解淀粉芽孢杆菌DGL1促燕麦生长分子机制及代谢通路探究
2022-12-01谢永丽乔有明陈海龙武玲玲
杨 雪,王 添,谢永丽,*,乔有明,陈海龙,陈 兰,武玲玲
(1.青海省青藏高原优良牧草种质资源利用重点实验室,青海 西宁 810016;2.青海大学省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016;3.青海大学农牧学院,青海 西宁 810016)
燕麦(Avenasativa)是一年生禾本科植物,植株高大,须根系发达,覆盖面积广,具有耐寒、耐盐碱、抗旱等特点,适宜青藏高原独特的自然地理环境,是治理草地退化的先锋物种[1-2]。同时,燕麦的茎秆嫩而多汁,营养丰富,适宜青饲和晒制干草,是优质的饲草饲料,能够解决青藏高原冬季家畜饲草短缺及营养不足的问题[3-5]。因此,燕麦对于高寒地区畜牧业生产和生态建设都具有重要的意义。芽孢杆菌作为重要的植物根际促生菌[6],可以通过产生植物激素[7]、增加植物对多种营养元素的吸收[8]、诱导植物系统抗性[9]、与病原菌竞争生态位[10]等方式与寄主植物建立有益的互作关系,提高植物叶绿素、脯氨酸的含量,且增加农作物的产量,为农业可持续发展带来了新的机遇[11]。因此,了解微生物-植物之间的互作机制,使有益菌能够更好的适应环境,对提高植物产量和品质具有重要的应用价值。
转录组测序技术能够从整体研究物种在某一发育阶段或功能状态下基因的表达水平,并因其灵敏度高,能发现新转录本等特点,为研究植物生长发育过程的分子机理奠定基础,被广泛应用于农业、临床医学、食品营养和卫生等多个领域[12]。因此,从植物响应芽孢杆菌的角度了解促生机制,通过转录组测序分析有助于更好地了解植物应答根际促生菌信号传导通路的复杂网络。Sun等研究发现枯草芽孢杆菌TR21通过调节JA(茉莉酸)和BRs(油菜素内酯)生物合成途径,提高香蕉植物对尖孢镰刀菌(Fusariumoxysporum)的抗性,缩短生长期并增加香蕉的产量[13]。