东革内酯通过InR-PI3K-mTOR-Bcl-2通路诱导斜纹夜蛾卵巢细胞凋亡
2023-02-03温雪梅秦子昕黄建辉欧阳嘉敏邵雪花
温雪梅, 秦子昕, 路 伟, 黄建辉,欧阳嘉敏, 万 妍, 邵雪花
(1.广东省农业科学院果树研究所, 农业农村部南亚热带果树生物学与遗传资源利用重点实验室, 广东省热带亚热带果树研究重点实验室, 广州 510640; 2.新疆农业大学农学院, 棉花教育部工程研究中心,自治区农林有害生物监测与安全防控重点实验室, 乌鲁木齐 830052; 3.广东省中山市坦洲镇农业服务中心, 中山 528400)
斜纹夜蛾Spodopteralitura(Fabricius)是一种世界性分布的农业害虫,其幼虫可为害十字花科、茄科、豆科以及花卉等300多种植物[1-2]。其繁殖力高、生命周期短和迁移能力强等特点增加了田间防治难度[3],化学防治为其主要防治手段,但随着人们对农产品质量安全的重视,对绿色食品、有机食品的需求越来越多,这为生物农药的发展带来新的契机。因此,挖掘靶向斜纹夜蛾的高活性天然产物非常重要。
东革内酯(eurycomalactone,EC)来源于东革阿里Eurycomalongifolia(Jack)根部。该类化合物是一类具有苦味的降三萜类物质,不仅在医学上具有抗疟疾[4]、抗肿瘤、抗癌[5-7]、抗炎、抗病毒[8]等药理作用,在农业上亦对多种害虫具有拒食和毒杀等生物活性。从印度黄楝树Samaderaindica种子和树皮中分离的4个苦木苦味素类化合物(indaquassin C和samaderines A、B、C)对斜纹夜蛾幼虫具有良好的拒食作用[9];从牛筋果Harrisoniaperforata(Blanco)枝叶中分离得到苦木苦味素类化合物perforalactone A和perforalactone B对蚜虫具有较好的毒杀活性[10];从鸦胆子Bruceajavanica(Linn)中分离得到的苦木苦味素类化合物鸦胆因D对小菜蛾Plutellaxylostella的拒食活性为印楝素的6.2倍[11]。近期,作者从东革阿里中分离得到了东革内酯、宽缨酮(eurycomanone)、13,21-dihydroeurycomanone、11-dehydroklaineanone、15β-hydroxyklaineanone、eurycolactone F、6α-hydroxyeurycomalactone等7个苦木苦味素类化合物。研究表明,宽缨酮对草地贪夜蛾Spodopterafrugiperda、斜纹夜蛾、小菜蛾及甜菜夜蛾Spodopteraexigua等鳞翅目害虫的拒食及抑制生长活性与印楝素相当或更高,同时兼具良好的植物内吸特性[12-18]。东革内酯具有抗癌、抗炎及抗病毒等药理作用,临床上常被作为有效的核因子κB(NF-κB)抑制剂使用[19],但其是否可抑制昆虫细胞增殖及毒理机制目前尚未见报道。