黑腹果蝇fat2 新突变体的鉴定*
2022-12-16罗俊鹏田应雪张庆海
孙 蓉,杨 敏,罗俊鹏,向 阳,田应雪,梁 丹,魏 微,张庆海,4
(1.贵州医科大学 基础医学院 生物学系,贵州 贵阳 550025;2.贵州医科大学 基础医学科学研究中心,贵州 贵阳 550025;3.贵州医科大学 生物与工程学院,贵州 贵阳 550025;4.贵州医科大学 医学昆虫重点实验室,贵州 贵阳 550025)
黑腹果蝇卵巢因其结构简单使其成为研究生殖细胞发育的优良模型[1]。卵巢的基本单位是卵巢管[1],雌性果蝇含有1 对卵巢,每1 个卵巢由16~20 个卵巢管组成[2-3],每1 个卵巢管里含有处于不同发育阶段的卵子。当干细胞不对称分裂时,果蝇卵子发生开始,一个细胞继续作为干细胞,另一个细胞成为胞囊干细胞。每个胞囊干细胞经历4 次有丝分裂,产生16 个细胞簇,由中胚层来源的滤泡上皮包围,形成卵室[4]。在4 次有丝分裂期间,不完全胞质分裂导致16 个生殖细胞通过一系列细胞质桥或环管相互连接,细胞质可以通过细胞质桥或环管流动。在16 个细胞中,1 个发育成卵母细胞,其他15 个充当护卫细胞[5]。通过14 个阶段的生长和发育,产生成熟的卵子。当与卵子发生有关的某个基因缺失或过表达就可能影响其生殖器发育,进而影响果蝇的生殖能力。
平面细胞极性(planar cell polarity,PCP)是许多上皮细胞的重要特征[6-7],其建立需要核心PCP基因和2 种钙粘蛋白(Fat 和Dachsous)。Fat 和Dachsous 起初被确定为肿瘤抑制因子[8-9],后来研究发现它们在PCP 调节中起重要作用[10-11]。果蝇卵巢卵室是研究组织伸长的理想模型[12],在卵子发生的初始阶段,伸长的卵泡(即卵室)围绕它们的前后轴(anterior-posterior,A-P)旋转。当卵泡细胞围绕其圆周轴迁移时,它们会形成1 个极化的细胞外基质[13]。一般认为,纤维化的细胞外基质和肌动蛋白丝的周向排列可提供一种分子紧身衣,它在生长过程中机械地限制卵室的周向扩张,从而促进卵室A-P 轴的优先伸长。Fat2 是一种与脂肪相关的钙粘蛋白。果蝇Fat2 含有1 个大的细胞外区域,其中包含34 个钙粘蛋白结构域、1 个层粘连蛋白A-G 结构域和膜近端区域中的5 个EGF 基序[14-15]。
转座子诱变技术可以获得基因突变体。Minos转座子最初分离鉴定自Drosophila hydei,属于Tc1/mariner 家族[16]。课题组前期获得1 个Minos转座子[17]随机插入的突变品系,对其生殖能力进行研究时发现:此纯合突变不可育,进一步研究发现突变的纯合雌蝇也不可育,而纯合雄蝇可育;解剖突变体纯合雌蝇卵巢发现处在14 阶段的卵室呈球形,背侧肢异常短小。通过反向PCR 法对Minos转座子插入位点进行鉴定发现:转座子插在kug(kugel)外显子内,kug也称为fat2。本研究中的突变体表型与fat2突变表型[18]一致。本研究对新的fat2突变体进行鉴定,以期为研究fat2基因功能提供新的研究材料。
