补播时间和补播草种对退化草甸草原植物群落的影响
2022-12-01刘玉玲王德平张泓博王国富李鹏珍戎郁萍
刘玉玲,王德平,张泓博,王国富,李鹏珍,戎郁萍*
(1.中国农业大学草业科学与技术学院,北京 100193;2.呼伦贝尔市陈巴尔虎旗西乌珠尔苏木综合保障和技术推广中心,内蒙古 呼伦贝尔 021500)
气候变化和不合理的利用导致草地生产力和优质牧草比例降低。草地补播是指在不破坏或少破坏原有植被的情况下,通过人工建植一些适应性强、有价值的优良牧草,来提高草地产量和品质的一种草地修复手段[1]。适宜的补播时间是确保补播成功的重要因素,不同播期的气候条件差异导致温度和降水差异,进而影响植物生长发育[2],刘锦昉等[3]发现在干旱半干旱区和湿润半湿润区生物量随年均温和降水量增加而增加。不同地区适宜的补播时间存在较大差异,吉林省中东部地区适宜春播[4],而在春季干旱多风、蒸发量大的地区适宜雨季补播[5],在内蒙古鄂尔多斯地区适宜秋播,最晚应在9月中旬前补播[6],甚至在环青海湖地区可以采用临冬寄籽方式补播[7],种子待来年春季水热条件满足时开始萌发[8]。因此,满足补播条件的播期主要是春季返青初期、夏季生长季及临冬3个时期。研究补播时间对产量影响的物种多为燕麦(AvenasativaL.),对其他牧草研究较少,周磊等[9]发现在北方农牧交错带推迟播期提高燕麦产量,在半干旱山区5月6日补播燕麦可提高产量和品质[10]。目前关于呼伦贝尔地区适宜补播时间的研究相对较少,为选择适宜的补播时期,有必要探究3个主要补播时间对呼伦贝尔草地生产力和多样性的影响。
适宜的补播草种是决定补播草地生产力的一个关键因素,大量研究表明当地多年生草种具有适应性强、建植率高等优点[11-13],张英俊等[1]建议选择对退化草原土壤呈中或正反馈效应的优质禾本科或豆科草种进行补播。闫子盟等[14]和杨增增等[15]发现补播豆科牧草和多年生禾草可以提高草地生产力、牧草品质及群落稳定性,物种丰富度和Shannon-Wiener指数等也有所提高但差异并不显著。另外,Foster等[16]、Zobel等[17]及王伟等[18]发现在不同类型退化草地补播,植物群落发生持续变化,且物种丰富度呈增加趋势,说明补播具有长期性影响。草原区降水变异大、气候波动强[19],应根据当地环境条件及退化程度选择适宜当地的补播时间及补播草种,从而制定合理的补播方案。近年来,呼伦贝尔草原生产力和优质牧草比例逐渐下降[20],畜牧业是呼伦贝尔农业经济的重要组成部分,对草产量及品质的要求较高,恢复草地生产力和品质是解决草畜矛盾的关键。本试验根据呼伦贝尔草甸草原退化情况选择合适的补播时间和补播草种,并持续追踪补播后植物生物量及多样性的变化,以期为呼伦贝尔草甸草原制定合适的补播方案提供理论基础。
1 材料与方法
1.1 试验地概况
试验区位于呼伦贝尔市海拉尔区特尼河九队(49°20′~49°26′ N,119°55′~120°9′ E,海拔628~649 m),草原类型是以羊草(Leymuschinensis(Trin.)Tzvel.)和贝加尔针茅(StipabaicalensisRoshev.)为建群种的温带草甸草原。该区域属温带大陆性季风气候,年际间温差变异大,年均温—2℃~—1℃,1月平均气温—24.0℃,7月平均气温20.5℃;1951—2020年年均降水量为353.6 mm,全年75%降水量集中于6—9月。土壤为黑钙土或暗栗钙土,有机质含量5.1%,有机碳含量33.15 g·kg-1,全氮含量2.96 g·kg-1。