基于此,本文以斜纹夜蛾卵巢细胞SL-221为研究对象,采用CCK-8法检测不同浓度东革内酯对SL-221细胞的毒力,并通过流式细胞术测定该化合物对SL-221细胞周期、线粒体膜电位及凋亡的影响;最后,利用实时荧光定量PCR(qPCR)技术探究东革内酯处理SL-221细胞后相关基因的表达情况,旨在探究东革内酯诱导SL-221细胞发生凋亡的分子机制,为斜纹夜蛾的绿色防控和东革内酯的开发利用提供新的视角。
1 材料与方法
1.1 供试材料
1.1.1供试细胞
斜纹夜蛾卵巢细胞SL-221,由广东省农业科学院果树研究所资源与环境实验室提供,用SIM SF昆虫细胞培养基(含5%胎牛血清),在27℃恒温箱中培养。
1.1.2供试药剂
东革内酯(eurycomalactone,EC),分子量348,CAS号:23062-24-0,由广东省农业科学院果树研究所资源与环境实验室提供,结构式见图1。

图1 东革内酯的结构式
1.1.3供试试剂和仪器
供试试剂及试剂盒:胎牛血清购自蒙马生物科技有限公司;SIM SF Expression Medium(MSF1)、CCK-8(M128)、细胞周期与细胞凋亡检测试剂盒(C1052)、Annexin Ⅴ-FITC/PI凋亡检测试剂盒(CA1020)、线粒体膜电位检测试剂盒(M8650)和二甲基亚砜(dimethyl sulfoxide,DMSO)均购自北京索莱宝科技有限公司;RNA提取试剂盒、呲溜极速反转录试剂盒和2×M5 HiPer Realtime PCR Super mix with Low Rox(SYBR green with anti-Taq)(MF797-05)均购自北京聚合美生物科技有限公司;PBS缓冲液购自博奥森生物技术有限公司;印楝素,纯度≥98%,由华南农业大学亚热带农业生物资源保护与利用国家重点实验室提供。
供试仪器:恒温培养箱(DH-360)购自北京科伟永兴仪器有限公司;多功能酶标仪(Spark)购自上海帝肯实验器材有限公司;流式细胞仪(FACS Verse)购自上海土森视觉科技有限公司;荧光定量PCR仪(Quant Studio 3)购自赛默飞世尔科技有限公司。
1.2 试验方法
1.2.1CCK-8法检测东革内酯对SL-221细胞的毒力
将冻存的SL-221细胞置于37℃水浴迅速搅拌融化,在超净工作台内将细胞液转入2 mL离心管,800 r/min离心5 min,弃上清,用2 mL含10%胎牛血清的SIM SF昆虫细胞培养基重悬细胞,将细胞悬液转于细胞培养瓶,27℃恒温箱中培养。用DMSO将东革内酯配制成10 mg/mL的母液,并用无血清SIM SF培养基逐级稀释至100、50、25、12.5、6.25、3.125 μg/mL和1.562 5 μg/mL。