1 材料与方法
1.1 果蝇品系
野生型果蝇品系w1118和y w;MI3266 FRT 3LR/TM6b,Tb(实验室自建)。
1.2 试验方法
1.2.1 果蝇育性检测
挑选突变品系纯合雄性果蝇和纯合雌性处女果蝇各10 只进行杂交,在培养管上记录日期并编号,以野生型果蝇w1118为对照组,杂交第3~10 天在体视显微镜下持续观察是否产卵、有无幼虫、蛹及成虫等情况,拍照并作好记录。
挑选突变品系纯合雌性果蝇10 只,与3 只野生型w1118雄果蝇杂交,检测雌蝇是否可育。挑选突变品系纯合雄性果蝇10 只,与10 只w1118野生型处女果蝇杂交,检测雄蝇是否可育。拍照并记录。
1.2.2 果蝇卵巢解剖及DAPI 染色观察
取待解剖的雌性果蝇用CO2在麻醉板上熏晕,用解剖镊夹取果蝇放至装有PBS 溶液的解剖皿中,于体视显微镜下从果蝇腹部第2 节处用解剖镊将外层表皮撕开,找到2 个卵巢并分离其周围组织,用镊子将卵巢放在装有PBS 溶液的EP 管内。用1 mL 移液枪吸走EP 管中的PBS 缓冲溶液,加入4%多聚甲醛1 mL 固定组织,于摇床上摇20 min;用1 mL 移液枪吸走4%多聚甲醛,向EP 管内加0.1% PBST 溶液1 mL 和DAPI 1 μL,于摇床上摇20 min;在体视显微镜下将卵巢周围的组织剥离干净,用镊子将卵巢放于载玻片中央,用指甲油封片,保存于4 ℃冰箱。于正置荧光显微镜观察果蝇卵巢的形态。
1.2.3 果蝇基因组DNA 提取
用装有研磨珠的1.5 mL EP 管收集5 只果蝇,放入-20 ℃中冷冻约1 h,加入DNA 裂解液200 μL 于组织破碎仪破碎3 min,将果蝇充分研磨成匀浆;70 ℃金属浴孵育30 min,14 800 r/min离心10 min;取上清液150 μL 到新的EP 管,加入异丙醇150 μL 和3 mol/L 醋酸钠(pH 5.2) 15 μL,混匀,静置10 min,14 800 r/min 离心10 min,看到絮状沉淀后,弃上清液;在EP 管中加入70%乙醇1 mL,14 800 r/min 离心5 min,重复2 次;弃乙醇溶液,空离心1 min,用移液枪吸掉剩余乙醇,静置至EP 管中乙醇完全蒸发后,加入超纯无菌水25 μL 溶解沉淀,完成果蝇基因组DNA 的提取。
1.2.4 果蝇基因组DNA 酶切
取果蝇基因组DNA 8.5 μL,限制性内切酶Sau3AI 0.5 μL,10×Buffer 1.0 μL,反应体系为10.0 μL。37 ℃于水浴锅中酶切2 h。
1.2.5 果蝇基因组DNA 酶切产物沉淀
向酶切产物中加入3 mol/L 醋酸钠1 μL 和无水乙醇1 mL,混匀,-20 ℃放置30 min;于4 ℃14 800 r/min 离心10 min,弃上清液;加入冰冷的70%乙醇500 μL,4 ℃ 14 800 r/min 离心5 min,弃上清液,晾干;加入ddH2O 25 μL。