1.2 试验设计
采用补播时间和补播草种双因素随机区组试验设计设置3个补播时间:2017年7月1日夏播(RT1),2017年9月30号秋播(RT2)和2018年5月1日春播(RT3);5个补播草种(扁蓿豆(MelissilusruthenicusL.,RS1)、黄花苜蓿(MedicagofalcataL.,RS2)、杂花苜蓿(Medicagovaria,RS3)及两个羊草品种—野生羊草(RS4)和‘中科1号’羊草(RS5)),以开沟不播种作对照处理(CK)。试验共3个区组,每区组6个处理,3次重复,共45个小区。区组间隔10 m,小区间隔2 m,小区为3 m×4 m。补播前对小区进行刈割处理,留茬高度为5 cm,开沟深度为5~7 cm,播种深度为2~3 cm,行距为30 cm,补播完成后原位土镇压回填。补播种子播量在20~50 kg·hm-2[8],赵娜等[21]在锡林郭勒盟典型草原的紫花苜蓿和羊草播量分别为22.5 kg·hm-2和37.5 kg·hm-2。由于呼伦贝尔地区气候变化大,草地枯落层厚及土壤紧实等因素不利于种子的萌发,按照种子的实际用价,加大种子播量,种子播量设置为野生羊草64.5 kg·hm-2,‘中科1号’羊草45 kg·hm-2,扁蓿豆和黄花苜蓿30 kg·hm-2,杂花苜蓿39.5 kg·hm-2。
1.3 样品采集与分析
2018—2020年每年8月中旬对小区进行群落调查,在每个小区内随机放置一个1 m×1 m的样方,测量样方内各物种密度。在调查样方内随机选取1 m×0.25 m的样方,将样方内植物分种齐地面刈割装入信封,并带回室内烘干至恒重后称量,计算小区产量。在Excel中对调查数据进行初步整理,计算Shannon-wiener指数(H′)、Simpson优势度指数(D)。
Shannon-wiener指数:
H′=-∑(Pi)ln(Pi)
Simpson优势度指数:
D=1-∑Pi2
式中Pi为此物种的个体数占群落中总个体数的比例。
2018—2020年试验地的气候数据收集整理来源于中国农业科学院呼伦贝尔站(http://www.hulun.cn)。变异系数的计算公式为:变异系数(CV)=(标准偏差/平均值)×100%。
1.4 统计分析
在Excel中初步整理样方数据,计算禾本科、豆科、其它科的生物量。在R 4.1.2中进行3因素方差分析,分析补播时间、补播草种、年份及其交互作用对群落生物量、功能群比例、Shannon-wiener指数和Simpson优势度指数等的影响。处理间的多重比较采用Duncan法,箱式图使用R中“ggplot 2”包制作,文中用表使用Word 2019制作。
2 结果与分析
2.1 补播时间和补播草种对地上生物量的影响
补播时间和补播草种对地上生物量无显著影响,年份、年份与补播时间交互作用显著影响地上生物量(表1,P<0.05)。2018年地上生物量分别比2019和2020年高69.92 g·m-2和59.9 g·m-2(图1,P<0.05)。地上生物量在2018年最高,2019和2020年地上生物量在RT3处理下高于其它两个补播时间(图1,P<0.05)。另外,地上生物量与年际间降水变异系数呈显著负相关关系(R2=0.58),与积温变异系数呈显著正相关关系(R2=0.40)(图2,P<0.05)。气温变异系数(CVT)和降水变异系数(CVP)对RT1处理下地上生物量解释度最大(R2=0.65),其中CVT解释了0.19,CVP解释了0.47(表3,P<0.05)。

