基于正交设计探究光照、温度、湿度对烤烟成熟生理的影响
2022-12-07卢绍浩张嘉雯赵铭钦
赵 喆,米 强,卢绍浩,张嘉雯,来 苗,王 坤,赵铭钦
(1.山东中烟工业有限责任公司 济南卷烟厂,山东 济南 250104;2.浙江中烟工业有限责任公司 技术中心,浙江 杭州 310024;3.上海烟草集团有限责任公司 技术中心,上海 201315;4.河南农业大学 烟草学院,河南 郑州 450002)
0 引言
随着全球气候变暖现象的加剧,不利气象因素对农业生产的影响日益增大[1],光照、温度、湿度作为主要的生态因子,影响着植物生长发育、成熟衰老的过程。植物光合作用随光照强度的变化而瞬时变化[2],当光照强度超过一定范围后,光合效率保持在最大值,过剩的光照强度易对细胞造成光损伤[3]。与正常叶片相比,高光照强度下鼠尾草叶片表现出更强的光保护需求,且其PSⅡ受损,叶片衰老加剧[4]。在连续光照情况下,随着光照强度的增加,生菜叶片内活性氧(ROS)的产生及脂质过氧化程度也逐渐增加[5]。
同时,温度作为最重要的环境因子之一,可通过调控净同化速率[6]和光合作用[7]影响植株生长。高温下植物体内叶绿素降解加快,活性氧增多,细胞易受损伤[8-9],据前人研究,温度大于35 ℃时超氧化物歧化酶(SOD)活性会降低[10];35~40 ℃时,净光合速率也会下降[11]。热处理提高了大豆体内乙烯含量,降低了光合作用速率和蔗糖含量,抑制了关键酶的促抗氧化剂活性,会导致叶片早衰[12]。同样,空气相对湿度也会影响植株光合作用及生理特性[13-14]。
目前,有关光照、温度对植物成熟衰老影响的研究较多,但涉及湿度影响的研究较少。因此,本文通过选取大田植物生长环境中最重要的3个气象因素,利用正交实验设计方法,初步探究不同因子下叶片光合特性、抗氧化系统的动态变化,以期为解析不同湿度条件对植株成熟的影响提供理论依据。
1 材料与方法
1.1 供试材料
本试验于2016年在江西省抚州市黎川县(东经116.52°,北纬26.89°)开展。供试材料为当地主栽品种云烟87。土壤为红壤土,其理化性状:pH值7.22,有机质8.72 g/kg,碱解氮51.27 mg/kg,速效磷16.84 g/kg,速效钾5.14 g/kg。试验地位于平坦开阔地块,通风良好,排灌、通电方便。烟苗于3月26日统一移栽,株距0.5 m,行距1.2 m,四周及各处理间设保护行,大田管理同当地常规水平一致。