1.2.6 果蝇基因组DNA 酶切沉淀产物连接及连接产物沉淀
取果蝇基因组DNA 酶切沉淀产物8.8 μL,T4 DNA Ligase 0.2 μL,10×Buffer 1.0 μL,反应体系为10.0 μL,于4 ℃冰箱中连接过夜。连接沉淀步骤同1.2.5 节。
1.2.7 反向PCR
反向PCR 是根据中间的1 段已知序列设计引物,分别向外扩增两端的未知序列。本研究利用Minos转座子的反向PCR 引物(F:CAAAAGCAACTAATGTAACGG;R:TTGCTCTTCTTGAGATTAAGGTA)对所插入位点的两侧DNA 进行反向PCR 扩增,从而找到Minos转座子插入位置。
反向PCR 扩增体系15.0 μL,包括连接沉淀产物6.5 μL,上游引物0.5 μL,下游引物0.5 μL,10×ExTaqBuffer 1.5 μL,dNTP Mixture 1.2 μL,TaKaRa ExTaq0.1 μL,ddH2O 4.7 μL。反 向PCR 扩增程序为:95 ℃预变性5 min,95 ℃变性30 s,50 ℃退火30 s,72 ℃延伸1 min,共35个循环;于72 ℃后延伸5 min,4 ℃终止反应。反向PCR 产物用1.0%琼脂糖凝胶电泳检测,经检测后的反向PCR 扩增产物送至生工生物工程(上海)股份有限公司进行测序,将测序结果于Flybase (http://www.flybase.org/)上进行BLAST 比对,找到Minos转座子插入位置,并判断其插入方向和影响的基因。
2 结果与分析
2.1 MI3266 生殖能力鉴定
如图1 所示:野生型果蝇w11183 d 时已有2 龄幼虫出现,5 d 时已有3 龄幼虫。与野生型w11183和5 d (图1a、e)相比,MI3266的雌性纯合处女果蝇与雄性纯合果蝇杂交后第3 天没有卵产生,且第5 天也未见卵及幼虫(图1b、f),而培养管内有菌滋生,说明MI3266突变品系是纯合不育的;雄性纯合MI3266与野生型w1118处女果蝇杂交后,第3 天有卵产生,且第5 天能观察到食物变稀且有大量卵孵化为幼虫(图1c、g),说明MI3266突变品系的纯合雄蝇是可育的;雌性纯合MI3266与野生型w1118雄性果蝇杂交后,第3 天没有卵产生,且第5 天未见任何卵及幼虫(图1d、h),培养管内有菌滋生,说明MI3266突变品系的纯合雌蝇是不可育的。
如图2 所示:杂交第7 天,野生型组以及纯合突变品系雄蝇与野生型处女果蝇w1118杂交组大部分均处于蛹期;而纯合突变品系雄蝇与纯合突变品系处女果蝇杂交组以及野生型雄蝇w1118与纯合突变品系处女果蝇杂交组均未出现3 龄幼虫和蛹,管内食物出现严重的裂开现象且有菌滋生,说明这2 个杂交处理无法产生后代。结合图1 和图2 可知:MI3266突变品系的纯合雌蝇不可育,其生殖系统可能存在可见的缺陷。

