光照与氮磷营养盐的协同作用对海带幼苗释放可溶性有机碳的影响
2022-12-21尼志杰张永生褚洪永张明亮孙琰晴胡顺鑫
尼志杰 张永生 褚洪永 李 斌 张明亮 孙琰晴 胡顺鑫
(1.上海海洋大学水产与生命学院, 上海 201306; 2.山东省海洋资源与环境研究院, 烟台 264006; 3.荣成市海洋与渔业执法大队, 威海 264300; 4.烟台市海洋经济研究院, 烟台 264003)
海带(Saccharina japonicaJ.E.Areschoug)是褐藻纲海带属的一种藻类, 原产于东北亚地区(日本、朝鲜北部沿海和俄罗斯太平洋沿岸), 后来在我国辽东半岛和山东沿海地区广泛分布, 逐渐成为我国海水养殖规模最大的海藻物种。据世界粮农组织(FAO)的数据统计, 2017年我国贡献了全球海带产量的18%[1,2]。目前, 我国养殖海带的主要品种包括“大连无边”“东方5号”“爱伦湾”和“901海带”等[3—5]。
近海大型藻类养殖碳汇是渔业碳汇的重要组成部分[6], 大型藻类的规模化养殖促进了海洋对大气CO2的吸收[7], 这一观点已经在不同海藻种类中得到证实。狐尾藻(Myriophyllum spicatum)在杭州湾的固碳效率为1.25 mg/(g DW∙h)[8], 龙须菜(Gracilaria lemaneiformis)在盐田湾的固碳效率最高达到了9.25 mg/(g DW·h)[9]。据估计我国每年养殖藻类固碳量约达3.52 Tg/Ca[10], 其中海带对碳汇渔业的贡献约占总养殖海藻的73%[11]。大型藻类作为海洋生态系统的初级生产者, 能够通过光合作用将海水中的溶解无机碳(Dissolved Inorganic Carbon,DIC)转化为有机碳, 并显著降低海水二氧化碳(CO2)分压, 促进了海洋对大气CO2吸收[12]。不仅如此, 大型海藻还能够将20%—30%光合作用产物以可溶性有机碳(Dissolved Organic Carbon, DOC)的形态释放到海水中[13]。据估算, 我国海藻养殖每年释放DOC总量约为(82.2—91.5)×107kg, 海藻释放的DOC经海洋微型生物碳泵(Microbial Carbon Pump,MCP)作用, 每年可生成60×107kg以上的惰性溶解有机碳(Recalcitrant Dissolved Organic Carbon,RDOC), 该碳汇量约为我国海岸带“蓝碳”埋藏量的1.7倍[10,13]。由此可见, 海藻养殖在碳汇渔业中起到重要作用。
但关于光照与营养盐的协同作用对海藻释放DOC的影响机制仍存在争议。目前主要存在两种假说, 一种是“溢出”假说, Fogg[14]认为藻类释放DOC与光照强度呈正相关, 光强促进了溶解态大分子物质的释放。在Cherrier等[15]的室内和海区实验中,浮游藻类的DOC释放速率均与光照强度呈正相关;BarrÃ3n等[16]对多个关于底栖藻类的实验数据研究后, 同样发现底栖海藻DOC净通量随光强增加而增大, 这些报道都表明光照强度与DOC释放速率有显著的相关性。另一种是“扩散”假说, Bjornsen[17]认为藻类释放DOC的速率与光照强度无关, 与营养盐含量密切相关, 营养盐促进了藻类中溶解态小分子物质的释放。