接种外生菌根真菌对不同种源栓皮栎幼苗生长和生理特征的影响
2023-01-12高晓磊张丽仔朱景乐
高晓磊,张丽仔,唐 窈,邹 锋,熊 欢,朱景乐,唐 骏
(1.中南林业科技大学 经济林培育与保护教育部重点实验室,湖南 长沙 410004;2.中国林业科学研究院经济林研究所,河南 郑州,450003;3.国有信阳市平桥区天目山林场,河南 信阳 464194)
栓皮栎Quercus variabilis是壳斗科Fagaceae栎属Quercus落叶乔木,是重要的珍贵用材树种,是我国软木、栲胶的重要来源[1]。栓皮栎在我国分布广泛,北起辽宁省南至云南省均有分布,在涵养水源、保持水土和维持生态平衡方面发挥着重要作用[2]。然而由于长期的乱砍滥伐和生境破坏,栓皮栎林的数量和质量不断下降,严重影响到我国软木、栲胶等产业的发展,经济和生态效益受损严重[3]。栓皮栎育苗方式为圃地播种,移栽造林过程中起苗易伤根,在丘陵山地造林实践中,存在苗木侧根稀少造成的苗木成活率低、生长不良等问题,严重阻碍了栓皮栎的造林进程。栓皮栎是外生菌根(Ectomycorrhiza,ECM)宿主植物,目前菌根化育苗技术被认为是提高苗木造林成活率的有效措施,在苗木培育、植树造林等方面具有广阔的应用前景。
植物激素调控着根系发育,其中生长素起主导作用[4],生长素等内源激素在根尖的积累和梯度分布诱导了宿主植物菌根的发育,从而改变了宿主植物的根系构型[5]。在经过接菌处理的福鼎大白茶Camellia sinensis ‘Fudingdabaicha’、苹果Malus pumila、番茄Lycopersicon esculentum等植株根系内均出现了IAA 和GA3等激素含量的增加以及根系的扩张[6-8]。接菌增强了植株对水分及养分的吸收能力,进而对宿主植物的生长有着良好的促进作用[9,10],Segnitz 等[11]也报道了ECM 对植物根系吸收土壤养分及生长的积极影响。Makita 等[12]对日本枹栎Q.serrata进行接菌处理,研究结果显示,ECM 可以通过调节植物对水分和养分的吸收来改变植物的形态特征,使枹栎的比根长增大,扩大根系的吸收范围。
光合作用是绿色植物重要的生理过程,植物的光合特性一定程度上能够反映植物对环境的适应情况[13],植物叶片发出的叶绿素荧光信号反映了植株的光合作用信息,可以利用叶绿素荧光参数研究植株光合作用的真实状况[14]。在对接菌植物光合特性的研究中发现,ECM 真菌接种能够显著提高叶片的净光合速率和水分利用效率,Martins 等[15]报道称菌根真菌促进了欧洲栗Castanea sativa叶片的净光合速率提升;高悦等[16]也对华美牛肝菌Boletus speciosus、彩色豆马勃Pisolithus tinctorius等外生菌根真菌接种的马尾松Pinus massoniana、湿地松P.elliottii和黑松P.thunbergii幼苗的叶绿素含量及其主要叶绿素荧光参数(ФPS Ⅱ和ETR)进行了研究。目前菌根化育苗技术在世界已有广泛应用,在栓皮栎领域,刘现刚等[17]研究了接种绵毛丝膜菌Cortinarius sublanatus和美味牛肝菌B.edulis对北京种源栓皮栎幼苗光合生理和气体交换特征的影响,而关于外生菌根对不同种源栓皮栎生长及生理影响的研究尚少见报道。