取对数生长期的SL-221细胞,用含5%胎牛血清的SIM SF培养基将细胞浓度调节至5×105cfu/mL,接种于96孔板,每孔100 μL,置于27℃恒温箱过夜。试验前弃去96孔板中原有的含血清培养基,分别加入各浓度含药培养基,每浓度4板重复,以含0.1% DMSO的无血清SIM SF培养基作为空白对照,以相同浓度印楝素处理作为阳性对照,置于27℃恒温箱孵育48 h。48 h后向每孔中加入10 μL CCK-8溶液,继续避光孵育2~3 h。最后用酶标仪测定其在450 nm处的吸光度,根据公式计算细胞抑制率,并利用SPSS软件计算其抑制中浓度(IC50)。
细胞增殖抑制率=(A零加药-A加药)/(A零加药-A空白)×100%,式中,A加药为含有SL-221细胞、含药培养基和CCK-8溶液孔的吸光度;A空白为含有含药培养基和CCK-8溶液孔的吸光度;A零加药为含SL-221细胞、CCK-8溶液但无药剂孔的吸光度。
1.2.2东革内酯对SL-221细胞形态的影响
用DMSO将东革内酯配制成10 mg/mL母液,在无血清SIM SF昆虫细胞培养基中添加东革内酯使其终浓度为4 μg/mL。用含5%胎牛血清的SIM SF培养基将SL-221细胞培养至对数生长期(5×105cfu/mL),取2 mL接种于细胞培养皿(d=6 cm)中,置于27℃恒温箱培养。待细胞密度达到85%~90%时,弃去原有培养基,更换为含4 μg/mL东革内酯的SIM SF培养基,培养0、3、6、12、24 h和48 h后在倒置显微镜下观察,其中0 h为对照。拍照记录细胞形态变化。
1.2.3流式细胞术检测东革内酯对SL-221细胞周期的影响
用DMSO将东革内酯配制成10 mg/mL母液,进而配制含1 μg/mL东革内酯的无血清SIM SF培养基。取对数生长期的SL-221细胞(5×105cfu/mL)接种至12孔板,每孔2 mL,于27℃恒温箱培养。待细胞密度达到85%~90%时,弃去原有培养基,加入含1 μg/mL东革内酯的SIM SF培养基,培养0、3、6、12、24 h和48 h,每个时间点设置3个重复孔, 试验重复3次,于27℃恒温箱中分别孵育0、3、6、12、24 h和48 h,其中0 h为对照。之后收集细胞,800 r/min离心5 min,弃上清。用PBS洗涤细胞,然后加入预冷的70%乙醇置于4℃冰箱过夜。次日用PBS洗涤细胞2次,加入100 μL RNaseA溶液重悬细胞,37℃水浴30 min;之后加入400 μL PI染色液,4℃避光孵育30 min。完成后立即用流式细胞仪在激发波长488 nm处检测红色荧光,用FlowJo软件统计分析处于G0/G1(DNA合成前期)、S(DNA合成期)和G2/M(DNA合成后期)的细胞数量。
1.2.4JC-1荧光标记法检测东革内酯对SL-221细胞线粒体膜电位的影响