在Marañón等[18]的藻类培养实验中, 没有发现DOC释放与光照强度有相关性, 且在Mueller等[19]开展的光照与营养盐的交叉实验中, 珊瑚共生藻DOC释放与光照强度的相关性在营养加富后消失。以上研究表明, 光照与营养盐是调节藻类释放DOC的两种重要环境因素。基于此, 本研究以海带幼苗为研究材料, 基于光照和营养盐浓度两个因素设置室内交叉实验, 探究光照与营养盐的协同作用对大型海藻释放DOC的影响机制, 以期为海藻增养殖提供理论与技术支持。
1 材料与方法
1.1 试验材料
2020年12月从山东省荣成市东楮岛附近的海带养殖区(E 122°56′, N 37°05′)采集鲜活海带样品,样品平均体长(34.26±8.99)cm, 平均湿重为(5.25±1.86)g。在实验开始前, 于室内循环水养殖系统中暂养5d, 暂养期间光合有效辐射(Photosynthetically Active Radiation, PAR)为(63±9)μmol photons/(m·s),水温为(13.5±0.5)℃。
1.2 试验方法
设置了自然海水+50%海面光强、自然海水+100%海面光强、营养盐加富海水+50%海面光强、营养盐加富海水+100%海面光强4个实验组,每组设置5个平行样。光照强度的设置参照曹昀等[20]和孙百晔等[21]的研究方法, 其中100% PAR和50%PAR分别为(102±5)和(51±5)μmol photons/(m·s)。实验使用的自然海水中的磷酸盐、无机氮浓度分别为4.2和101 μg/L, 氮磷加富海水是在自然海水的基础上, 使用磷酸氢二钾、硝酸钾分析纯试剂分别配制成的100和1000 μg/L标准液进行加富, 使海水中营养盐的浓度达到藻类生长饱和浓度[22], 加富后各组氮、磷营养盐摩尔比分别为50%光强+自然海水(0.97﹕0.03)、100%光强+自然海水(0.92﹕0.06)、50%光强+氮磷加富(11.35﹕0.31)和100%光强+氮磷加富(11.42﹕0.81)。实验过程为从暂养的海带幼苗中挑选健康且规格相近的单株海带幼苗放入2 L经过酸洗的玻璃瓶中, 将海带幼苗按照上述4种不同的处理分别在光照培养箱中进行8h培养, 培养时水温保持在(14±0.5)℃。
1.3 海带幼苗释放DOC的速率检测
在培养开始和结束时, 分别取50 mL水样, 使用孔径0.45 μm滤膜进行抽滤, 抽滤后的水样于零下20℃冷藏柜(SC/SD-332)冷冻保存。使用岛津TOCLCPH总有机碳分析仪测定样品DOC含量, 具体操作参照Mueller[19,26]的方法。DOC释放速率[RDOC,μmol/(g·h)]指单位质量海带幼苗(干重)在单位时间内引起的水体DOC含量的变化,RDOC的计算公式为:
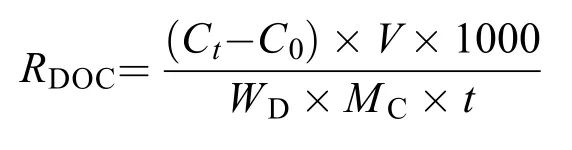
式中,Ct为实验组培养结束时的DOC浓度(mg/L),C0为对照组培养结束时的DOC浓度(mg/L),V为养殖用海水体积(L),WD为试验海带幼苗的干质量(kg),MC为碳的相对分子质量,t为试验处理时间(h)。
1.4 海带幼苗氧气释放速率检测
