床医学研究领域的研究热点之一。
dCas9蛋白的出现为CRISPR系统的改造打开了突破口。通过对Cas9结构域的改造,使之丧失内切酶活性,保留sgRNA导向的靶向性。利用该特性,将修饰后的dCas9蛋白(dead Cas9, dCas9)和腺嘌呤脱氨酶进行融合。通过dCas9的靶向性,成功实现了在靶点处不造成DNA双链断裂的情况下,将A-T碱基对置换为G-C碱基对,该碱基编辑器被称为腺嘌呤碱基编辑器(adenine base editors, ABE)[30]。基于相似的原理,将失去活性的dCas9蛋白与胞嘧啶脱氨酶融合,也会实现在不产生DSBs的情况下将C-G碱基对替换为T-A碱基对。但由于体内存在着尿嘧啶DNA糖基化酶,该酶可识别错配的U-G错配,从而通过碱基切除修复的途径将U逆转为C。故而对比腺嘌呤碱基编辑器,胞嘧啶碱基编辑器(cytosine base editors, CBE)[31]需要在融合蛋白质上再加入尿嘧啶DNA糖基化酶抑制剂(uracil DNA glycosylase inhibitor, UGI)。单碱基编辑器的出现,不仅为CRISPR系统在临床上针对遗传疾病治疗提供了新的曙光,也说明了对CRISPR系统的改造在实际研究应用上的可行性。举例而言,在遗传性耳聋中,有80%是隐性遗传突变,需要精确的位点编辑才可使之恢复正常的生理功能。2018年,Liu等[32]团队基于单碱基编辑的CRISPR技术,运用AAV载体装载分为两部分的单碱基编辑器(AAV的运载能力限制,需将单碱基编辑器拆分为Cas9及sgRNA),而后2种AAV载体共同转染同一细胞。研究结果显示,两部分的单碱基编辑器能在体内重新结合并靶向到特异部位,且几乎无能检测到的脱靶现象。实验中原本完全丧失听觉的小鼠可对响亮的声音做出反应,表明小鼠至少已经恢复了部分听力。该单碱基编辑技术成功地于体外细胞研究中找到隐性突变位点并修复,在整体研究中通过对单碱基编辑器的重新设计再现体外细胞研究结果,证明了AAV载体能有效递送CRISPR系统,也为隐性遗传疾病的治疗提供了新思路。
通过对Cas9蛋白的改造,CRISPR系统不仅适用于靶基因的敲除,也可以利用不同机制对靶基因的表达进行双向调控。实际上,不止单碱基编辑器能在不产生DSBs的情况下对目的基因进行编辑,CRISPRi[33]、CRISPRa[34, 35]、CRISPRon[36]和CRISPRoff[37]技术同样也可以达到相同的目的。有趣的是,无论是单碱基编辑器还是上述的4个延伸的CRISPR基因编辑技术,其原理都是类似的,都是基于对dCas9蛋白的改造实现的。不同之处在于dCas9蛋白所融合的蛋白组件不同。其中,CRISPRi、CRISPRa是与转录调控因子结合,从而分别削弱或增强目的基因的表达。而CRISPRon、CRISPRoff技术则是分别与去甲基化酶和甲基化酶融合,从而在表观层面影响基因的表达。然而,现有研究对sgRNA的改造更注重于其结构与长度的改变对靶向效率的影响[38]。也有相关的研究在sgRNA上引入适体核酸组件作为开关,通过调节sgRNA的活性从而调控Cas9活性,实现人为控制CRISPR系统对靶基因编辑的时空调节[39]。
