棉铃虫性信息素生物合成研究进展
2023-02-03卢雯慧魏纪珍安世恒
李 祥, 卢雯慧, 魏纪珍, 安世恒
(河南农业大学植物保护学院, 小麦玉米作物学国家重点实验室, 河南省害虫绿色防控国际联合实验室, 郑州 450002)
棉铃虫Helicoverpaarmigera(Hübner)属鳞翅目Lepidoptera夜蛾科Noctuidae,是一种世界性农业害虫。该虫适应能力强且寄主广泛,当外部环境适宜时常大面积暴发成灾[1-2]。棉铃虫曾是我国和世界棉花种植产业的重要威胁[3],后来随着转基因抗虫棉的大面积推广,在一定程度上抵御了棉铃虫的为害[4];但是除棉花外,棉铃虫还为害玉米、高粱、小麦等粮食作物,以及辣椒、番茄、菜豆等蔬菜,其寄主植物多达30多科200余种作物[5-6]。我国幅员辽阔、地形复杂,农作物种植面积广、种类多、分布杂,农民对害虫的防治水平参差不齐,这都为棉铃虫的持续为害提供了便利。
化学杀虫剂是当前防治棉铃虫的重要手段,但长期、大量、不合理地使用化学农药已经造成了严重的生态污染和人畜安全问题。在过去的十多年间,全国农业技术推广服务中心连续对华北棉区的棉铃虫进行了抗性监测,发现该虫对拟除虫菊酯类杀虫剂的抗药性已达到高抗水平(抗性倍数为113~342倍),对有机磷类(如辛硫磷)、双酰胺类(如氯虫苯甲酰胺)和大环内酯类药剂(如甲氨基阿维菌素苯甲酸盐)的抗性处于中等水平,并且这些抗性水平还在不断上升[7-11]。与化学杀虫剂相比,Bt杀虫蛋白具有靶向性强、不杀伤天敌、对环境友好等优点,转Bt基因棉花累计种植面积已超过10亿hm2[4]。但是在2008年时,美洲棉铃虫Helicoverpazea对表达Cry1Ac杀虫蛋白的棉花产生了抗性,这成为首个Bt作物防治失败的案例[12]。此后,转Bt基因棉田间防治失败的案例不断增加[13-14],甚至在不同蛋白之间出现了交互抗性[15]。近年来,棉铃虫对Cry1Ac的抗性也显著提高[16-17]。鉴于棉铃虫对化学农药和转Bt基因作物的抗性问题日益严重,迫切需要新的治理手段和更为安全有效的综合防治策略。
大多数昆虫繁衍后代进行物种延续时是通过两性交配来完成的,这需要同一物种两性个体的成功相聚。性信息素(sex pheromone)是昆虫两性通讯的化学信号,在物种繁衍中担当着重要的角色。昆虫性信息素又称性外激素,是由特殊腺体所分泌的一类微量化学物质,对同种异性个体具有远距离引诱和促进交配的作用[18]。鳞翅目蛾类通常依赖雌蛾释放的性信息素作为远距离信息交流的化学信号,这种信号是雄蛾成功搜寻到配偶的关键因子。在求偶期,雌蛾通过延伸产卵器末端来暴露其性信息素腺体(pheromone gland,PG),并释放性信息素;雄蛾能够感知这些化学物质并做出定向反应来完成交配。以性信息素为基础的害虫诱杀和交配干扰等防治手段具有灵敏度高、选择性强、对天敌安全、不污染环境等优点,在害虫综合治理和绿色防控体系应用中深受青睐。本研究概述了棉铃虫性信息素的组成和功能,生物合成时的信号转导途径和调控机制,以及性信息素前体的生成和特异性组分的碳链修饰过程,并总结了当前研究的不足,旨在为今后的基础研究和生产应用提供指导。