1.2 试验设计
依据前人研究[15-17],本试验采取正交试验方法。根据其设计原则[18-19],利用SPSS 21.0软件进行3因素3水平的正交设计,具体设计如表1所示。选择不同光照强度(A)水平为50%(A1)、85%(A2)和120%(A3)3种自然光照强度,平均每日午间光照强度约为465.5×100、791.35×100、1117.2×100 lx;温度(B)水平为(29±1) ℃(B1)、(33±1) ℃(B2)、(37±1) ℃(B3)3种;湿度(C)水平为(64±1)%大气相对湿度(RH)(C1)、(73±1)% RH(C2)、(82 ±1)% RH(C3)3种。

表1 三因素三水平正交设计表
不同处理通过大棚温室精确调控烟株成熟期光照强度、温度、湿度3个指标。每个处理烟株约90株,通过透明无滴膜、遮阳网及植物灯(棚鲜牌36W)调控光照强度,并用照度计(TES-1330A)测定各处理光照强度;通过制冷机(YJL-18)、加湿器(浩奇HQ-2008A6)、除湿器(多乐信DR-1382L)、风机(贝德尔FZY-400)、控温插座(YWK-F)、控湿插座(W2105)调控各处理所需温、湿度条件。
大棚于6月20日(烤烟成熟中后期)搭建完毕并开始处理,每隔6 d各处理取样测定其生理指标,取样部位为从下至上第15~16叶位,每次处理3个重复,共取样3次。
1.3 测定项目与指标
光合色素用80%丙酮提取,按照Lichtenthaler等[20]的方法测定其含量。抗氧化酶类活性根据Wei等[21]的方法,采用NBT光还原法测定超氧化物歧化酶(SOD)活性,愈创木酚法测定过氧化物酶(POD)活性,紫外吸收法测定过氧化氢酶(CAT)活性。移栽后96 d,在上午9:30~10:30利用快速光合测定仪(ECA-PC0401)测定叶片光合速率,每个处理5次重复。
1.4 数据分析
采用SPSS 21.0软件进行正交设计、方差分析,采用Microsoft Excel 2013软件制作表格,Origin 8.0软件绘图。
2 结果与分析