芽孢杆菌还可以提高植物对逆境胁迫的耐受能力,Samaras等报道称枯草芽孢杆菌MBI600能够激活水杨酸信号通路中的基因PR-1A和GLUA,茉莉酸/乙烯信号通路中的基因CHI3,LOXD,PAL等与防御机制相关基因的上调表达,这些结果表明,在番茄植株上施用促生菌后激活了番茄系统抗性,增强了对病原菌的抵抗能力[14]。
解淀粉芽孢杆菌DGL1分离自海拔3 010 m的大格勒干旱沙地白刺(N.tangutorum)根围,具有较好的固氮活性和降解纤维素活性,且能够抑制锐顶镰孢病菌(Fusariumacuminatum)、瓜类枯萎病菌(Fusariumoxysporum)、小麦赤霉病菌(Fusariumgraminearum)病菌,并发现菌株DGL1能够显著提高燕麦‘青燕1号’的发芽率、根长、芽长[15],但其促生分子机制尚不清楚,本研究通过转录组测序技术初步探究芽孢杆菌DGL1对燕麦品种‘青燕1号’促生作用的调控机理,以期为芽孢杆菌菌肥促高寒牧草生长提供理论依据。
1 材料与方法
1.1 试验材料
植物材料:燕麦(Avenasativa)品种为‘青燕1号’,由青海省青藏高原优良牧草种质资源利用重点实验室提供。
供试菌株:解淀粉芽孢杆菌DGL1(B.amyloliquefaciens)保藏于-80℃超低温冰箱。
1.2 DGL1菌悬液制备
采用三区划线法将DGL1菌株接种于无菌的LB固体培养基上,37℃恒温培养12 h后,再将新长出的单菌落接种于20 mL LB液体培养基中,在恒温摇床上(37℃,200 r·min-1)培养12 h,将发酵液离心收集菌体,用无菌水调整菌液细胞浓度为106cfu·mL-1,制成芽孢杆菌DGL1菌悬液[16]。
1.3 燕麦与DGL1菌悬液互作