1 棉铃虫性信息素组分的鉴定和功能
对昆虫性信息素的成分鉴定始于20世纪50年代,Butenandt等从50万头家蚕Bombyxmori雌蛾中分离并鉴定出了第一个昆虫性信息素——反8,顺10-十六碳二烯-1-醇(trans-8,cis-10-hexadecadien-1-ol,E8,Z10-16∶OH)[19],由此开始了研究昆虫性信息素化学结构的时代。鳞翅目昆虫是农业上为害较重的一类害虫,这类昆虫视觉很差,但拥有发达的嗅觉交流机制,是昆虫性信息素研究的主要对象。目前已有超过700种蛾类的性信息素组分被鉴定出来(https:∥lepipheromone.sakura.ne.jp/pdb_top.html;http:∥www.pherobase.com/)。其中,大约75%的蛾类性信息素是长度在10~18C的直链脂肪族化合物,具有0~3个不饱和键,末端具有羟基、醛基或乙酰基等官能团,称为醇类、醛类或酯类性信息素;具有这样特征的蛾类性信息素也被定义为I型性信息素[20]。
以棉铃虫为代表的醛类性信息素的生物合成和调控过程,是目前研究较早也较为透彻的一类性信息素。1977年,首个被鉴定出的棉铃虫性信息素组分为顺-11-十六碳烯醛(cis-11-hexadecenal,Z11-16∶Ald),是从雌蛾第八到第九腹节的节间膜处提取到的[21]。随后通过固相微萃取和溶剂浸提的方法先后从PG中鉴定出了其他9种组分,包括顺-11-十六碳烯醇(cis-11-hexadecenol,Z11-16∶OH)、顺-9-十六碳烯醛(cis-9-hexadecenal,Z9-16∶Ald)、顺-7-十六碳烯醛(cis-7-hexadecenal,Z7-16∶Ald)、十六醛(hexadecanal,16∶Ald)、十六醇(hexadecanol,16∶OH)、顺-9-十四碳烯醛(cis-9-tetradecenal,Z9-14∶Ald)、十四醛(tetradecanal,14∶Ald)、壬醛(nonanal,9∶Ald)和庚醛(heptanal,7∶Ald)[22-26]。
这10种化合物的含量在棉铃虫不同地理种群之间略有差异。其中,Z11-16∶Ald的含量显著高于其他组分,占性信息素总混合物的90%左右。大量室内和田间试验也证明了该物质是棉铃虫雌蛾吸引异性的关键物质[21-24, 27-28]。实际上单一的Z11-16∶Ald对雄蛾的吸引力并不高,加入1%~10%的Z9-16∶Ald后能大大提高引诱效果[24, 29]。这两种物质的混合被认为是棉铃虫性信息素的主要组分。通常,Z11-16∶Ald和Z9-16∶Ald的比例为97∶3时,引诱效果最佳[28]。有学者认为,其他几种组分可能是性信息素生物合成过程中的中间产物或副产物。但后来的研究发现,9∶Ald、Z9-14∶Ald、Z7-16∶Ald、Z11-16∶OH等物质不仅能够引起雄蛾触角强烈的电生理反应,将这些组分添加到棉铃虫性信息素主成分后,其引诱效果也发生了不同情况的变化[23-24, 30]。