图1 杂交第3 和5 天各培养基内卵孵化情况Fig.1 The hatching of eggs in culture medium of the 3rd and the 5th day

图2 杂交第7 天各培养管内果蝇生长情况Fig.2 The growth of each vial on the 7th day of hybridization
2.2 MI3266 卵巢表型检测
野生型组w1118的果蝇卵巢结构正常,卵巢管由一串处在不同发育阶段的卵室构成,处于14阶段的卵室呈椭球型,背侧肢细长(图3a、c);突变品系组MI3266的卵巢结构形态异常,处于14阶段的卵室呈球型,背侧肢明显变短(图3b、d)。
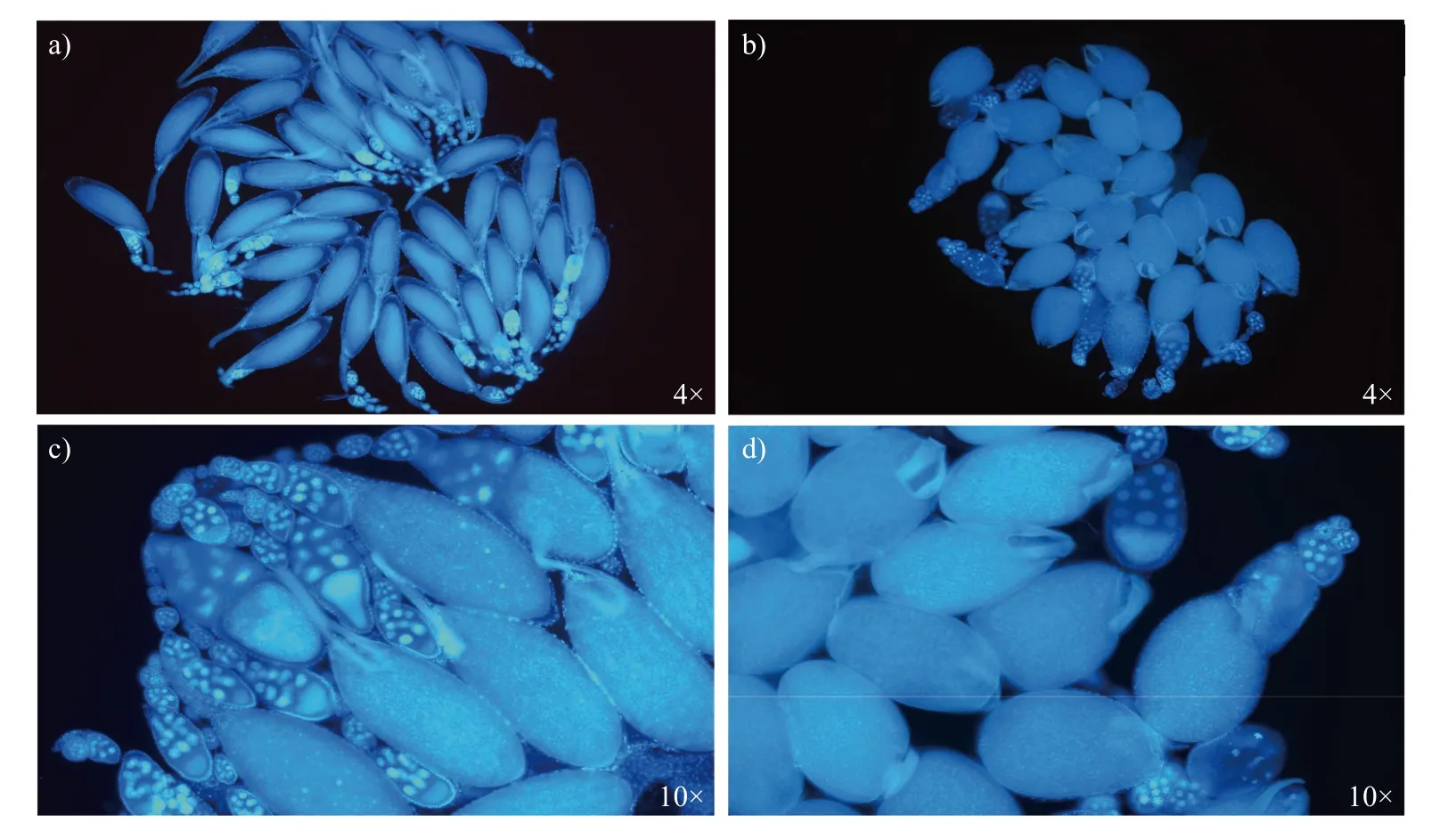
图3 果蝇卵巢DAPI 染色Fig.3 DAPI staining results of Drosophila ovary
2.3 MI3266 插入基因鉴定
由图4 可知:与DNA 2000 Marker 相比,突变品系扩增条带大小约为700 bp,水和w1118泳道无条带,说明扩增到MI3266插入基因组的特定条带。
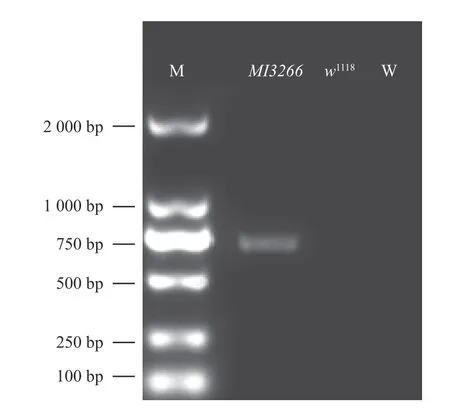
图4 电泳检测结果Fig.4 Electrophoresis results
反向PCR 扩增产物测序结果为:CCGGGTGGGAGGATGCATTGGTATGTGTTATCTTTAGTAGTATTGATAATATAGTGTGTTAAACATTGCGCACTGCAAAAAAACATGCTGTTCGAATTAATAGTGGTTGGGGCTCGTATATCCGTCTTGGATGTGAACGACAATTGCCCCTTGTTTGTCAATATGCCCTATTATGCCACAGTCTCTATTGACGATCCAACCCCTTGTGTCATGTCGGCGACCCTACGCCCCCAACTGAGAGAACTCAAAGGTTACCCCAGTTGGGGCACTACTCCCGAAAACCGCTTCTGACCTGGGAAGCTTCCGTTACATTAGTGCTTTTG。BLAST 结果显示:MI3266中的Minos 转座子插入到果蝇三号染色体左臂20015792 和20015793之间,突变基因为fat2,基因编号为CG7749。结合反向PCR 引物、BLAST 匹配序列方向与果蝇基因组序列的方向,判断转座子的插入方向为5′在右、3′在左,转座子位于匹配片段左侧,方向向左,与fat2基因编码方向相反,但插入fat2中部外显子区域,直接造成fat2突变。
3 讨论