挑选种皮完好、大小均等、无病虫害的燕麦草种‘青燕1号’,将种子置于20% 次氯酸钠溶液消毒处理30 min,无菌水反复冲洗5次后,播入灭菌营养土的穴盆中(30株·盆-1),每3 d浇灌20 mL蒸馏水,培养条件为25℃,光周期16 h/8 h;将培养12 d的燕麦幼苗从穴盆中取出,分为5组,无菌水清洗根部数次,滤纸吸去根部水分;其中4组燕麦根部完全浸于细胞浓度为1×106cfu·mL-1的DGL1菌悬液中,建立燕麦与菌液DGL1的互作体系,互作时间为2 h,4 h,8 h,12 h,其余1组为燕麦根部浸于无菌水处理,作为CK对照组(图1)。

图1 燕麦根部与菌悬液互作
1.4 燕麦叶部RNA提取及测序文库构建
将互作2 h,4 h,8 h,12 h的燕麦植株分别从菌悬液取出,滤纸吸去根部液体;快速用灭菌剪刀剪断叶部,分别用锡纸包好,液氮速冻后置于-80℃超低温冰箱保存待用,每一处理组3次重复。样品委托上海美吉医药生物公司完成RNA提取、测序和文库构建。转录组测序试验步骤如下:将燕麦叶部提取总RNA(5组样品,每组3个重复,共15个样品),对提取到的15个样品进行RNA质检,将完整的mRNA片段化,将mRNA在逆转录酶的作用下反转成cDNA,随后合成二链,形成稳定的双链结构,最后上机测序[17]。
1.5 转录组数据分析
1.5.1差异表达基因分析 运用TopHat2软件将测序数据与燕麦参考基因组序列进行比对;使用DESeq2软件对比较组间差异表达的基因进行统计分析,默认参数:p-adjust <0.05、|log2FC|≥1,以无菌水互作的燕麦组为对照,使用SeqPrep软件进行测序数据质控。
1.5.2GO功能分析及KEGG富集分析 采用软件Goatools进行GO富集分析[18],分析燕麦与菌株DGL1互作后的差异表达基因功能,利用KEGG[19]数据库对差异基因进行富集分析,并分析基因的表达变化情况,本研究主要分析与促生相关差异基因。
2 结果与分析
2.1 转录组测序数据统计