表1 植物地上生物量、物种多样性及年份的三因素方差分析
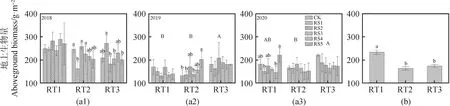
图1 不同补播时间和补播草种下植物地上生物量的年变化(a),和补播处理地上生物量的年变化(b)
2.2 补播时间和补播草种对群落结构的影响
补播时间显著影响豆科比例、物种丰富度、Shannon指数和Simpson指数,补播时间与年份交互作用显著影响禾本科和豆科比例(表1,P<0.05)。在2018年,禾本科比例在RT1处理下显著高于RT2和RT3,豆科比例在RT3处理下显著高于RT1和RT2,其他科比例在RT2处理下显著高于RT1,物种丰富度在RT2处理下显著高于RT1和RT3(图3,P<0.05)。在2020年,豆科比例在RT1处理下显著高于RT2和RT3,Shannon和Simpson指数在RT1处理下显著高于RT3(图3,P<0.05;图4,P<0.05)。生长季累积积温和降水与Simpson指数呈显著负相关关系,而降水变异与禾本科比例呈显著正相关关系,与其他科比例和物种丰富度呈显著负相关关系,积温变异与物种丰富度呈显著正相关关系(图2,P<0.05)。CVP对RT1处理下禾本科生物量解释度最大;CVT和CVP对RT1和RT2处理下其他科生物量的解释度较高,R2分别为0.516和0.503(表3,P<0.05)。
补播草种显著影响禾本科比例、豆科比例、Shannon指数和Simpson指数,补播草种与补播时间交互作用显著影响物种丰富度(表1,P<0.05)。禾本科比例在2020年RT1处理下补播RS4显著高于CK,豆科比例在2019年RT2和RT3处理下补播RS1显著高于CK(图3,P<0.05)。物种丰富度在2018年补播草种和补播时间处理下差异显著,其中在RT1处理下各补草种均显著低于CK,在RT2处理下补播RS2显著低于CK,在RT3处理下补播RS4显著低于CK;在2019年RT3处理下补播RS3显著高于RS5,但各补播草种与CK无显著差异;在2020年RT2处理下补播RS1显著低于CK(图4,P<0.05)。Shannon指数和Simpson指数在2020年RT3处理下补播RS4显著低于CK。

图3 补播处理对2018—2020年禾本科比例(a)、豆科比例(b)、其他科比例(c)的影响

图4 补播处理对2018—2020年植物群落物种丰富度(a)、Shannon-wiener指数(b)、Simpson优势度指数(c)的影响
年份显著影响禾本科比例、其他科比例、Shannon-Wiener指数、Simpson优势度指数和物种丰富度(表1,P<0.05)。随年份增加,其他科比例、物种丰富度、Shannon指数和Simpson指数呈下降趋势(表2,P<0.05)。
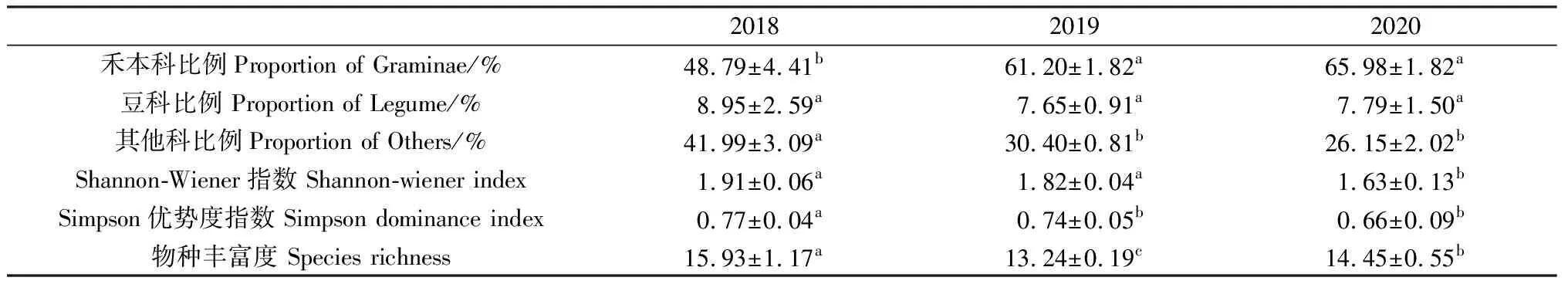
表2 不同年份间植物各功能群比例及多样性的影响
3 讨论