例如,在Z11-16∶Ald与Z9-16∶Ald(97∶3)的混合物中加入适量(0.3%)的Z9-14∶Ald能大幅提升混合物对雄蛾的引诱能力[24];但添加较高含量的Z9-14∶Ald或Z11-16∶OH则会显著抑制诱蛾效果[23, 28]。进一步研究发现,Z11-16∶OH主要在雌蛾羽化初期时大量释放,以阻止雄蛾与尚未性成熟的雌蛾交配[30-31]。Z7-16∶Ald不能提高雌蛾对雄蛾的远距离引诱效果,但在两性成虫近距离接触时,Z7-16∶Ald的存在能促进种群交配率的提高[23]。其他化合物的具体生物学意义,还有待进一步的研究和发掘。
2 棉铃虫性信息素生物合成的信号转导
棉铃虫雌蛾通常在羽化后2~3 d的暗期4.5 h左右性信息素释放达到最高水平,求偶和交配高峰则发生在性信息素大量释放后的2~3 h[32]。昆虫交配需要在特定的发育阶段和光周期条件下进行,因此,性信息素的生物合成过程也需要被精准地调控。直接激发并控制棉铃虫性信息素生物合成的因子是性信息素合成激活肽(pheromone biosynthesis-activating neuropeptide,PBAN),这是一种由33个氨基酸组成的神经激素,在蛾类昆虫的进化中高度保守[33-34]。大多数蛾类性信息素的合成和释放都受PBAN调控,这种调控作用会受到内部神经系统以及外部环境因子(如光照、昼夜节律、温度等)的共同影响。通常温度越低、光周期越短时棉铃虫性信息素组分的合成量就越高[35];如果用异常光周期或不同光质处理雌蛾,会导致性信息素释放量下降,求偶节律混乱等情况[36]。
在特定光暗周期为主的外部环境刺激下,PBAN由昆虫咽下神经节(subesophageal ganglion,SOG)中的部分神经细胞合成和分泌,再经由血淋巴输送到PG细胞。血淋巴中的PBAN与PG细胞膜上的G蛋白偶联受体(PBANR)结合,利用Ca2+和cAMP作为第二信使来完成性信息素生物合成的信号转导[37-43]。一方面,PBAN与PBANR结合后,激活细胞膜上的钙离子通道,促使细胞外的Ca2+向内流动。伴随着细胞内Ca2+浓度的升高,钙调蛋白(calmodulin,CaM)被激活并与钙调磷酸酶(calcineurin,CaN)相结合。CaN通过去磷酸化丝氨酸84和丝氨酸92位点来活化性信息素生物合成的限速酶——乙酰辅酶A羧化酶(acetyl coenzyme A carboxylase,ACC),该酶催化乙酰辅酶A(acetyl coenzyme A,acetyl CoA)生成丙二酰辅酶A(malonyl coenzyme A,malonyl CoA),并在脂肪酸合成酶(fatty acid synthases)的作用下生成长链脂肪酸,即性信息素特异性组分的前体物质[42, 44-45]。PBAN/PBANR/Ca2+/CaN信号通路在蛾类I型性信息素的生物合成中十分保守,在所有已检测的蛾类昆虫中均起着主导作用[37-43, 46-49]。另一方面,在以棉铃虫为代表的醛类性信息素合成时,PBAN能同时刺激腺苷酸环化酶(adenylate cyclase,AC)的活性,促进环腺苷酸(cyclic adenosine monophosphate,cAMP)的水平升高。cAMP作为第二信使激活下游的蛋白激酶A(protein kinase A,PKA),从而降低AMP激活蛋白激酶(AMP-activated protein kinase,AMPK)的活性。AMPK是ACC的负调节因子,其活性被抑制能保证CaN对ACC的去磷酸化激活活性,最终使性信息素生物合成得以顺利进行[42]。