在培养开始前和结束时, 分别取100 mL水样,使用碘量法测定样品中溶解氧含量, 具体操作参照《海洋调查规范: 海水化学要素观测》(GB/T 12763.4-1991)。海带幼苗氧气释放速率[ΔCO2, μmol/(g·h)]是单位质量(干重)海带幼苗在单位时间内引起的水体溶解氧含量的变化。ΔCO2的计算公式为:
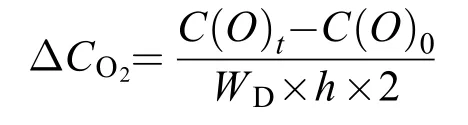
式中,C(O)t为试验结束时的氧气浓度[μmol/(g·h)],C(O)0为实验开始时的氧气浓度[μmol/(g·h)];V为养殖用海水体积(L);WD为试验海带幼苗的干质量(kg);t为试验处理时间(h)。
1.5 海带幼苗释放DOC占净初级生产力的比例检测
释放DOC占净初级生产力比重(Net Primary Productivity, NPP)比例(P, %), 是在假设碳固定与净产氧的摩尔比平衡(即1 mol碳固定等于1 mol氧气(O2)释放)条件下[19], 相同时间内海带幼苗释放DOC占净初级生产力比例。计算公式为:
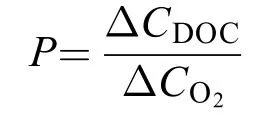
式中, ∆CDOC为单位时间内DOC浓度变化[μmol/(g·h)]; ΔCO2为单位时间内O2浓度变化[μmol/(g·h)]。
1.6 海带幼苗释放DOC的光谱斜率检测
在培养开始前和结束时, 分别取50 mL水样, 使用0.45 μm滤膜进行过滤, 抽滤后的水样于零下20℃冷藏柜(SC/SD-332)冷冻保存。使用UV-5100B紫外可见分光光度计测定水样的紫外可见吸收光谱, 具体操作参照陈昭宇等[24]的方法。吸收系数a(λ)的计算公式为:
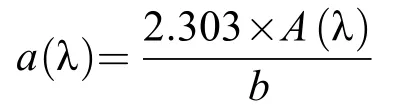
式中,A(λ)为吸光度,b为光程路径(m)。S275—295反映DOC相对分子质量与光反应活性, 相对分子质量越小值越大[25], 光谱斜率S的计算公式为:

式中,a(λ)是DOM吸收系数(/m ),λ是波长(nm),λ0是参照波长(nm)。
1.7 数据分析
实验数据通过Excel 2019进行整理, 使用SPSS 26.0进行单因素方差分析, 差异显著水平设置为P<0.05, 用LSD法进行多重比较, 图中不同字母表示差异水平为P<0.05, 相同字母表示差异水平为P>0.05, 使用OriginLab OriginPro 2021b SR1 v9.8.5.204进行作图。
2 结果
2.1 不同实验条件下海带幼苗的DOC释放速率
在自然海水条件下, 100%光强条件下海带幼苗DOC释放速率显著高于50%光强条件(P<0.05),表明此条件下光强与海带幼苗释放DOC速率呈正相关性。在加富海水条件下, 100%光强与50%光强释放DOC速率差异不显著(P>0.05), 表明此条件下光强与海带幼苗释放DOC无相关性。但在同等光照强度条件下, 加富海水组释放DOC速率显著高于自然海水组(P<0.05), 表明营养加富显著提升了海带幼苗DOC释放速率(图1)。
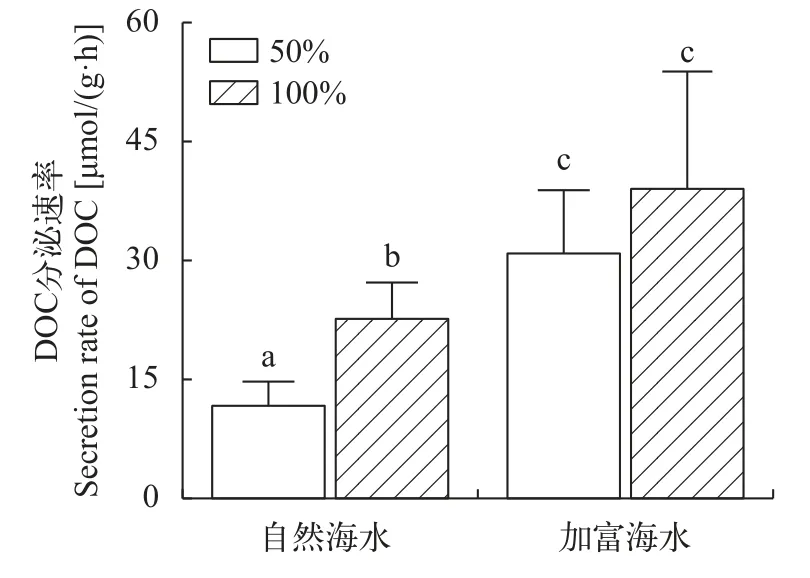
图1 不同实验条件下海带幼苗DOC释放速率Fig.1 DOC secretion rate of juveniles of S.japonica under different experimental conditions