2.1 光照强度、温度、湿度3个因子对成熟后期叶片光合速率的影响
2.1.1 不同因子水平下烤烟叶片光合速率影响的边际值分析 光合速率的边际值为所有处理中某一因子水平下光合速率的平均值,可以表示某因子水平变化对叶片光合速率的影响效应。由各因子光合速率边际值可知(表2),随着光照强度的升高,光合速率呈先升高再降低的变化趋势,3个光照强度水平下以85%光照强度时光合速率最高,85%和120%光照强度对应的光合速率间差异未达到5%显著水平。随着温度的升高,光合速率呈逐步降低的趋势,38 ℃时对应的光合速率显著低于29 ℃的。随着空气相对湿度的增加,光合速率呈先升高再降低的变化趋势,但差异未达到5%显著水平。
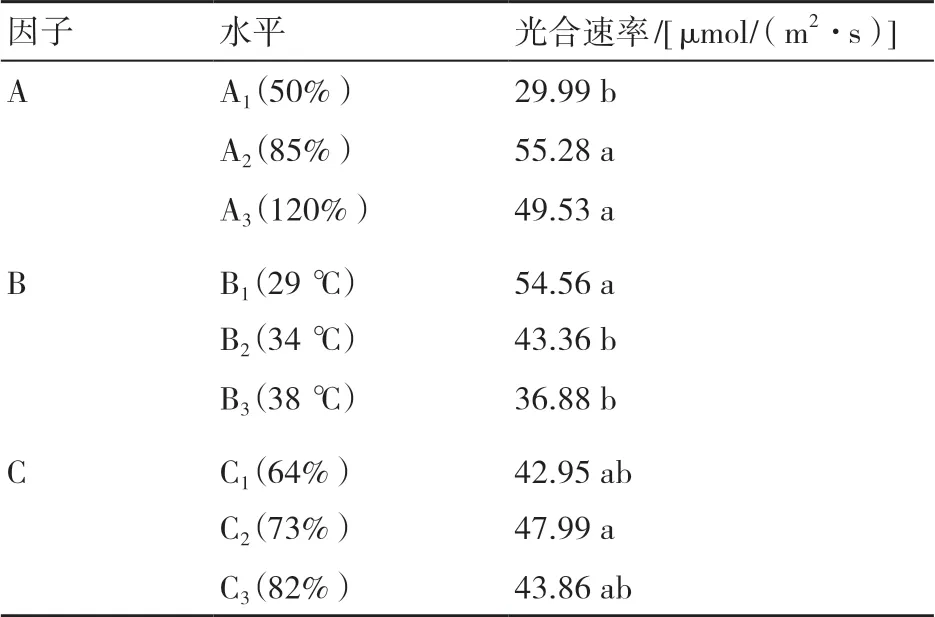
表2 不同因子水平下烤烟叶片光合速率的边际值
2.1.2 生态因子对烤烟叶片光合速率影响的主效应分析 采用正交试验中常用的极差分析法[15,17]进行数据分析。极差越大,即因子的效应值越大,表明因子变化对结果的影响越大。由表3可以看出,3个生态因子对于光合速率的影响均达到极显著水平,且对光合速率的影响排序为光照强度>温度>湿度,光照强度为光合速率的主要影响因子。
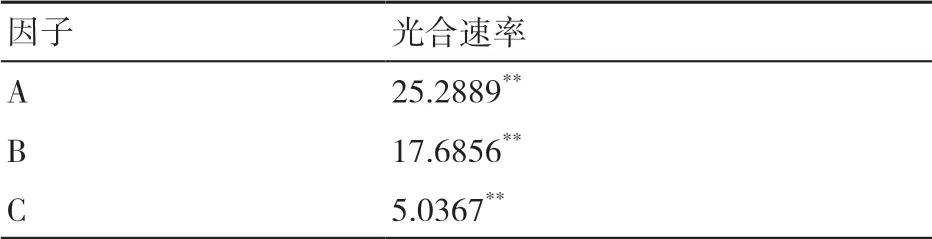
表3 各因子对光合速率影响的极差 μmol/(m2·s)
2.2 光照强度、温度、湿度3个因子对成熟后期叶片光合色素含量的影响
2.2.1 不同因子水平下烤烟叶片光合色素含量影响的边际值分析 由图1A可知,随着光照强度的增加,3个时期叶绿素a(Chl a)含量均呈降低的趋势,且以移栽后90 d弱光条件下降幅度最大;随着温度的升高,3个时期Chl a含量同样呈下降的趋势;从湿度变化可以看出,移栽后90 d在82%湿度条件下Chl a含量最高,移栽后96 d在73%湿度条件下Chl a含量最高,而移栽后102 d则在64%湿度条件下表现最好,但彼此间并无显著差异。
由图1B可知,叶绿素b(Chl b)含量随着光照强度、温度的增大而逐渐降低,但随着湿度的增加呈现逐渐升高的变化趋势。叶片衰老期间,叶绿素较易降解,而类胡萝卜素(Carotenoids)较为稳定,故衰老期间叶片表现为黄色。