本研究采用Illumina测序平台完成转录组测序,对获得的15个样品的序列数据进行Q20,Q30质量值进行碱基准确性的评估,共完成15个样品的转录组分析,共获得220.87 Gb Clean Data,15个样品的Clean Data均达到6.24 Gb以上,Q30在95.16%以上,表明数据测序错误率低,测序质量较好。使用SeqPrep软件去除掉接头序列、低质量、含N过多的reads,运用TopHat2软件将15个样本分别与燕麦参考基因组序列进行比对,比对率均大于86.68%,GC含量在54%~57%之间(表1)。
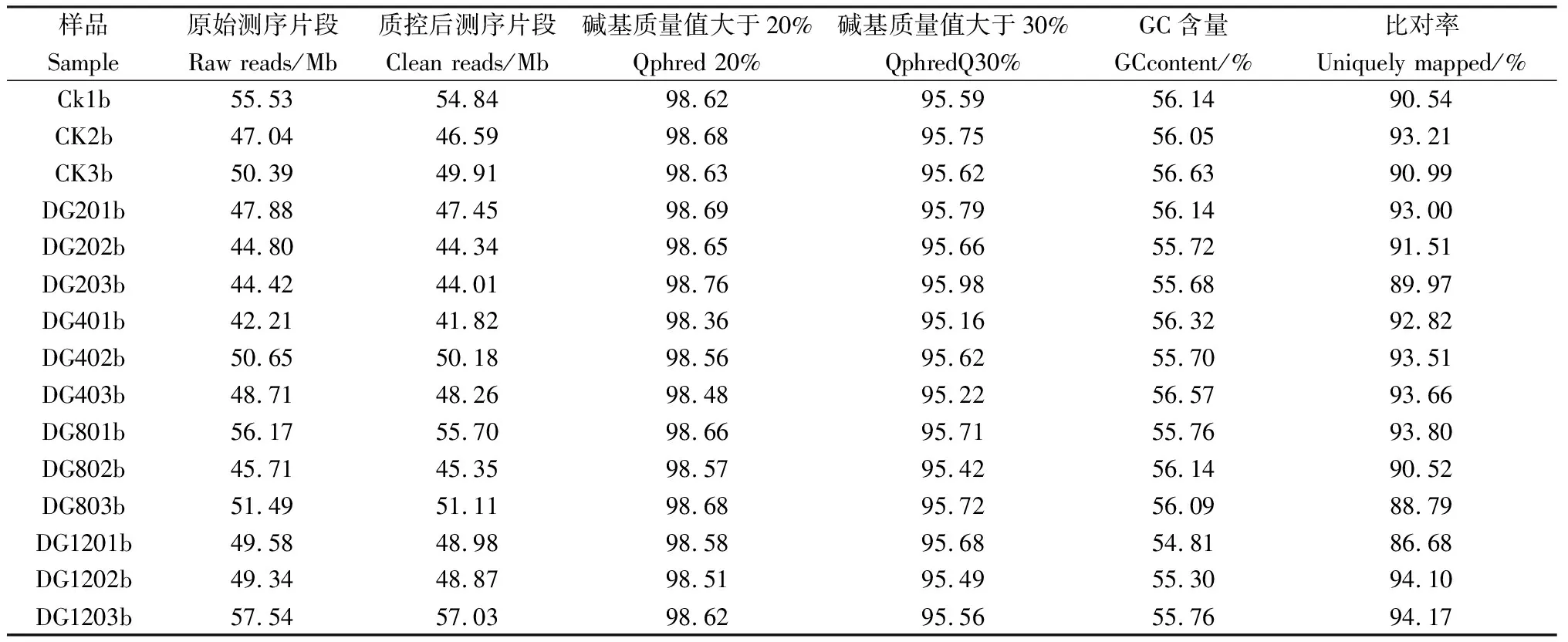
表1 转录组测序数据统计
2.2 差异表达基因分析
将4个处理组b2(2 h互作组)、b4(4 h互作组)、b8(8 h互作组)、b12(12 h互作组)与对照组(CKb)比对。结果表明:随着互作时间的增长,差异表达基因(DEGs)的数量也随之增加,分别在互作2,4,8和12 h时筛选到1 894,6 130,8 033和12 215个差异表达基因。在互作2 h时,619个基因发生上调表达,1 275个基因发生下调表达;在互作4 h时,1 998个基因发生上调表达,4 232个基因发生下调表达;在互作8 h时,3 010个基因发生上调表达,5 023个基因发生下调表达;在互作12 h时,4 678个基因发生上调表达,7 537个基因发生下调表达(图2)。
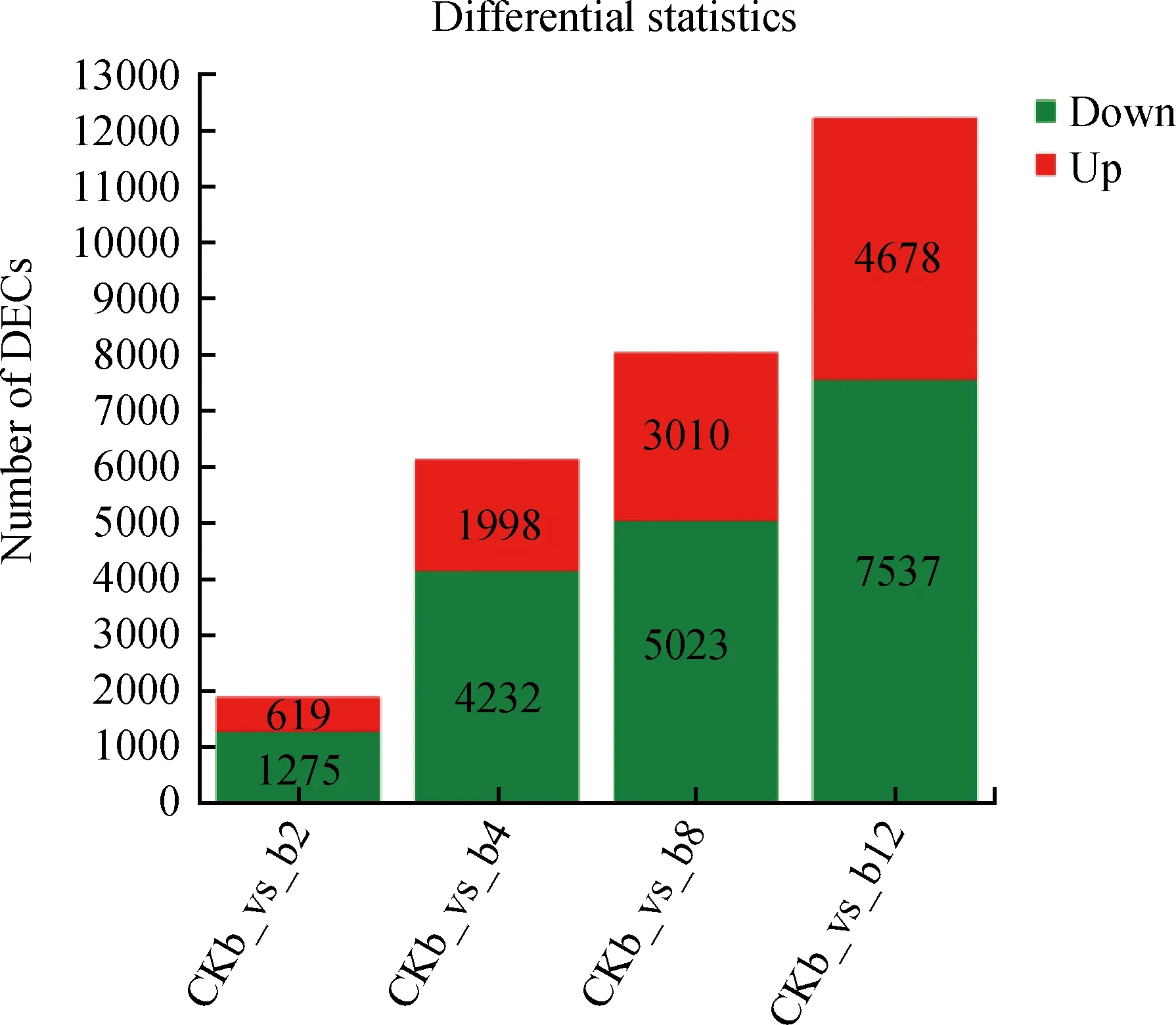
图2 差异表达基因的统计量汇总
通过差异表达维恩图分析发现,燕麦与芽孢杆菌互作4个不同时间处理组共有410个差异表达基因,b2(2 h互作组)与b4(4 h互作组)共有581个差异表达基因(图3),b8(8 h互作组)与b12(12 h互作组)共有3 248个差异表达基因,b4(4 h互作组)与b8(8 h互作组)共有1 843个差异表达基因,b2(2 h互作组)和b8(8 h互作组)共有686个差异表达基因,b2(2 h互作组)和b12(12 h互作组)共有746个差异表达基因(图3)。
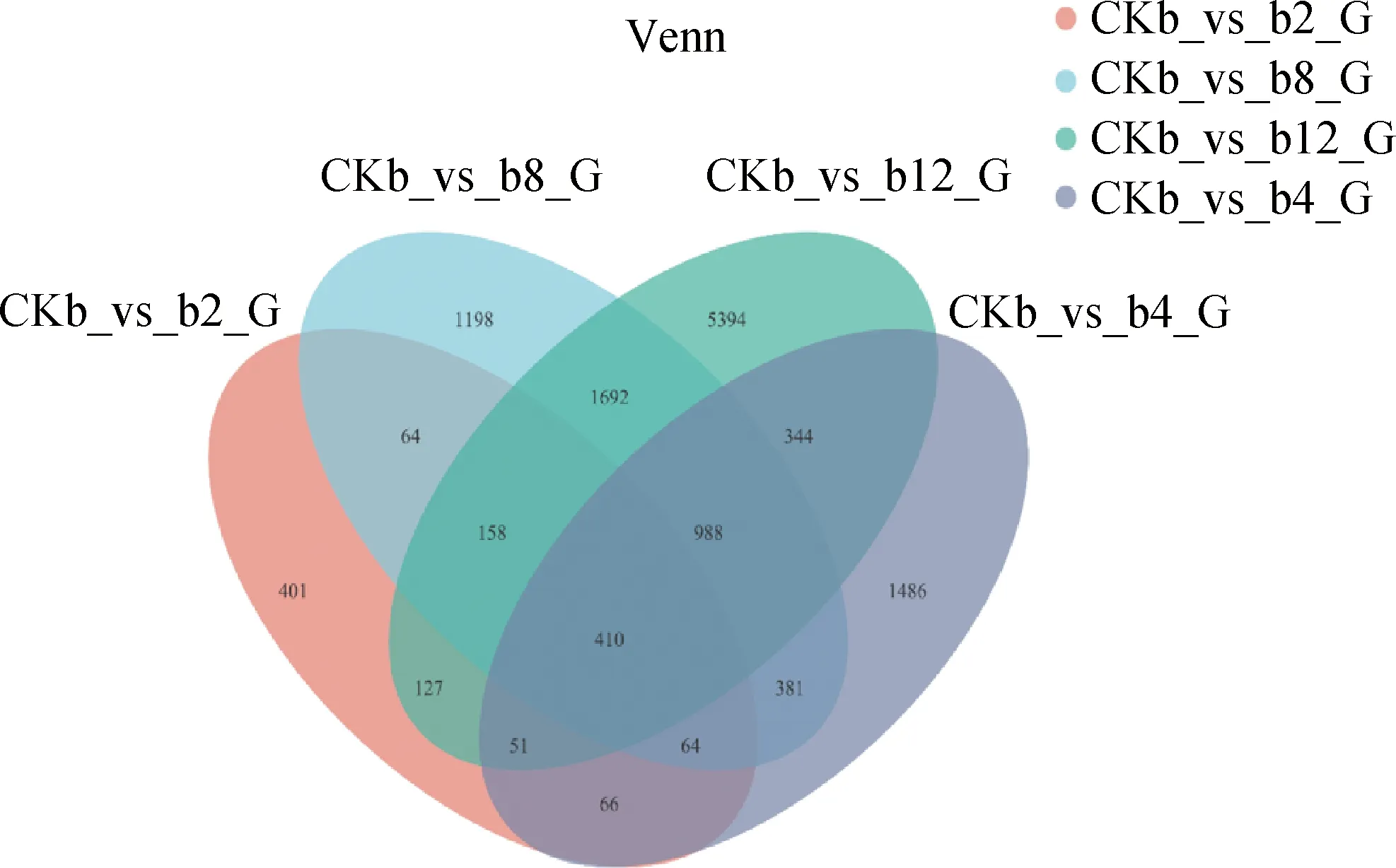