2.2 不同实验条件下的海带幼苗释放DOC的光谱斜率
在自然海水条件下, 100%光强S275—295显著低于50%光强S275—295(P<0.05), 表明此条件下随着光强增加, 海带幼苗释放的DOC相对分子量增大(表1)。但在加富海水条件下, 100%光强S275—295与50%光强S275—295差异不显著(P>0.05), 表明此条件下光照强度并没有改变海带幼苗释放DOC的相对分子量。加富条件下的海带幼苗在100%光强时S275—295显著高于自然海水条件下100%光强S275—295(P<0.05), 表明在同等光照条件下, 营养盐加富显著降低了海带幼苗释放DOC的相对分子量(表1)。

表1 不同实验条件下海带幼苗释放DOC的S275—295值Tab.1 S275—295 values of DOC released from juveniles of S.japonica under different experimental conditions
2.3 不同实验条件下海带幼苗氧气释放速率
在自然海水、加富条件下, 100%光强组氧气释放速率均显著高于50%光强组(P<0.05), 表明海带幼苗不管处于何种营养水平, 光照强度提升均能显著促进氧气释放。但在同等光照水平下(100%光强、50%光强), 营养盐加富组氧气释放速率与自然海水组差异不显著(P>0.05), 表明海带幼苗不管处于何种光照水平, 营养盐加富未能显著促进氧气释放(图2)。
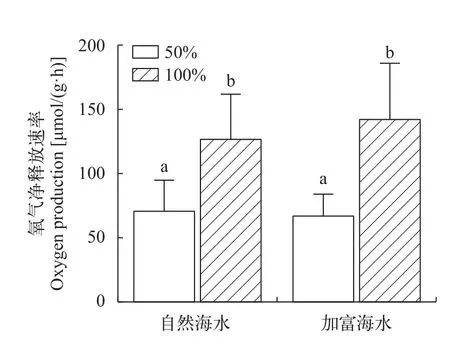
图2 不同实验条件下海带幼苗氧气释放速率Fig.2 Oxygen production of juveniles of S. japonica under different experimental conditions
2.4 不同实验条件下海带幼苗释放DOC占净初级生产力(NPP)比重
在自然海水条件下, 100%光强组与50%光强组释放DOC占NPP比例差异不显著(P>0.05), 表明在此条件下, 光照提升并未显著改变海带幼苗的DOC净释放率。在加富条件下, 50%光强组释放DOC占NPP比例显著高于100%光强组(P<0.05), 表明在此条件下, 光照强度提升反而降低了海带幼苗的DOC净释放率。在同等光照强度水平下(100%光强、50%光强), 加富组释放DOC占NPP比例均显著高于自然海水组(P<0.05), 表明通过营养盐加富促进了海带幼苗的DOC净释放率(图3)。
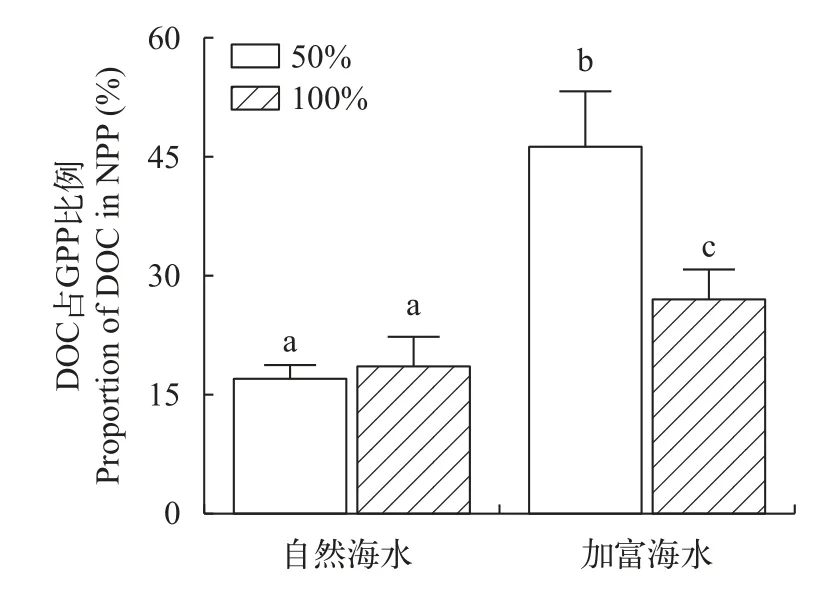
图3 不同实验条件下海带幼苗DOC释放量占NPP比例Fig.3 Proportion of DOC secretion in NPP of juveniles of S.japonica under different experimental conditions
3 讨论