由图1C可知,3个时期中Cars含量随着光照强度的升高而增大,随着温度的升高而减小,其随湿度的变化情况与Chl a含量随湿度变化情况类似,都是移栽后90 d在82%湿度条件下、移栽后96 d在73%湿度条件下、移栽后102 d在64%湿度条件下Cars含量最高。
由图1D可知,光照强度与Chl a/b值间存在正相关关系,随着光照强度的增加,成熟后期3个阶段Chl a/b值皆升高。同时,随着温度的升高,Chl a/b值同样升高。湿度与Chl a/b值变化存在负相关关系:当湿度在64%条件下时,3个时期的Chl a/b值皆为最大。Chl a/b值大,表示Chl a降解较Chl b慢,即在相同叶绿素含量时,64%湿度条件下叶片对蓝紫光的吸收不如其他湿度条件的。

图1 不同因子水平下烤烟叶片Chl a、Chl b、Cars、Chl a/b值的边际值
2.2.2 生态因子对烤烟叶片光合色素含量影响的主效应分析 由各因子对光合色素含量影响的极差可以看出(表4),光照强度对Chl a含量的影响最大,3个时期皆达到极显著水平;随着时间的增加,叶片Chl a含量对于外界光照强度及温度的响应逐渐变弱。温度和湿度条件下,Chl a的极差基本上未达到显著水平。对于Chl b含量来说,光照强度为主要影响因子,温度为次要影响因子,湿度影响最小。
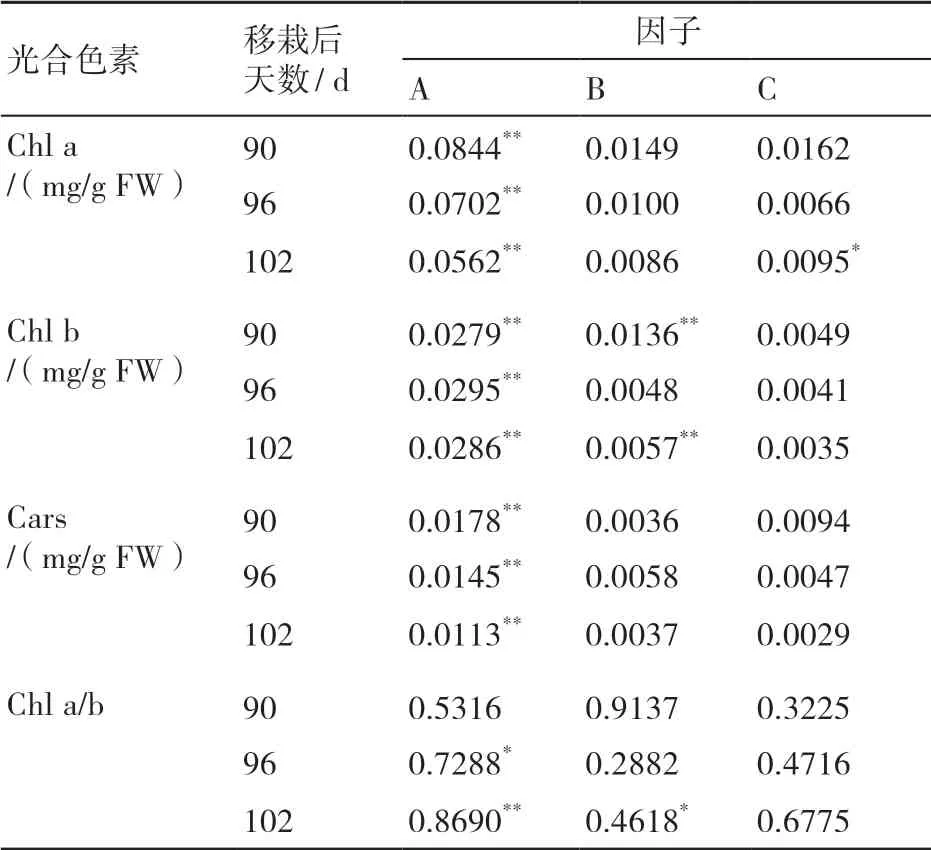
表4 各因子对光合色素含量影响的极差
在Cars含量的极差中,光照强度在3个时期的极差均最大,3个时期均达到极显著水平,其影响随着时间的推移逐渐减弱。温度对Cars含量影响的极差在各个时期变化较小。湿度因子在移栽后90 d对Cars含量影响的极差仅次于光照强度因子,在其他2个时期极差减小。
Chl a/b值对于3个因子的响应中,移栽后90 d,温度对其影响的极差最大;其他2个时期,光照强度对Chl a/b值影响的极差最大,移栽后102 d时达到极显著水平,湿度影响次之,温度影响最弱。
2.3 光照强度、温度、湿度3个因子对成熟后期叶片抗氧化酶类活性的影响