细胞处理方法和重复情况同1.2.3。收集1 μg/mL东革内酯分别处理0、3、6、12、24 h和48 h后的细胞,用PBS清洗细胞2次。之后加入0.5 mL JC-1染色工作液,37℃避光孵育20 min;孵育结束后,600 r/min离心4 min,弃上清,用JC-1染色缓冲液(1×)清洗细胞2次;再加入1 mL JC-1染色缓冲液(1×)重悬细胞。最后置于流式细胞仪检测激发波长585 nm,发射波长590 nm处的PE(红色)荧光;激发波长515 nm,发射波长529 nm处的FITC(绿色)荧光,并计算红绿荧光的相对比例来衡量线粒体膜电位变化情况(红绿荧光的相对比例=红色荧光百分比/绿色荧光百分比)。
1.2.5Annexin Ⅴ-FITC/PI双染检测东革内酯对SL-221细胞凋亡的影响
Annexin Ⅴ-FITC/PI凋亡检测试剂盒是一种采用Annexin Ⅴ-FITC与PI双染法进行细胞早期凋亡分析的检测试剂盒。正常细胞具有完好的细胞膜,此时细胞膜中的磷脂酰丝氨酸(phosphotidylserine,PS)位于胞膜内侧。细胞早期凋亡时,细胞膜结构发生改变,使胞膜内侧的PS外翻到胞膜表面。根据Annexin Ⅴ易于结合PS并对其高度亲和的特性可检测凋亡的细胞。荧光染料碘化丙啶(propidiumiodide,PI)不能通过正常细胞,但能对凋亡晚期的细胞进行染色,因此将Annexin Ⅴ与PI配合染色,以区别凋亡早期细胞和凋亡晚期细胞。细胞处理方法和重复情况同1.2.3。收集1 μg/mL东革内酯分别处理0、3、6、12、24 h和48 h后的细胞,用预冷的PBS清洗细胞2次,800 r/min离心5 min,弃上清;加入250 μL Binding Buffer重悬细胞;取100 μL细胞悬液放入流式管中,按照试剂盒说明书加入5 μL Annexin Ⅴ-FITC和10 μL PI溶液,轻摇混匀,避光孵育15 min。置于流式细胞仪检测激发波长535 nm,发射波长615 nm处的PI(红色)荧光;激发波长488 nm,发射波长525 nm处的FITC(绿色)荧光,并用FlowJo软件分析细胞凋亡情况。
1.2.6RT-qPCR法检测东革内酯对SL-221细胞相关基因表达情况的影响
细胞处理方法同1.2.3,以SL-Actin作为内参基因,引物序列见表1,由生工生物工程(上海)股份有限公司合成。收集1、2 μg/mL和4 μg/mL东革内酯分别处理0、3、6、12、24 h和48 h后的SL-221细胞,其中0 h为对照,利用RNA提取试剂盒提取其总RNA,再通过反转录试剂盒合成cDNA。反应体系(20 μL):5×Sprint gDNA Remover Mix 2 μL、RNA模板1 μL、DEPC-ddH2O 7 μL,将反应液混匀,42℃温育2 min,冰上冷却;再加入5×M5 Sprint RT Mix 4 μL、DEPC-ddH2O 6 μL,50℃ 5 min、85℃ 5 s。