硬皮马勃Sclerodermasp.是一类常见的菌根真菌,与壳斗科植物普遍共生。本研究以北京和河南种源的栓皮栎为试材开展硬皮马勃接种盆栽试验,比较分析了不同处理栓皮栎幼苗生长、叶绿素荧光参数、光合参数、根尖内源激素含量的差异,旨在阐明ECM 真菌对不同种源栓皮栎幼苗生长和生理特征的影响,以期为探索主要造林树种的菌根应用技术提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于湖南省长沙市中南林业科技大学苗圃(112°59′32″E,28°08′12″N)。试验地海拔92 m,年平均气温17.2℃,年降水量为1 200~1 400 mm,属亚热带季风性湿润气候,气候温和,降水充沛,雨热同期,该地区母岩为典型的第四纪网纹红壤,风化程度较深,区内土壤为低酸性土壤或中性土壤,适宜植物及菌根真菌生长,地带性植被为常绿阔叶林。
1.2 试验材料制备
1.2.1 菌剂制备
从湖南省浏阳市板栗人工林采集子实体后,进行分离纯化,经分子鉴定为硬皮马勃,简称Sc[18]。于MMN 培养基进行平板培养,然后用不含琼脂的MMN 培养基做液体培养,最后在草炭和蛭石2∶1 混合的基质上扩大培养,得到固体菌剂[19]。
1.2.2 播种
于2019 年3 月中旬进行播种,将黄心土∶泥炭按照体积分数2∶1 混合作为容器育苗基质,经过37%的甲醛消毒处理,通风阴干后分装入口径20 cm,高22 cm 的塑料容器中,将北京(BJ)和河南(HN)种源栓皮栎种子经0.5%的KMnO4溶液浸泡2 h,用清水洗净阴干后进行播种。BJ 和HN 种源各播种150 盆,每盆播种一粒种子,2 周后,发芽率分别为89.33%和92.67%,选取出苗时期、苗高、地径相对一致的幼苗作为试验材料。
1.2.3 接种
本试验采取随机区组试验设计,共设置北京种源-接菌(BJ-Sc)、河南种源-接菌(HN-Sc)、北京种源-不接菌(BJ-CK)、河南种源-不接菌(HN-CK)等4 个处理,3 次重复,每个重复20 株,共240 株。待栓皮栎苗长出2~3 片真叶,在幼苗周围挖5 cm 深的孔洞,将硬皮马勃固体菌剂施入其中并覆土,每株幼苗接种5 g 固体菌剂,对照施入不含菌剂的基质(CK)。各处理摆放间距0.5 m以上,以防止相互侵染。育苗期间正常浇水管理。
1.3 测试指标及方法
1.3.1 苗高、地径测量
5 月7 日至7 月7 日(每隔10 d)、9 月5 日、11 月14 日观测栓皮栎幼苗的生长情况,使用卷尺测量苗高(cm),使用游标卡尺测量地径(mm)。