2.3.1 不同因子水平下叶片抗氧化酶类活性影响的边际值分析 由图2A可知,SOD活性分别随着光照强度、温度、湿度的增加而显著降低。POD活性随着光照强度和温度的升高而增强,尤以不同光照强度下最为显著;不同湿度水平下,在64%RH时3个时期的POD活性均为最高,之后随着湿度的增加而显著降低(图2B)。CAT活性在不同因子水平下变化不一:随着光照强度的增加,整体呈上升趋势;随着温度的变化则呈先升高后下降的趋势;而随着湿度的增加,CAT活性呈逐渐降低的变化趋势。
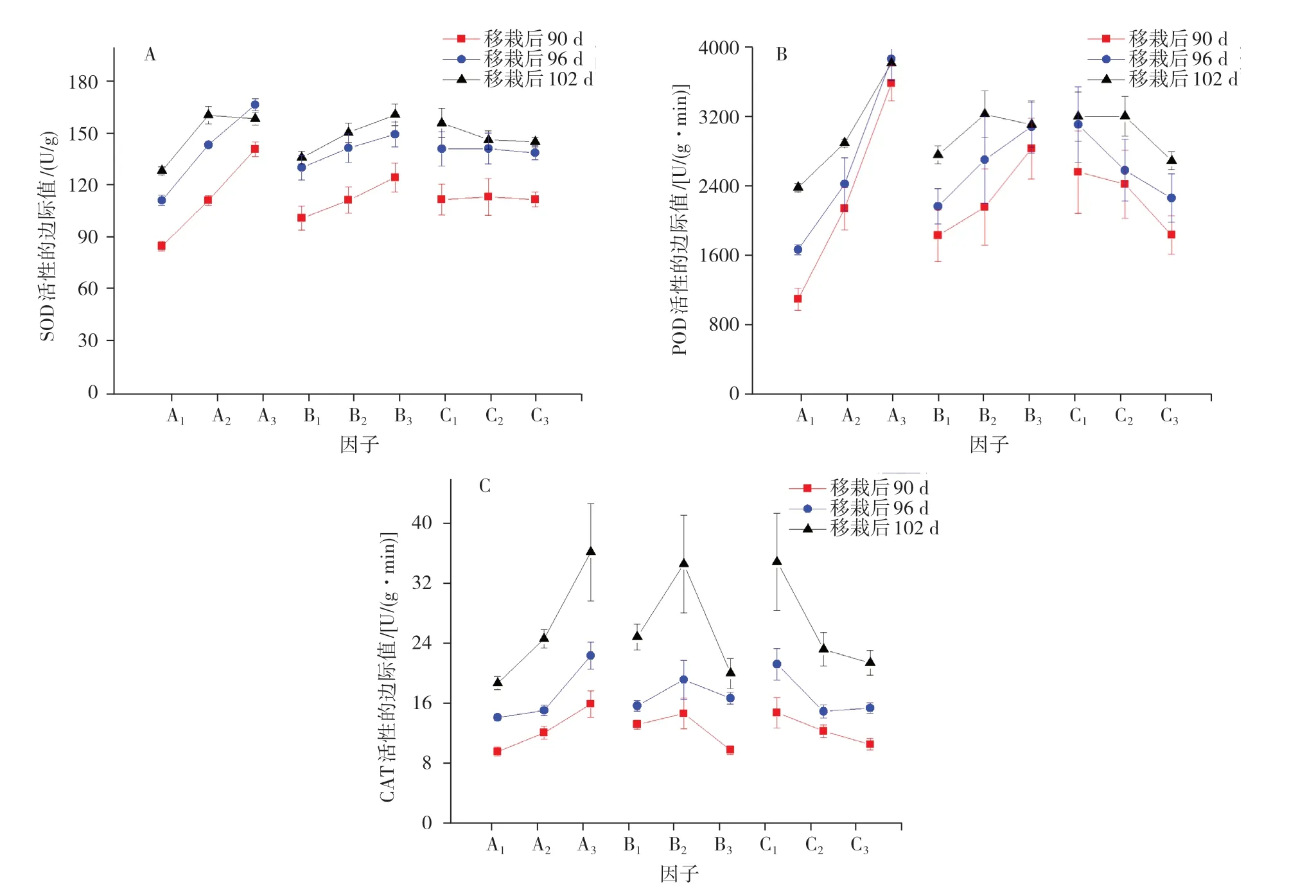
图2 不同生态因子对烤烟叶片SOD、POD、CAT活性的边际值
2.3.2 生态因子对叶片抗氧化酶类活性影响的主效应分析 极差的变化可以反映各因子影响力的变化。由表5可知,在光照强度、温度、湿度3个因素中,光照强度对SOD、POD、CAT活性影响最大,是抗氧化酶类活性的主要影响因子,温度对抗氧化酶活性影响次于光照强度,湿度影响最小。在不同时期,SOD、POD活性对光照强度的响应逐渐减弱,CAT活性对光照强度的响应逐渐增强,移栽后102 d时湿度对CAT活性的影响逐渐增强。
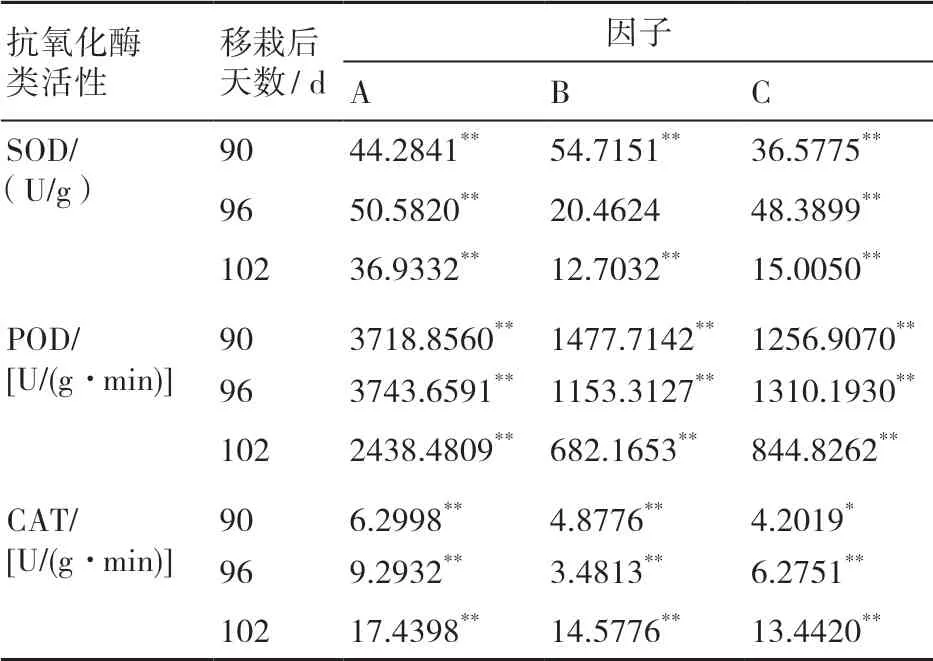
表5 各因子对抗氧化酶类活性影响的极差
3 讨论
光合作用是植株体内物质形成的基础[22-23],作为一个复杂的生理过程,植物光合作用受外界环境因子调节,对环境变化极其敏感[24]。本研究表明,光照强度是影响叶片光合速率的关键因子,温度为次要因子,湿度对光合速率的影响最小。随着光照强度的增加,光合速率呈先升高再降低的变化趋势,表明高光照强度使叶片出现了光抑制现象;随着温度的升高,光合速率呈逐步降低的趋势,光合作用减弱;随着湿度的增加,光合速率呈先升高再降低的变化趋势;这与Xu[3]、Elias[25]等的研究结果一致。
光合色素包括叶绿素和类胡萝卜素2类,叶绿素负责光能的吸收、传递和转化,类胡萝卜素则负责捕获光能及光破坏防御[26]。在成熟后期不同阶段,Chl a含量与光照强度、温度呈负相关关系,但其最适湿度则随着时间的推移而逐渐变小。Chl b的含量,在成熟后期不同阶段,随光照强度的升高而显著降低,但对于温度和湿度变化的响应并不灵敏。