利用SYBR green with anti-Taq试剂盒进行PCR扩增,反应体系(10 μL):cDNA 1 μL、2×M5 HiPer Realtime PCR Super mix with Low Rox 5 μL、Primer 0.5 μL,RNase Free ddH2O 3.5 μL。反应程序为:95℃预变性 30 s;95℃ 5 s,60℃ 30 s,40个循环;95℃ 15 s,60℃ 1 min,95℃ 15 s。每样品重复3次,通过荧光定量PCR得到目的基因与内参基因的Ct值,利用2-ΔΔCt法(△Ct=Ct目的基因-Ct内参基因)计算不同处理时间下SL-221细胞相关基因的相对表达量。

表1 斜纹夜蛾生长发育相关基因 RT-qPCR 引物序列
1.3 数据分析
本试验结果均为3次以上重复试验所得,运用IBM SPSS Statistics 19软件分析数据,各处理组所得平均值之间的差异通过单因素方差分析及Duncan氏多重比较进行检验评估,P<0.05表示在统计学上具有显著性差异,使用ImageJ软件处理细胞图片,FlowJo软件分析细胞流式数据,Origin 2017进行绘图。
2 结果与分析
2.1 CCK-8法检测东革内酯对SL-221细胞的毒力
检测结果(表2)显示,东革内酯对SL-221细胞48 h的IC50为1.98 μg/mL,而阳性对照印楝素48 h的IC50为7.51 μg/mL,东革内酯对SL-221细胞有明显的抑制细胞增殖的活性。浓度为1.562 5、3.125、6.25、12.5、25、50 μg/mL和100 μg/mL的东革内酯处理48 h的细胞增殖抑制率分别为43.87%、58.96%、66.52%、74.42%、79.41%、81.51%和86.97%(图2)。东革内酯对SL-221细胞的抑制增殖活性与浓度呈正相关。

表2 东革内酯对SL-221细胞的毒力
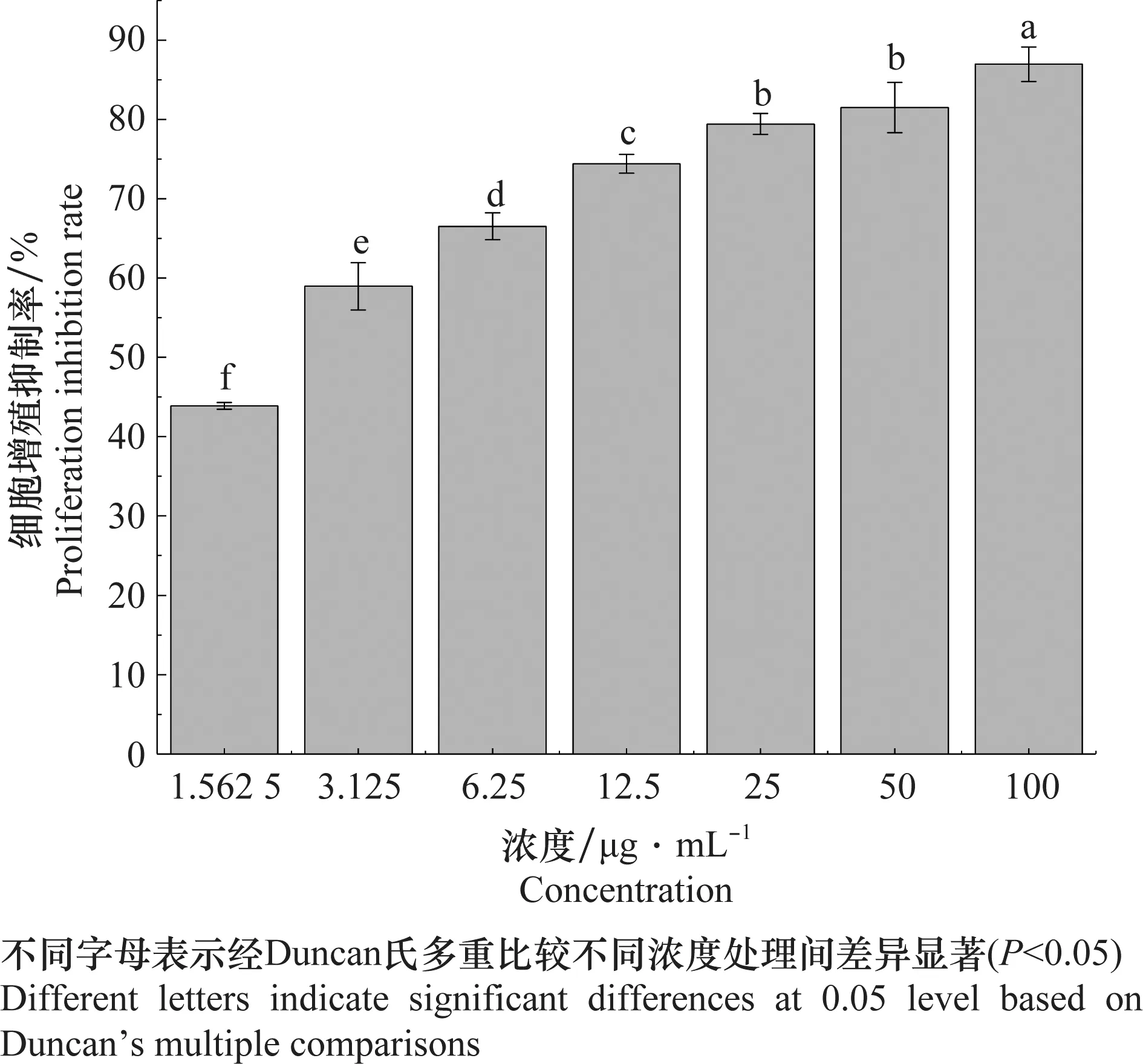
图2 东革内酯处理SL-221细胞48 h的细胞增殖抑制率
2.2 东革内酯处理后SL-221细胞形态变化
经4 μg/mL东革内酯处理后SL-221细胞形态出现了明显变化。从图3可以看出,0 h(对照)时,SL-221细胞形态饱满光滑,呈圆形或椭圆形;处理 3 h 和6 h后,细胞膜光滑度下降,部分细胞膨胀;12 h后,细胞膨胀明显,细胞膜表面粗糙;处理24 h后,细胞膜粗糙,细胞肿胀破裂,并可见凋亡小体(图3e);48 h后,细胞内容物溢出,凋亡小体数量明显增多,大部分细胞不能维持贴壁(细胞培养瓶底)生长。综上,随着药剂处理时间延长,细胞凋亡现象明显,细胞活力下降,东革内酯对SL-221细胞增殖的抑制活性呈现时间依赖性。