1.3.2 外生菌根形态观察
每个处理随机抽取3 株幼苗,取其根系浸泡在水中,洗去浮土,剪成5 cm 长的根段,将根段放入含有少量水的表面皿中,置于OLYMPUS SZX16 体视显微镜下观察其菌根形态[20]。
1.3.3 石蜡切片
将菌根根尖在Carnoy 固定液中抽气10 h,完全排除材料中的空气,置于70%的酒精溶液中,4℃下储存。对固定的材料进行适当修整,并使用6 种浓度酒精进行脱水,用二甲苯使样品透明,嵌入石蜡中,然后用石蜡切片机(徕卡RM2235,德国海德堡)切片,厚度为10 µm。用1%番红染色,制作封片,并使用显微镜(德国海德堡徕卡DM2500)获取照片[18]。
1.3.4 细根生长指标测定
将上述根系经扫描仪EPSON V7000 扫描成图片,利用根系分析软件分析直径为0~2 mm 的细根根长、根表面积、根体积等指标[21]。
1.3.5 叶片SPAD 值测定
SPAD 值代表叶绿素相对含量,其与叶绿素含量呈显著正相关,能够有效反映叶绿素的水平。每个处理随机选取3 株幼苗,使用便携式SPAD-502 Plus 叶绿素仪测定叶绿素含量,每株幼苗选择同一叶位、无病虫害的成熟叶片,测定3次,记录数据[22]。
1.3.6 叶绿素荧光参数的测定
于8 月下旬晴朗天气的上午,每个处理随机选取3 株幼苗,选择生长一致且受光方向相同的叶片,采用英国Hansatech 公司生产的FMS2 型脉冲调制荧光仪,参照冯玉龙等[23]的方法,测定光系统Ⅱ(PS Ⅱ)最大光能转换效率(Fv/Fm)、PS Ⅱ实际光化学效率(ФPS Ⅱ)及电子传递速率(ETR)。
1.3.7 光合参数的测定
对于上述幼苗,于晴朗天气上午10:00,选择同一叶位叶片(从顶端数第5 片功能叶),使用Li-6400 便携式光合仪测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)及蒸腾速率(Tr),并计算水分利用率(WUE):水分利用率(WUE)=净光合速率(Pn)/蒸腾速率(Tr)[17]。
1.3.8 根系内源激素含量测定
取上述幼苗的根尖,采用酶联免疫吸附测定法(ELISA)试剂盒测定玉米素核苷(ZR)、生长素(IAA)、赤霉素(GA3)和脱落酸(ABA)含量[24],每个样本重复测定3 次。
1.4 统计分析
使用Excel 2018 软件进行数据处理,用SPSS 13.0 软件实现方差分析及LSD 多重比较,显著性水平设为P<0.05。
2 结果与分析
2.1 接种硬皮马勃的菌根形成情况