胁迫条件下,植物能够通过叶黄素循环进行热耗散,减轻光氧化的伤害[27-28]。本研究中,高光照强度下Cars含量显著增加,有利于植株消耗过剩的光能,保护叶片不受强光损伤;但高温条件下Cars含量则较低。对于湿度来讲,随着叶片衰老程度的加剧,其最适湿度条件由82%、73%最终变为64%。可以推断,烤烟成熟后期高温、高湿条件可能会阻碍类胡萝卜素的产生,影响叶黄素的循环,不利于叶片通过叶黄素循环耗散过剩光能。
光照强度减弱,Chl a/b值减弱,有利于捕获漫射中占优势的蓝紫光,在一定程度上提高了光能的利用率,是植物对于弱光的适应反应;同时叶绿体对2,6-二氯酚靛酚的还原能力增加,叶绿体光合磷酸化活性提高[29]。当植物的光合机构接受的光能超过所能利用的限度时,则会发生光抑制,产生过剩激发能[30]。烤烟成熟后期,当外界湿度大时,Chl a/b值较低,若此时强光照射,植物会捕获更多的蓝紫光,使得光抑制现象严重加剧,造成光损伤及光合机构的破坏。仅从Chl a/b角度上考虑,高光照强度和低湿度条件较为相配。综合边际值结果可知,烟株成熟后期光照强度对光合色素含量影响最大,温度和湿度条件是次要因子,光合色素含量受三者共同调控。
强光下,过剩的光能通过电子传递链与分子氧结合形成超氧阴离子自由基[31]。抗氧化系统中,SOD将超氧阴离子自由基转化为H2O2,过剩的H2O2则被POD和CAT清除,以抵御细胞氧化损伤[32-33]。本研究表明,烤烟成熟后期SOD活性受光照强度、温度、湿度3个因子共同调控,且随三者的增大而降低。即植株在高光、高温、高湿条件下,叶片对超氧阴离子自由基清除能力下降,根据自由基伤害假说[34]可知,膜脂过氧化作用加剧,叶片易受损伤,导致叶片提前衰老。POD和CAT活性则主要受光照强度影响。值得注意的是,高光照强度、高温条件下使得POD活性升高,而高湿条件下则使得POD活性降低,即高光照强度、高温条件时,叶片清除过氧化物能力增强,但高湿条件时则阻碍了这一清除过程。
本研究结果表明,光照强度、温度、湿度作为主要的3个生态因子,协同调控着烤烟的成熟衰老。光照强度是影响成熟期烤烟光合作用及抗氧化活性的主要因子,温度和湿度这2个因子影响力稍弱。成熟后期不同阶段,各生态因子对烤烟的影响规律基本一致。其中光照强度和温度因子表现为相同的影响趋势,湿度因子在部分光合及抗氧化指标中表现出与光照强度、温度因子不同的影响趋势。