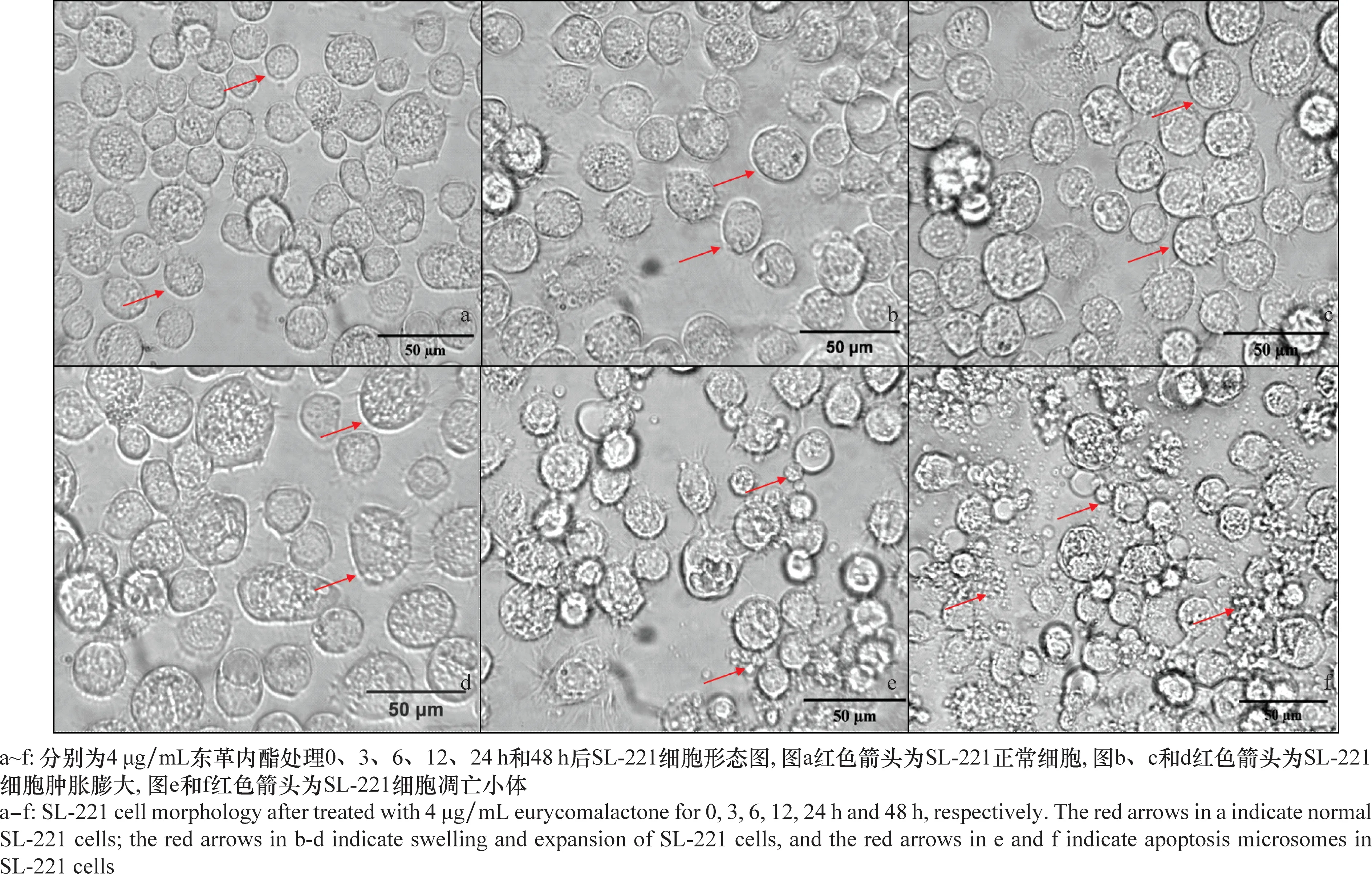
图3 4 μg/mL东革内酯处理后SL-221细胞形态图
2.3 流式细胞术检测东革内酯对SL-221细胞周期的影响
经1 μg/mL东革内酯处理0、3、6、12、24 h和48 h的SL-221细胞中处于不同周期的细胞百分率的分布情况见图4 a,其统计结果见图4 b。从结果可以看出,与0 h(对照)相比,12、24 h和48 h的G0/G1期细胞百分率均显著下降P<0.05),而G2/M期细胞百分率均显著上升(P<0.05)。其中,处理48 h的G0/G1期细胞所占百分率从(35.80±3.58)%下降至(20.67±2.10)%,S期细胞所占百分率由(33.47±8.34)%下降至(16.90±3.03)%,而G2/M期细胞所占百分率从(30.47±5.43)%增加至(62.67±0.67)%。由此可见,东革内酯可将SL-221细胞周期阻滞于G2/M期。

图4 东革内酯对SL-221细胞周期的影响
2.4 JC-1荧光标记法检测东革内酯对SL-221细胞线粒体膜电位的影响
经1 μg/mL东革内酯处理不同时间的SL-221细胞线粒体膜电位的分布情况见图5a。在线粒体膜电位较高时,JC-1聚集形成聚合物,可以产生红色荧光;在线粒体膜电位较低时,JC-1为单体,可以产生绿色荧光。SL-221细胞线粒体膜电位变化情况的统计结果见图5 b,随着处理时间的增加,SL-221细胞线粒体红/绿荧光比例显著下降(P<0.05)。0 h(对照)SL-221细胞线粒体红/绿荧光比例为11.55,处理3、6、12、24 h和48 h时,SL-221细胞线粒体红/绿荧光比例分别为3.51、1.40、0.33、0.29和0.19,与0 h(对照)相比下降了69.61%、87.88%、97.14%、97.49%和98.35%。可见,东革内酯可显著降低SL-221细胞的线粒体膜电位,且呈现时间依赖性。
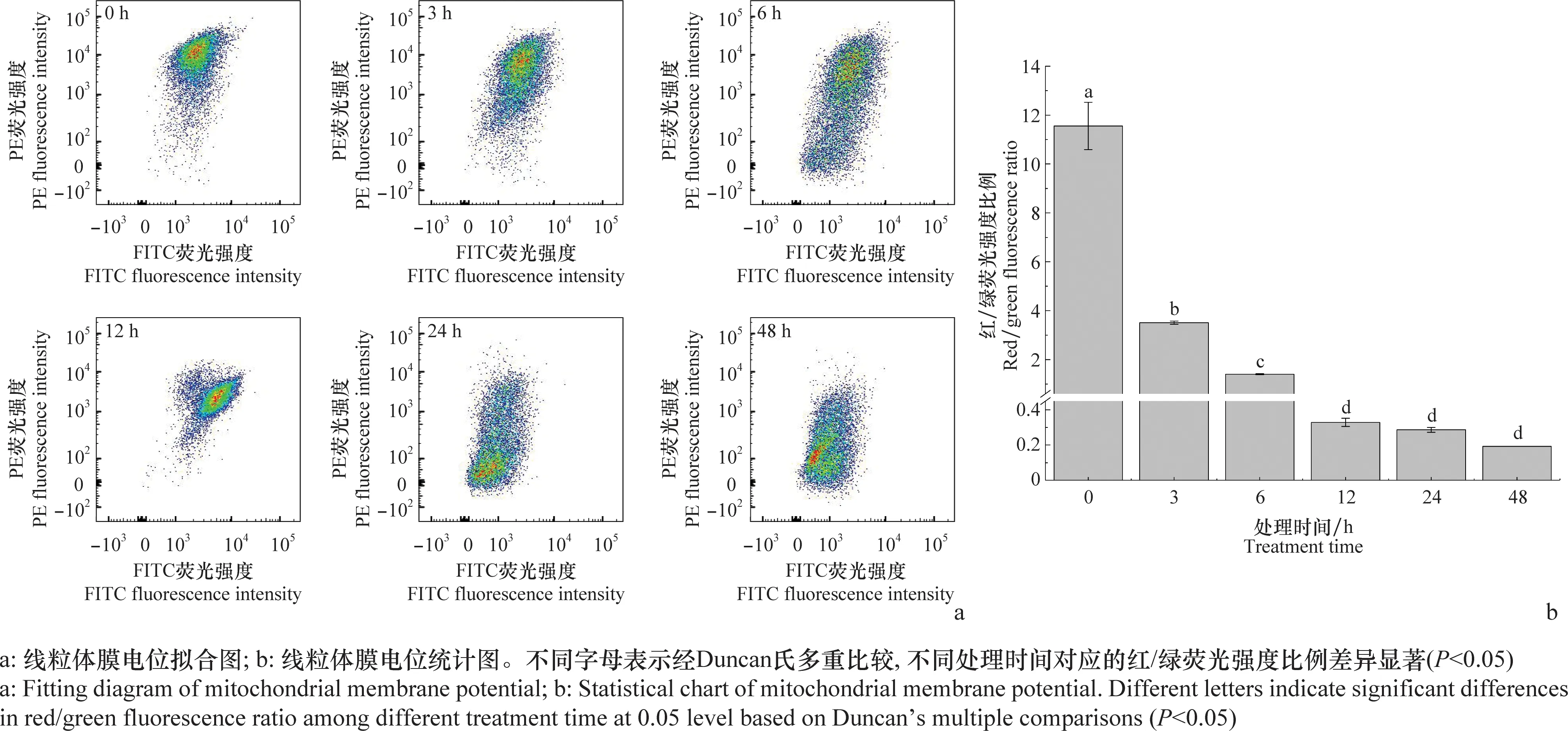
图5 东革内酯对SL-221细胞线粒体膜电位的影响
2.5 Annexin Ⅴ-FITC/PI双染检测东革内酯对SL-221细胞凋亡的影响
Annexin Ⅴ-FITC/PI双染检测1 μg/mL东革内酯处理不同时间下,不同类型的SL-221细胞的分布情况见图6 a。其中Q1象限为坏死细胞,Q2和Q3象限分别为晚期凋亡细胞和早期凋亡细胞,Q4为正常细胞。Annexin Ⅴ-FITC/PI双染统计结果见图6 b。结果显示,东革内酯处理不同时间下,SL-221正常细胞百分率与0 h(对照)相比显著下降(P<0.05),而早期凋亡细胞百分率显著上升(P<0.05)。其中,处理48 h与0 h相比,SL-221正常细胞百分率由75.47%降低至32.80%,早期凋亡细胞百分率由8.42%增加到32.03%,晚期凋亡细胞百分率由11.43%增加到33.13%。由此可见,东革内酯可诱导SL-221细胞凋亡,且随着时间的延长凋亡加剧。