对CRISPR系统的模块进行改造不单能改进其功能,还会优化其效能。受限于Cas9系统的靶标为DNA,导致人们未能及时关注到单碱基编辑器是否会对RNA产生脱靶编辑,Grünewald等[40]率先在细胞层面检测了单碱基编辑器的RNA的脱靶编辑效应。不容乐观的是,该研究结果显示,BE碱基编辑器会在人类细胞中导致广泛的RNA胞嘧啶脱氨,频率为0.07%~100%,涉及到38%~58%的表达基因。而ABE单碱基编辑系统也存在相似的RNA脱靶编辑效应。面对单碱基编辑器导致RNA层面的脱靶效应弊端,Grünewald等对胞嘧啶脱氨酶进行改造,保留了单碱基编辑器对目的基因的编辑效能,显著减少RNA脱靶编辑,为单碱基编辑系统的优化指出了方向。随着研究的进展,人们不再满足单碱基编辑器较低的编辑效率和编辑窗口,Zhang等[41]开发了双碱基编辑器,该研究将胞嘧啶脱氨酶hAID-腺嘌呤脱氨酶-Cas9n融合在一起,实现在同一基因上单独的C>T和A>G,或者二者的同时进行。Grünewald等[42]将腺苷脱氨酶和胞嘧啶脱氨酶分别融合到Cas9切口酶的N-端和C-端,开发了一种可同时引入A>G和C>T替换的双碱基编辑器。目前,更多的双碱基编辑器也在开发应用中。随着研究的深入,碱基编辑器或能为治疗遗传疾病、癌症等难治性疾病开创新的治疗途径。
4 CRISPR系统的局限及其优化改造
CRISPR/Cas系统在进行基因编辑时存在着脱靶编辑,PAM区识别限制以及递送困难等障碍,能否通过改造CRISPR元件来减少实际应用中的障碍是临床研究中亟待解决的问题之一。CRISPR/Cas9系统是细菌的一种获得性的免疫防御系统,在对外源DNA的清除过程中为保证较高清除率,系统本身就存在一定的错配忍受性。且哺乳动物细胞基因组大小远超细菌,这意味着,在使用CRISPR系统用于哺乳动物基因编辑时脱靶概率的相应提升[43]。随着对CRISPR系统的了解,发现对Cas9蛋白或sgRNA的优化或改造可有效减少脱靶编辑。对Cas9蛋白的优化的主要方式为设计其类似物或者对其进行改造。Kleinstiver 等[44]团队通过编辑spCas9蛋白与非靶基因结合的关键氨基酸,构建了增强型特异性的蛋白变体“eSpCas9”,该变体的脱靶性相较于原蛋白质得到显著降低。而对sgRNA的改造主要针对GC含量、长度及化学修饰等方式来降低脱靶率,用已有相关的数据算法来预测sgRNA的脱靶率,并给出sgRNA设计的较优序列[43]。
此外,Cas9蛋白在发挥核酸内切酶功能时,还需识别靶序列的PAM,因此CRISPR系统并非能自由编辑所有序列,可被编辑的序列必须存在PAM区。令人振奋的是,Kleinstiver等[45]团队对SpCas9进行升级改造,使之成为了几乎不受PAM限制的变体蛋白SpRY,其可识别的PAM序列为NRN和NYN,显著扩展了CRISPR系统的编辑窗口。CRISPR系统在基因疗法中的病毒递送载体通常为低免疫原性的腺相关病毒(adeno-associated virus, AAV),但AAV载体的装载能力是有限的。SpCas9的氨基酸长度为1 368,而在金黄色葡萄球菌中发现的saCas9的氨基酸长度仅为1 058,更适合包装成AAV病毒。寻找长度更短,分子量更小的Cas9蛋白类似物,是解决CRISPR系统递送困难的方案之一,但不改变spCas9原有功能的条件下对其进行截短可能也是潜在的优化方式。
5 问题与展望
CRISPR基因编辑系统对了解生物体内关键基因功能,潜在药物靶点的筛选以及不同基因转录背景下基因表达的联系等研究具有巨大的价值。随着对CRISPR组分结构和功能的解析,基于CRISPR系统的改造研究也在不断深化。通过对Cas9蛋白的改造,CRISPR系统不再只能实现基因敲除,其还能在转录层面以及表观遗传调控水平实现对靶基因的双向调控,甚至能用于活体细胞成像。CRISPR系统的改造也显著降低了其原先脱靶率高、递送困难等应用障碍。但将CRISPR系统真正转化到实际生产与临床应用时,仍需关注到改造后的系统是否存在新的以往未知的安全性隐患,系统本身已知的风险是否可被接受,实际的研究能否做到透明公开,是否通过了伦理审查等。随着研究的不断深化进展,CRISPR系统的改造也会不断深化,进而反哺和推动临床医学研究与应用的发展。