在体视显微镜下可见接种Sc 的栓皮栎幼苗根系出现黄白色短棒状菌根,白色絮状菌丝(图1A—B)。对菌根横截面进行切片观察,可见与正常根相比,菌根的表皮细胞外覆盖有一层致密的菌套(Mantle),哈氏网(Hartig net)延伸至表皮和皮层细胞间(图1C—D)。硬皮马勃与栓皮栎的亲和性较强,接菌后60 d,BJ 和HN 幼苗的菌根侵染率分别为43.33%和45.00%;接菌后90 d,达到76.67%和81.33%;接菌120 d 时,两个种源幼苗的菌根侵染率均超过90%,CK 组菌根侵染率为0(图1E)。

图1 栓皮栎幼苗菌根形态特征及侵染率Fig.1 Morphological characteristics and colonization rate of mycorrhiza in Q.variabilis seedlings
2.2 接种硬皮马勃对栓皮栎幼苗生长动态的影响
接种硬皮马勃处理的栓皮栎幼苗生长表现明显优于未接菌幼苗。如图2 所示,5 月8 日—6 月27 日(接菌后10~60 d),幼苗生长速率较高;6 月27 日接菌处理与对照间幼苗高和地径出现显著差异(P<0.05);6 月27 日—7 月7 日(接菌后60~70 d),菌根苗仍维持较高生长速率,对照的幼苗生长速率明显减缓;11 月4 日(接菌后190 d),接菌的BJ、HN 栓皮栎苗高分别为37.02 和37.69 cm,与相应对照相比,增幅分别达到19.5%和18.9%,2 个种源栓皮栎菌根苗地径分别为4.55 和4.60 mm,较相应对照高14.9%和14.4%,表明接种硬皮马勃能够显著促进栓皮栎幼苗的生长。
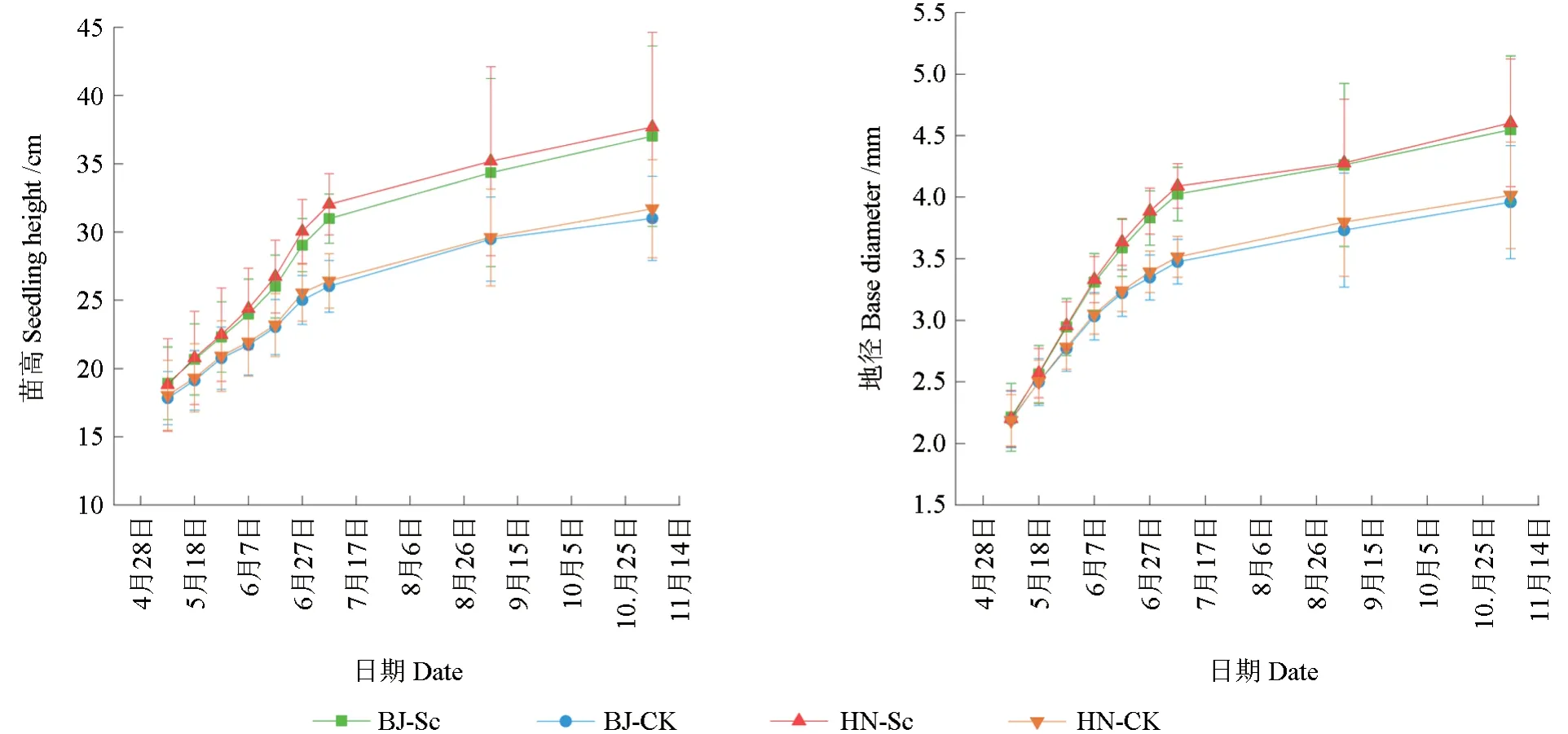
图2 不同处理的栓皮栎幼苗生长动态Fig.2 Growth dynamics of the Q.variabilis seedlings under different treatments
2.3 接种硬皮马勃对栓皮栎幼苗根系形态的影响
由表1 可见,接种硬皮马勃的BJ、HN 栓皮栎幼苗细根根长分别比相应对照高166.9%和97.6%,根表面积高163.4%和83.5%,根体积高127.7%和64.6%,均达到显著水平(P<0.05),表明接种硬皮马勃能够显著促进栓皮栎幼苗细根的生长;两个种源间未接菌幼苗的细根根长、根表面积及根体积无显著差异,BJ 菌根苗的细根根长、根表面积及根体积显著高于HN 菌根苗。
2.4 接种硬皮马勃对栓皮栎幼苗叶片叶绿素荧光参数和SPAD 值的影响
接种硬皮马勃显著提高了HN 栓皮栎叶片的实际光化学效率(ФPS Ⅱ)和PS Ⅱ电子传递速率(ETR),对光能转换效率(Fv/Fm)的影响不显著。与对照相比,ФPS Ⅱ和ETR 分别提高了53.6%和62.3%。然而,接种硬皮马勃对BJ 栓皮栎幼苗的叶片叶绿素荧光参数无显著影响。接种硬皮马勃显著提高了BJ 和HN 栓皮栎幼苗叶片SPAD 值,增幅分别为13.9%和15.4%。BJ 和HN 间未接菌栓皮栎幼苗叶片的叶绿素荧光参数及SPAD 值均无显著差异。
2.5 接种硬皮马勃对栓皮栎幼苗光合参数的影响