组织器官表皮细胞的重要特征是具有极性,控制平面细胞极性(PCP)的分子途径首先报道自果蝇的研究。在果蝇翅和眼中形成PCP 需要保守核心PCP 基因和调节基因参与[10-11]。果蝇卵巢卵室表皮是研究PCP 的良好模型。早期发现的突变体kug其卵室呈球形,明显与野生型细长的椭圆形卵室不同,肌动蛋白纤维束方向与卵室形状有关[19]。野生型果蝇卵室表皮细胞里肌动蛋白纤维束方向垂直于果蝇卵室前后轴,而突变体kug卵室表皮细胞里肌动蛋白纤维束方向紊乱,故认为垂直于卵室前后轴分布的肌动蛋白束提供分子束缚力,限制卵室垂直方向的生长,使卵室呈细长的椭圆形。突变体kug分离于几十年以前,但直到2009 年才鉴定出其影响的基因为钙粘着蛋白Fat2[18]。目前,kug的突变体有很多种,最早报道的kugel 003突变体在第75 位赖氨酸发生终止,从而产生突变[18-19]。本研究将实验室自建突变体MI3266进行序列比对,Minos 转座子插入到果蝇三号染色体左臂20015792 和20015793之间。结合fat2 结构域可知:突变体在第19 个钙粘蛋白结构域发生突变,表型与fat2表型相同,经鉴定影响的基因是fat2,表明MI3266是fat2的新突变体。从果蝇到哺乳动物,Fat 蛋白家族都是保守的,果蝇有Fat 和Fat-like (Fat2) 2 个成员,脊椎动物有Fat-J、Fat1、Fat2 和Fat3 等4 个成员[20]。果蝇Fat2 与脊椎动物Fat1、Fat2 和Fat3 序列相似度更近。
肌动蛋白纤维束与培养的表皮细胞或者成纤维细胞基底部的应力纤维相似。应力纤维形成受整联蛋白影响。整联蛋白是跨膜的异二聚体,它把胞内肌动蛋白骨架和细胞外基质连接在一起。滤泡细胞里肌动蛋白纤维末端与整联蛋白相连,整联蛋白异二聚体成员mys和mew突变体卵室变圆,发现mys和mew影响肌动蛋白纤维束的极性[21]。受体酪氨酸磷酸酶Lar 与细胞外基质和肌动蛋白细胞骨架有关,它也影响肌动蛋白纤维束的极性[21]。Fat2 胞内区域与卵室延长无关,与Fat2在细胞内的分布密切相关[22]。在卵室发育1~8 时期,卵室会绕前后轴发生旋转,曾认为卵室旋转是卵室延长的先决条件。但是,去除胞内区的Fat2 在体外条件下其卵室可以延长至正常卵室长度,却不发生旋转,暗示卵室旋转不是卵室延长的先决条件[22]。研究发现:缺失胞内区Fat2 的卵室也可以旋转,只是速度减慢,因此,卵室旋转仍与卵室延长相关[23]。此外,Dystrophin-Dystroglycan 复合体也与卵室延长有关[24],但PCP 仍有很多问题没有解释清楚。根据反向PCR 扩增MI3266插入基因组序列和fat2对应的编码蛋白的核酸序列比对结果可知:MI3266插入在Fat2第19 个钙粘蛋白结构域,该结构域之后还有10 余个钙粘蛋白结构域、跨膜结构域和胞内区,因此,Minos转座子是插入到fat2胞外区。Minos转座子长7 267 bp,且MI3266插在fat2外显子内,如此大的转座子片段里很容易出现终止密码子。当蛋白翻译时,插入片段后面的编码序列无法翻译,导致Fat2 的胞内区、跨膜区以及部分胞外区缺失,而失去跨膜区的Fat2 是没有功能的[22],故推测fat2突变体的功能应完全缺失。
4 结论
本研究鉴定到1 个不育果蝇突变体,其雌蝇不育,且雌蝇卵巢晚期卵室呈球形,背侧肢短小。该突变影响的基因是fat2,其功能可能完全缺失。