在上述模型中,PBAN同时利用Ca2+和cAMP为作为第二信使;PBAN/PBANR/Ca2+/CaN信号通路与PBAN/PBANR/cAMP/PKA信号通路的共同作用保证了棉铃虫醛类性信息素的顺利合成。以家蚕为代表的醇类性信息素的生物合成仅依赖PBAN/PBANR/Ca2+/CaN一条信号转导途径,与棉铃虫明显不同[34, 50-51]。酯类性信息素的生物合成研究较少,目前仅报道了亚洲玉米螟Ostriniafurnacalis性信息素的PBAN信号转导机制,当主要成分顺-12-十六碳烯酯被合成时,PBAN仅招募Ca2+为第二信使,cAMP的胞内水平未见明显变化[49]。
3 棉铃虫性信息素特异性组分的合成
蛾类依赖其性信息素在结构方面的多样性(如链长、含氧官能团类型、双键的数量和位置等)以及各组分的精确释放比例来促进种内两性通讯,并维持种间生殖隔离[20]。蛾类性信息素往往由两种或两种以上的化学物质组成,其中一种化学物质含量较高,为主要成分,如家蚕性信息素主要由E8,Z10-16∶OH和反10,顺12-十六碳二烯醛(trans-10,cis-12-hexadecadienal,E10,Z12-16∶Ald)两种组分构成,其中E8,Z10-16∶OH的含量高达92%[52]。有趣的是,烟芽夜蛾Heliothisvirescens、棉铃虫、烟青虫Helicoverpaassulta和H.subflexa等多种夜蛾科昆虫都是利用特定混合比例的Z9-16∶Ald和Z11-16∶Ald作为自身的性信息素成分;甚至在人工干预下,棉铃虫和烟青虫可以杂交并产生不育后代[53]。因此可以认为,性信息素各组分之间的合成滴度与释放比例控制是昆虫准确识别同种异性并维持种间隔离的关键。
3.1 棉铃虫性信息素特异性组分的合成
大多数雌蛾释放的性信息素组分是末端具有不同官能团的不饱和脂肪链,前体物质经过去饱和、碳链缩短、官能团生成等碳链修饰反应,最终生成具有生物活性的信息化合物[54]。去饱和酶(desaturase,△)、脂肪酰基还原酶(fatty acyl-CoA reductase,FAR)、醇氧化酶(alcohol oxidase),以及β-氧化途径相关酶(β-oxidation related-enzyme)被认为是参与前体脂肪链合成后步骤的主要功能酶系,负责特异性性信息素组分的生成[42, 55]。当前对特异性组分合成调控的研究主要集中在对去饱和酶和FAR的功能鉴定上。其中,去饱和酶主要负责性信息素前体中双键或三键的形成;通过在脂肪链中的特定位置引入氧原子,后经脱氢反应生成H2O,形成顺式或反式的不饱和结构[56]。FAR主要负责将性信息素的脂肪链前体末端的脂酰辅酶A催化生成相应的醇类,后经氧化作用或乙酰化反应生成相应的醛类或酯类化合物[57]。