补播时间是影响补播效果的关键因素,不同补播时间较大的水热条件差异是影响补播草种萌发的重要因素[22]。补播当年夏播和春播地上生物量较高,而补播第2和3年地上生物量在春播处理下最高。由于补播当年较为干旱[22],可能导致补播草种定植率低;另外,夏季播前刈割和划破草皮处理对补播当年原生植被破坏性较大[7],同时增加地面蒸腾量,导致土壤含水量较低,不利于草地恢复。而春播前刈割对原生植被损伤较小,在一定程度上降低了原生植被遮阴作用及物种丰富度[23-24],进而提高补播草种光竞争能力,增加禾本科比例和豆科比例;且相较于夏播和秋播,春播恢复时间较长,有助于克服夏播带来的这种不利影响,进而有利于提高草地生产力[25]。秋播有利于其他科比例和物种丰富度提高,秋播播前通过刈割处理增加地上枯落物,进而有利于加速养分周转[26],改善土壤理化性质,从而利于群落多样性的提高。划破草皮处理能够为早春一年生阔叶植物种子萌发和生长提供光照和养分,降低禾本科和豆科竞争优势,从而增加其他科物种竞争优势和物种多样性[26]。
补播羊草和豆科草种影响群落结构,提高了禾本科比例,并降低其他科比例,有助于提高草地品质。Lambers等[27]发现高产物种是氮竞争者或氮固定者,氮素是影响植物生长发育的重要因素,豆科植物的固氮作用有利于增加土壤有效氮含量[28],有助于补播物种获取氮素,所以补播扁蓿豆有助于增加豆科比例和物种多样性,但并未显著提高地上生物量。补播第3年豆科比例在补播草种间无显著差异,这表明天然草地在自然条件下很难通过一次补播长期改变豆科比例。但是,补播羊草使禾本科比例增加,一方面可能是因为羊草通过补播和无性繁殖策略表现出竞争优势,另一方面也可能是补播豆科牧草能够促进土壤氮循环[29],导致禾草获取更多的氮增加竞争优势,从而提高禾本科比例。
年份显著影响补播的地上生物量、功能群比例及物种多样性,年份间差异来源主要是生长季温度和水分的变异。随补播年限增加,地上生物量、其他科比例和群落物种丰富度均显著降低,禾本科比例增加。温度和水分是植物生长的限制因素,北方六省草地生产力与积温和降水量呈显著正相关关系[30],而Liu等[31]和Wood等[32]分别对中国和美国植被驱动因子进行分析发现生产力与降水量呈正相关关系,与积温呈负相关关系。本试验中地上生物量与生长季积温变异呈显著正相关关系,与生长季降水变异呈显著负相关关系。近年来全球变暖,植被驱动力由温度转变为水分[33],大量研究表明降水时变异降水总量对牧草生产力影响更大[34-35],且生物量随降水变异增加而增加[36],这与本试验结论一致。气温和降水大幅波动同时也导致物种丰富度降低[36-37],生物量与物种丰富度的关系主要呈单峰型[38-39]和线性[40-41],本试验物种丰富度与生物量呈线性相关。不同年份下植物群落结构发生改变,由于群落不同功能群间存在竞争[42],补播草种的定植会提高禾本科和豆科植物的竞争优势,增加禾本科与豆科比例,同时降低其他科比例;另外,划破草皮可能通过降低土壤紧实度,刺激羊草的无性生殖,提高禾本科优势度[43],从而导致禾本科比例增加。而豆科草种在天然草地存活率低,养分、光照的限制导致豆科比例未发生显著变化[44]。
4 结论
生长季温度和水分的变异是影响补播的重要自然因素,地上生物量与生长季积温呈正相关关系,与生长季降水量呈负相关关系。补播当年较为干旱的气候和春播处理较短的恢复时间导致补播当年夏播和春播生物量较高,而在补播第2和3年春播生物量及物种丰富度高于夏播处理。补播羊草显著提高禾本科比例,降低其他科比例。随补播年限增加,其他科比例、物种丰富度、Shannon-Wiener指数和Simpson优势度指数显著降低,而禾本科比例显著增加。综上,呼伦贝尔地区在干旱年份适宜春季补播羊草来修复草地。