图6 东革内酯对SL-221细胞凋亡的影响
2.6 RT-qPCR法检测东革内酯对SL-221细胞凋亡相关基因表达情况的影响
1、2、4 μg/mL东革内酯处理SL-221细胞后,SL-p53、SL-CytochromeC和SL-InR基因表达均呈现上调趋势(图7),处理48 h,SL-p53基因表达与对照相比分别上调17.53、67.92倍和90.10倍;SL-CytochromeC基因表达与对照相比分别上调38.03、65.15倍和74.15倍;SL-InR基因表达与对照相比分别上调2.34、4.19倍和4.34倍。SL-Bcl-2、SL-mTOR和SL-PI3K基因的表达量随处理时间的延长均呈现下降趋势。其中,1、2、4 μg/mL东革内酯分别处理48 h后,与对照相比,SL-Bcl-2基因表达分别下调94.76%、96.49%和96.94%;SL-mTOR基因表达与对照相比分别下调96.10%、97.30%和98.14%;SL-PI3K基因表达与对照相比分别下调86.55%、87.01%和93.05%。
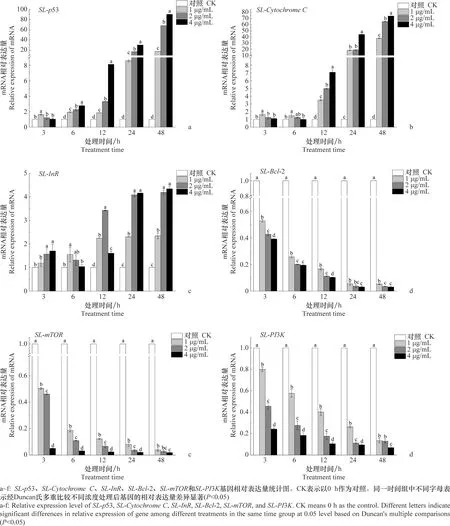
图7 东革内酯对SL-221细胞凋亡相关基因表达情况的影响
综上,1、2、4 μg/mL东革内酯可诱导凋亡标志基因SL-p53、SL-InR和SL-CytochromeC显著上调表达(P<0.05),同时抑制凋亡抑制因子SL-Bcl-2的表达。细胞凋亡信号通路上游的SL-mTOR和SL-PI3K基因表达均受到显著抑制(P<0.05)。表明东革内酯可通过InR-PI3K-mTOR-Bcl-2信号通路诱导SL-221细胞凋亡。
3 结论与讨论
斜纹夜蛾属于重大农业害虫,目前主要依靠化学农药进行防控,但监测表明其已对多种化学杀虫剂产生了抗性[20-21]。因此,寻找高效低毒的药物及靶点是农药领域研究的热点和难点。从天然产物中挖掘新活性物质,研发替代防治产品至关重要。本文比较了印楝素与东革内酯对SL-221细胞增殖的抑制作用,发现东革内酯能以更小的剂量(IC50为1.98 μg/mL)抑制SL-221细胞增殖,随着东革内酯浓度增高,SL-221细胞数目减少,细胞膨胀、核固缩(细胞核染色质DNA浓聚、皱缩,使核体积减小)破碎,细胞碎片增多,提示其可能具有开发为防控斜纹夜蛾的药物的潜力。东革内酯能否通过诱导凋亡来抑制SL-221细胞增殖,目前仍不清楚。我们通过流式细胞术发现,东革内酯能使SL-221的细胞周期停滞于G2/M期,降低线粒体膜电位,且能显著诱导细胞凋亡。