接种硬皮马勃显著提高了BJ 和HN 栓皮栎幼苗叶片的净光合速率(Pn)(表3)。两个种源的净光合速率分别比相应对照高40.2%和74.5%,Pn大小顺序为HN-Sc >BJ-Sc >BJ-CK >HN-CK。HN 菌根苗叶片气孔导度(Gs)及蒸腾速率(Tr)显著高于CK。接菌处理对两个种源幼苗叶片的胞间CO2浓度(Ci)及水分利用效率(WUE)均无显著影响。两个种源间菌根苗叶片的Pn、Gs、Ci及WUE 无显著差异,其未接菌栓皮栎幼苗叶片的Pn、Gs、Ci亦无显著差异,HN 的未接菌栓皮栎幼苗叶片的Tr值显著低于BJ 种源。
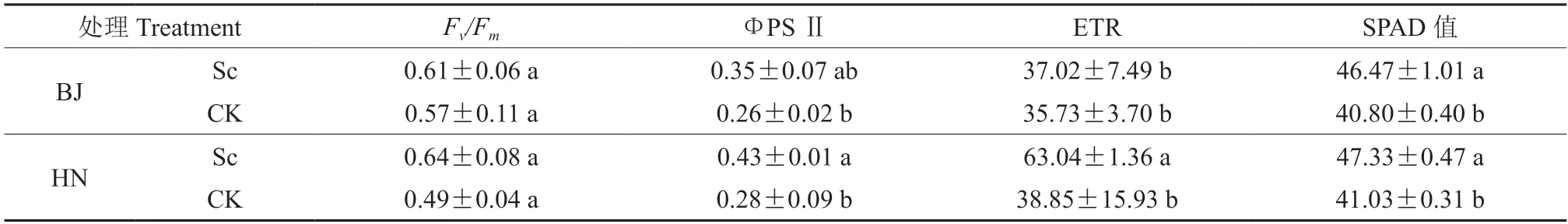
表2 不同处理的叶片叶绿素荧光参数及SPAD 值比较Table 2 Comparison of the chlorophyll fluorescence parameters and SPAD values of leaves under different treatments
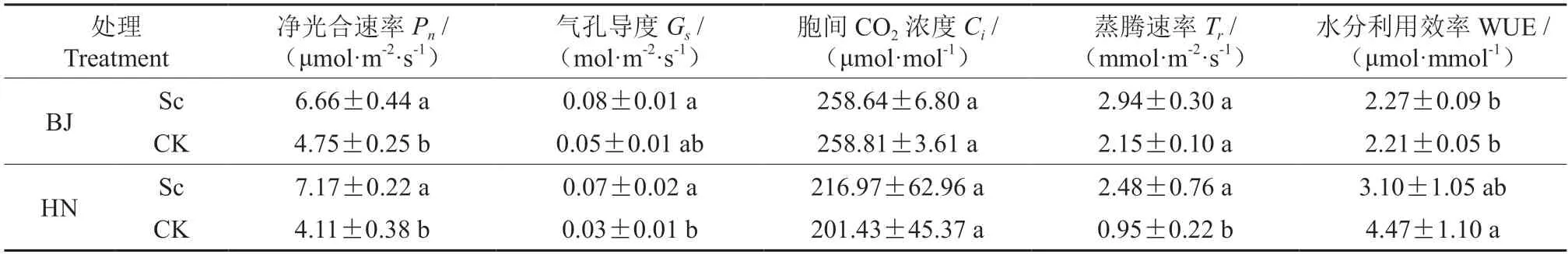
表3 不同处理的栓皮栎叶片光合参数比较Table 3 Comparison of the photosynthetic parameters of the Q.variabilis leaves under different treatments
2.6 接种硬皮马勃对栓皮栎根尖内源激素含量影响
BJ-Sc 的根尖IAA 含量为99.8 ng·g-1,较CK显著提升12.1%,ABA 含量为96.64 ng·g-1,较CK显著增加12.2%。HN-Sc 的根尖IAA 和ABA 的含量也显著高于相应的未接菌苗,增幅分别为15.3%和17.4%(P<0.05)(表4)。接种硬皮马勃对两个种源栓皮栎幼苗根尖的GA3及ZR 含量均无显著影响,HN-Sc 的IAA 和ZR 含量显著高于BJSc。两个种源间未接菌幼苗根尖的各激素含量无显著差异。
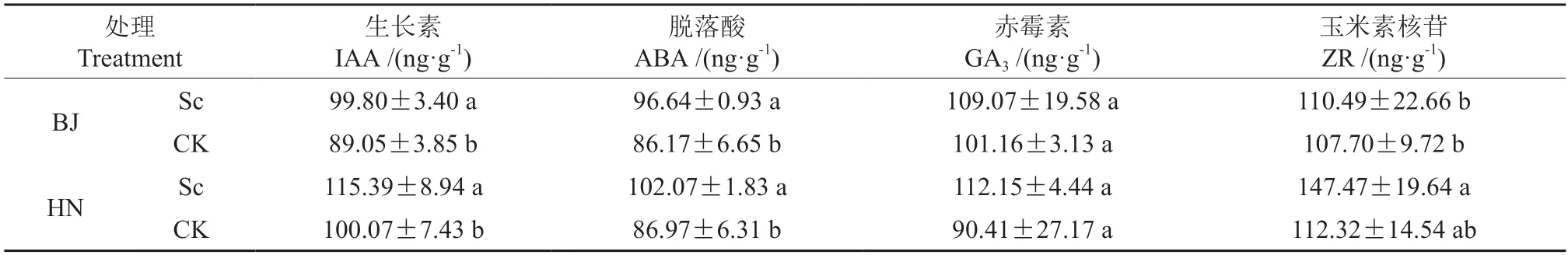
表4 不同处理的栓皮栎根尖内源激素含量比较Table 4 Comparison of the endogenous hormone content of Q.variabilis root tips under different treatments
3 结论与讨论
3.1 讨 论
植物内源激素在调节根系生理活动方面起着非常重要的作用,报道显示,外生菌根真菌能够诱导植物根尖激素的积累进而促进细根的形成[25]。本研究中,Sc 接种处理的BJ 及HN 栓皮栎幼苗根系内IAA 及ABA 含量显著高于未接种处理。细根是植物吸收水分和养分的主要器官[22],试验中接种Sc 能够增加栓皮栎幼苗的细根长度、表面积和体积,扩大根系的吸收范围,且对北京种源栓皮栎幼苗根系的促生效果更好。祁金玉等[26]亦研究证明,接种外生菌根真菌褐环乳牛肝菌(Suillus luteus)能促进油松(P.tabuliformis)根系构型扩张。说明ECM 真菌侵染宿主植物根系后,能够影响宿主植物内源激素含量并改变其平衡状况,促进细根生长,进而能够更好地适应环境,增强抗逆能力。