在棉铃虫中,经过PBAN的激活作用和级联反应生成了性信息素前体,即十六碳酰基辅酶A(16∶CoA)和少量十八碳酰基辅酶A(18∶CoA)。研究发现,棉铃虫PG细胞的转录组中同时包含△11和△9这两类去饱和酶基因,且△11基因的表达水平显著高于△9,这与棉铃虫性信息素混合物中Z11-16∶Ald与Z9-16∶Ald的比例接近。有学者因此推测△11主要通过作用在16∶CoA而形成Z11-16∶CoA(Z11-16∶Ald的合成前体),而△9则利用16∶CoA为前体生成Z9-16∶CoA(Z9-16∶Ald的合成前体)[58-60]。这一推测在烟青虫的研究中得到了验证,但是棉铃虫中的情况似乎并不支持这一推论。甚至有学者指出,在棉铃虫PG中未发现△9的表达(这可能是受当时的测序条件限制,△9的表达丰度过低而未能检测到)。关于棉铃虫性信息素主要组分Z11-16∶Ald的形成,学者们的意见比较一致,都认为是16∶CoA在△11的氧化作用下,首先形成不饱和的Z11-16∶CoA,再在FAR的催化作用下生成Z11-16∶OH,后经醇氧化酶的作用最终生成Z11-16∶Ald[54, 58-62]。在合成次要组分Z9-16∶Ald时,比较认可的途径是18∶CoA作为前体物质经过△11的氧化作用先生成Z11-18∶CoA,再经过β-脂肪氧化途径去除两个碳原子后形成Z9-16∶CoA。Z9-16∶CoA再经过酰基还原作用和醇氧化作用生成少量的Z9-16∶Ald。美洲棉铃虫与棉铃虫具有相似的性信息素组成,这一推论也在对美洲棉铃虫的研究中得到了验证[59, 62-63](图1)。
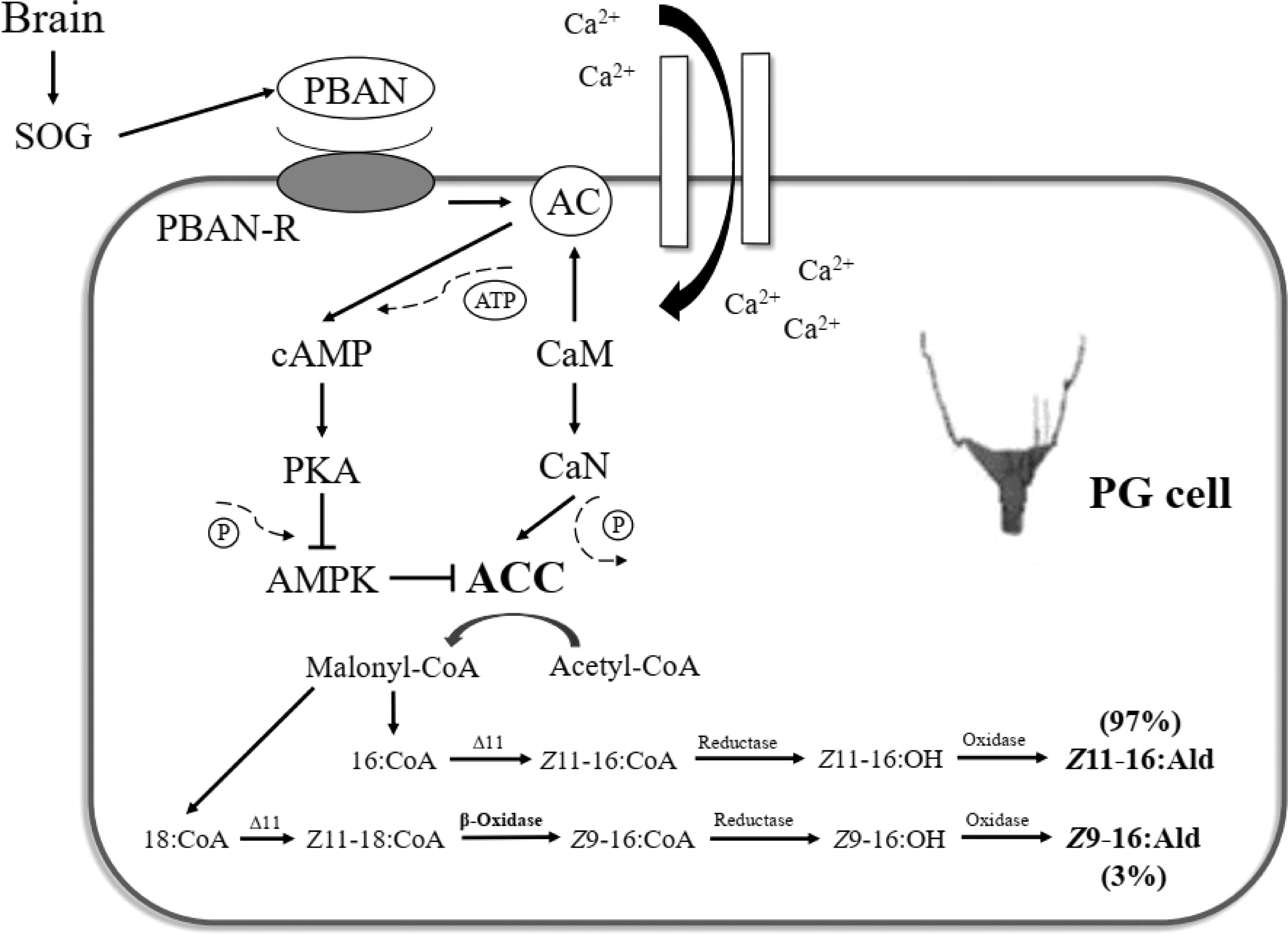
图1 棉铃虫性信息素生物合成模式图
3.2 棉铃虫性信息素特异性组分的比例控制
性信息素特异性组分的合成及滴度控制主要依赖脂肪链前体合成后的碳链修饰过程,其中,FAR和去饱和酶基因的适量表达可能是维持性信息素各组分适宜释放比例的重要因素。对不同亚型的欧洲玉米螟Ostrinianubilalis性信息素生物合成进行研究时发现,长链脂肪酸前体的总量对各组分最终合成滴度的影响很小,而FAR是调节性信息素各组分释放比例的关键因子[64]。也有学者提出,棉铃虫中特异性的去饱和酶可能是产生不同性信息素组分的关键因素[58-59]。但如前文所述,棉铃虫两种主要组分的生物合成均依赖于△11,这与该结论似乎矛盾。Hagström等构建酵母表达体系对以烟芽夜蛾为主的4种蛾类(包括棉铃虫)的性信息素合成进行了研究,指出FAR(而非去饱和酶)对底物的半选择性(semi-selective)是维持各组分特定合成比例的一个重要因素[63]。结合特异性组分的合成机制,推测FAR的底物偏好性可能是控制棉铃虫Z11-16∶Ald和Z9-16∶Ald合成比例的关键因素,但具体的调控机制还有待进一步的明确。
4 棉铃虫性信息素生物合成的终止调控