细胞凋亡(apoptosis)是一种特殊的细胞死亡方式,又被称为程序性细胞死亡(programmed cell death, PCD),指为维持内环境稳定,由基因控制的细胞自主有序的主动死亡过程[22]。在形态方面,细胞凋亡表现为:细胞质浓缩、染色质聚集、核酸内切酶活化和出现凋亡小体等特征。发生过程包括:凋亡的起始、凋亡小体形成和凋亡小体被邻近细胞吞噬并消化[23]。通常情况下,细胞凋亡是机体为了维持各器官的稳定性而进行的一种自我调控,但当细胞凋亡调控失衡时,可引起细胞过度增殖或过度凋亡,导致相关疾病的发生[24]。细胞凋亡主要有内源性和外源性两种途径。内源性途径主要包括细胞应激、DNA损伤、发育信号、存活因子缺失等[25]。这个途径是由Bcl-2家族介导,该家族存在两种类型的蛋白,促凋亡蛋白(Bax、Bad等)和抑凋亡蛋白(Bcl-2)[26]。在内源性途径中,CytochromeC的表达对细胞凋亡起着关键作用[27]。CytochromeC从线粒体释放到胞质中,形成凋亡复合体,进而激活核酸内切酶切割DNA,促使细胞凋亡[28]。p53是一种肿瘤抑制因子,它可以对细胞应激或DNA损伤作出响应,被激活后的p53可诱导细胞的周期停滞、凋亡和衰老[29-30]。当蛋白表达异常时可能促使细胞进入不可逆的凋亡阶段。我们发现东革内酯可抑制凋亡因子SL-Bcl-2的表达,促进SL-p53、SL-InR和SL-CytochromeC基因的表达。InR-PI3K/mTOR是一条经典的胞内信号通路,主要调节细胞的增殖、自噬和凋亡等,其异常活化或被抑制均会诱导下游信号途径的紊乱。PI3K是该信号通路的重要组成因子,可在多种生长因子的刺激下被激活,参与细胞自噬的调节,降低细胞自噬力[31-32]。mTOR位于PI3K/mTOR信号通路的下游,属于磷酸肌醇激酶家族成员,PI3K激活后,其下游的mTOR也被激活,mTOR激活可抑制细胞自噬[33]。本研究表明SL-mTOR和SL-PI3K基因表达均显著下调(P<0.05),进一步证实了东革内酯可诱导SL-221细胞凋亡,这与印楝素可诱导SL-1细胞凋亡的结果一致[34]。
近年来研究表明,活性成分可通过上调InR,负反馈抑制PI3K/Akt通路来诱导细胞凋亡[35]。本试验中,我们发现东革内酯可显著上调InR基因的表达,同时抑制PI3K和mTOR基因的表达,因此,我们推测东革内酯诱导SL-221细胞发生的凋亡可能与抑制PI3K和mTOR的磷酸化、抑制Bcl-2的表达,促使细胞凋亡有关。
综上所述,东革内酯可通过InR-PI3K-mTOR-Bcl-2信号通路诱导SL-221细胞发生凋亡,后续将进一步研究东革内酯是否还通过诱导其他死亡方式发挥抑制昆虫细胞增殖及其具体机制,为东革内酯的开发和利用提供新的依据和思路。