更进一步而言,根系的扩张促进了植株对土壤水分、养分及矿质元素的吸收,进而影响叶片光合作用[27]。植物叶片的SPAD 值与叶绿素a、叶绿素b 和总叶绿素含量间均存在极显著正相关关系[28],本研究中,菌根苗叶片SPAD 值较对照高14%以上,反映了接菌能够增加栓皮栎叶片的叶绿素含量,这一结论与徐洪文等[29]证实接种菌根真菌能够显著提高玉米(Zea mays)叶片SPAD 值的结果一致,这可能是由于接菌促进了根系对N、Mg 等营养元素的吸收[30],为叶绿素的合成提供了物质基础。此外,硬皮马勃的接种显著提高了HN栓皮栎的实际光化学效率和PS Ⅱ电子传递速率,显著提升了BJ 和HN 栓皮栎叶片的净光合速率,HN-Sc 处理的Pn值最高,达到7.17 μmol·m-2·s-1,气孔导度也显著大于对照,而细胞间CO2浓度无显著差异,说明接菌导致净光合速率的提升是由非气孔因素引起的。综合SPAD、ФPS Ⅱ、ETR、Pn的变化,可以认为,接种Sc 能够提升叶绿素含量,光合色素吸收光能的增加进而促进了光合电子的传递,增强了PS Ⅱ反应中心光化学活性,提高了叶片净光合速率。
植株根系吸收能力及叶片光合速率的增强,共同促进了植株地上部分的生长[31]。栓皮栎接种Sc 后,苗高、地径得到了显著提高,接种后190d,接菌的BJ 和HN 栓皮栎苗高分别较对照提高19.5%和18.9%,地径分别较对照提高14.9%和14.4%,表明ECM 真菌能极大促进幼苗的生长,与阎秀峰等[32]发现接种铆钉菇(Gomphidius viscidus)和臭红菇(Russula foetens)能够促进辽东栎(Q.wutaishanica)生长的结果相似。7 月,接菌处理和对照的栓皮栎幼苗生长速率均有不同程度的减缓(图2),可能是由于夏季高温,栓皮栎幼苗正值种子供给营养与叶片光合供给营养的转换期,在夏季生长缓慢[33]。BJ 和HN 栓皮栎未接菌幼苗的蒸腾速率(Tr)及水分利用效率(WUE)出现显著差异,且Sc 对两个种源栓皮栎幼苗的根系和光合作用促进程度不同,可能是由于不同种源栓皮栎在不同生境下的适应性及对硬皮马勃的亲和性不同。
3.2 结 论
综上研究结果,接种硬皮马勃能够显著增加北京及河南种源栓皮栎幼苗根尖内IAA 及ABA 含量,促进细根生长,并进一步提高栓皮栎叶片的SPAD 值,改善叶片光能利用效率,显著提高幼苗叶片净光合速率,促进栓皮栎幼苗的生长。接种硬皮马勃对栓皮栎生长促进作用显著,且对北京栓皮栎的根系促生作用优于河南,对河南栓皮栎的光合作用促进效果优于北京。