尽管光照与营养盐的协同作用对藻类释放DOC的影响机制尚不明确, 但似乎“溢出”“扩散”两种假说均不足以解释其全部过程, 各有大量研究结果支持某一项“假说”也间接说明了这一点。如在Mueller等[19]的光照与营养盐交叉实验中发现, 在寡营养盐条件下, 珊瑚藻(Coralline algae)释放DOC与光强具有正相关性, “溢出”机制起作用; 但在添加氮磷营养盐后, 释放DOC与光强差异不显著, 表明在营养充足的条件下, “扩散”机制起作用。结合已有研究结果, 我们发现往往是在寡营养条件下,DOC释放与光照强度呈正相关, 表现为“溢出”机制。如前人对浮游藻类、珊瑚藻(Coralline algae)和巨藻(Macrocystis pyrifera)等藻类的研究发现, 当藻体处在营养盐浓度较低的自然海水中时, DOC释放随光强的增大而升高, 与“溢出”机制相符[15,26,27]。而在营养充足的条件时, “溢出”机制消失, 表现为“扩散”机制占主导。在Marañón[18]的研究中, 富营养条件下浮游藻类的DOC释放与光强没有显著相关性, 在Wyatt[22]对刚毛藻[Cladophora glomerata(L.)Kütz.]的研究中也有相同现象。为此有研究提出, 很有可能“溢出”“扩散”2种机制均存在, 只是在特定条件下某项机制占主导。在本研究中, 我们发现在自然海水(寡营养)条件下, 海带幼苗释放DOC与光照强度呈正相关, 且释放DOC相对分子量也随光强上升而提高, 表明在寡营养条件下, 海带幼苗释放DOC受“溢出”机制调节(图1)。在加富条件下, 海带幼苗释放DOC与光强无相关性, 表明“溢出”机制消失; 但在同等光照强度条件下, 加富组释放DOC速率显著高于自然海水组(图1), 不仅如此,加富还显著提升了DOC净释放率(图2), 且加富显著降低了释放DOC相对分子量(表1), 表明“扩散”机制占主导。
本研究推测海带幼苗释放DOC过程中“溢出”“扩散”机制均存在。在寡营养条件下, “溢出”机制占主导; 而营养盐水平一旦超过藻类生长饱和浓度, 此时“溢出”机制消失, “扩散”机制占主导。研究推测, 藻类光合作用受光照强度调节, 而细胞的生长则受无机营养盐限制。在N、P营养盐限制条件下, 随着光照强度升高, 藻类细胞光合作用产出将超过细胞生长对有机质的需求。此时细胞光合固定有机碳的速率将超过N、P供给速率, 从而导致细胞内C元素大量富集, 细胞内C﹕N﹕P比上升, 生成的有机质主要以高分子量物质为主。此时藻类表现为光强的增加促进有机质生成, 从而加剧DOC的释放, “溢出”机制占主导。当环境中N、P营养盐富集时, 此时藻类细胞内C﹕N﹕P比下降, 因此生成的有机质主要以低分子量物质为主。低分子量的DOC跨膜运输受细胞膜内外浓度差控制, 表现为“扩散”机制占主导。
本研究发现, 海带幼苗在100%光强+氮磷加富条件下DOC释放速率最高(图1)。为提升养殖海带碳汇效率, 可适当调整养殖模式, 通过同时提升海区营养盐及光照强度以促进养殖海带DOC的释放。海带养殖区往往营养盐含量较低, 呈现氮磷营养盐限制的状态, 为促进海带释放DOC, 可在海带养殖区进行N、P营养盐的缓释。虽然大型海藻对赤潮微藻生长具有一定的抑制作用[28,29], 但也应注意N、P营养盐的缓释可能引发微藻暴发性增殖造成的赤潮风险。在进行营养盐缓释的同时, 适当降低海带养殖密度,使海带叶片在单位面积上能够接收到更多光照。房景辉等[30,31]的研究表明, 降低养殖密度的标准化养殖同时也能提升养殖海带品质与产量。本研究可以为养殖海带增汇提供一定的理论依据与技术参考。