由PBAN介导的棉铃虫等蛾类昆虫的性信息素生物合成的激活机制已经被较好地阐明,但关于性信息素合成的终止调控研究非常有限。性肽(sex peptide)是一种产生于雄性附腺的小分子肽,能通过交配传递给雌性昆虫,用于抑制其性信息素的合成和释放[65-67]。将纯化后的黑腹果蝇Drosophilamelanogaster的性肽注射到雌虫体内,可以降低雌虫对雄虫的敏感性,使雌虫拒绝交配,并刺激其产卵。研究还发现,性肽能够同时刺激保幼激素(juvenile hormone,JH)含量升高[65-66]。有趣的是,性肽能够跨物种发挥作用,将纯化后的果蝇性肽注射到棉铃虫体内,发现了与果蝇相似的抑制作用,以及JH滴度的显著上升[68]。
JH可能是抑制性信息素生物合成的另一个关键因子。用JH处理新羽化的棉铃虫未交配雌蛾,PBANR的表达水平显著降低,进而抑制了性信息素的合成和释放。而交配所导致的棉铃虫雌蛾JH的水平上升,也关联了其性信息素释放量的减少[69-70]。这似乎都说明JH可能是降低性信息素生物合成的一个重要因素。但是也有研究发现,用JH处理羽化前1 d的雌蛹,能诱导PG提前响应PBAN信号,启动性信息素的合成[70]。关于JH在蛾类性信息素的生物合成中的具体功能还存在争议,还需要更深入的研究。
Yang等[71]最新的研究揭示了章鱼胺(octopamine)终止棉铃虫性信息素生物合成的新机制。他们发现,棉铃虫性信息素释放水平与体内章鱼胺滴度呈负相关,用章鱼胺处理后,雌蛾性信息素的合成被显著抑制。进一步发现章鱼胺利用OctβR为功能受体,通过抑制细胞内Ca2+水平的上升和下游CaN/ACC的活性来阻止PBAN信号的转导,最终终止棉铃虫性信息素的生物合成[71](图2)。

图2 章鱼胺终止棉铃虫性信息素生物合成的调控机制(仿Yang等[71])
5 总结与展望
鳞翅目是昆虫纲的第二大目,包含20多万个已知物种,其中90%为蛾类昆虫。尽管不同种类的飞蛾在形态和生物学上存在差异,但是由于蛾类昆虫几乎不依赖视觉功能,大多数蛾类在两性交配时以性信息素作为种内沟通的信号物质,这类具有种间特异性的化合物是保证种群有序繁衍的重要因子。了解性信息素的生物合成及调控机制,对于了解昆虫化学通讯的奥秘、揭示害虫暴发的机理以及指导田间应用等都具有重要的意义。
本文以棉铃虫为例,总结了蛾类昆虫中醛类性信息素生物合成和调控的一般机制,这对于研究其他蛾类性信息素的生物合成机制具有借鉴和比较意义。尽管棉铃虫性信息素的研究相比其他昆虫较为完善,但仍有许多盲点有待进一步阐明。这包括:1)从棉铃虫PG中鉴定出的多种次要组分的功能还有待进一步研究,尤其是这些化合物在两性成虫近距离接触时是否具有行为调控功能,以及发挥了怎样的作用,都还有待发掘和阐明;2)尽管不少研究已经揭示了去饱和酶和FAR能够控制棉铃虫等蛾类性信息素各组分的种类和滴度,但调控机制尚不明确;3)性信息素释放的终止调控是环境、行为、激素等多重因素的综合作用,当前对终止机制的内在分子机理研究较少,关键基因的发掘和功能鉴定,以及磷酸化蛋白组学的调控机制等都不清楚;4)以棉铃虫为代表的醛类性信息素与酯类或醇类性信息素在生物合成和调控方面的异同点,也是今后研究的重点方向。此外,如何将当前和今后的理论研究成果融入到害虫绿色防控和综合治理体系,也是学者们需要考虑的问题,这提示我们今后应该更加注重以应用为导向的机理研究。