图3 差异表达基因的统计量汇总
2.3 差异表达基因GO富集分析
采用软件Goatools对4个处理组的差异基因进行GO富集,分别阐述互作2 h,4 h,8 h,12 h组中差异表达基因所参与的生物过程、分子功能、细胞组分的信息。
互作2 h,在生物过程中,差异表达基因显著富集在多胺分解代谢过程(GO:0006598)等与植物细胞代谢相关过程中;在细胞成分过程中,基因显著富集在与膜的组成部分(GO:0016021)、胞间连丝(GO:0009506)、细胞连接(GO:0030054)等与植物细胞、细胞膜的形成相关过程中(图4A)。
互作4 h,在生物过程中,基因显著富集在光系统(GO:0006720)、对光刺激反应(GO:0009416)、叶绿素生物合成(GO:0015995)相关过程中;在细胞过程中,基因显著富集在与叶绿体类囊体膜(GO:0009535)、光系统II(GO:0009523)、光系统I(GO:0009522)相关过程中(图4B)。
互作8 h过程中,在生物过程中,基因显著富集在光合作用中光能的捕获(GO:0009765)、植物细胞壁的生物合成(GO:0009832)有关过程中;在细胞过程中,基因显著富集在与光系统I(GO:0009522)、细胞器膜的形成(GO:0031967)相关过程中;在分子功能过程中,基因显著富集在磷酸烯醇式丙酮酸羧化酶活性(GO:0008964)、四吡咯结合(GO:0046906)相关过程(图4C)。
互作12 h,在生物过程中,基因显著富集在硫胺素生物合成(GO:0052837)、多胺分解代谢(GO:0006598)等与植物的生长发育以及胁迫相关的代谢过程;在细胞组分中,基因显著富集在与叶绿体膜间隙形成(GO:0031972)、质体膜间隙(GO:0009529)等与植物细胞、细胞器、膜相关的过程;在分子功能中,基因显著富集在苏氨酸醛缩酶活性(GO:0004793)、苹果酸合酶活性(GO:0004474)等与植物催化活性相关的过程中(图4D)。
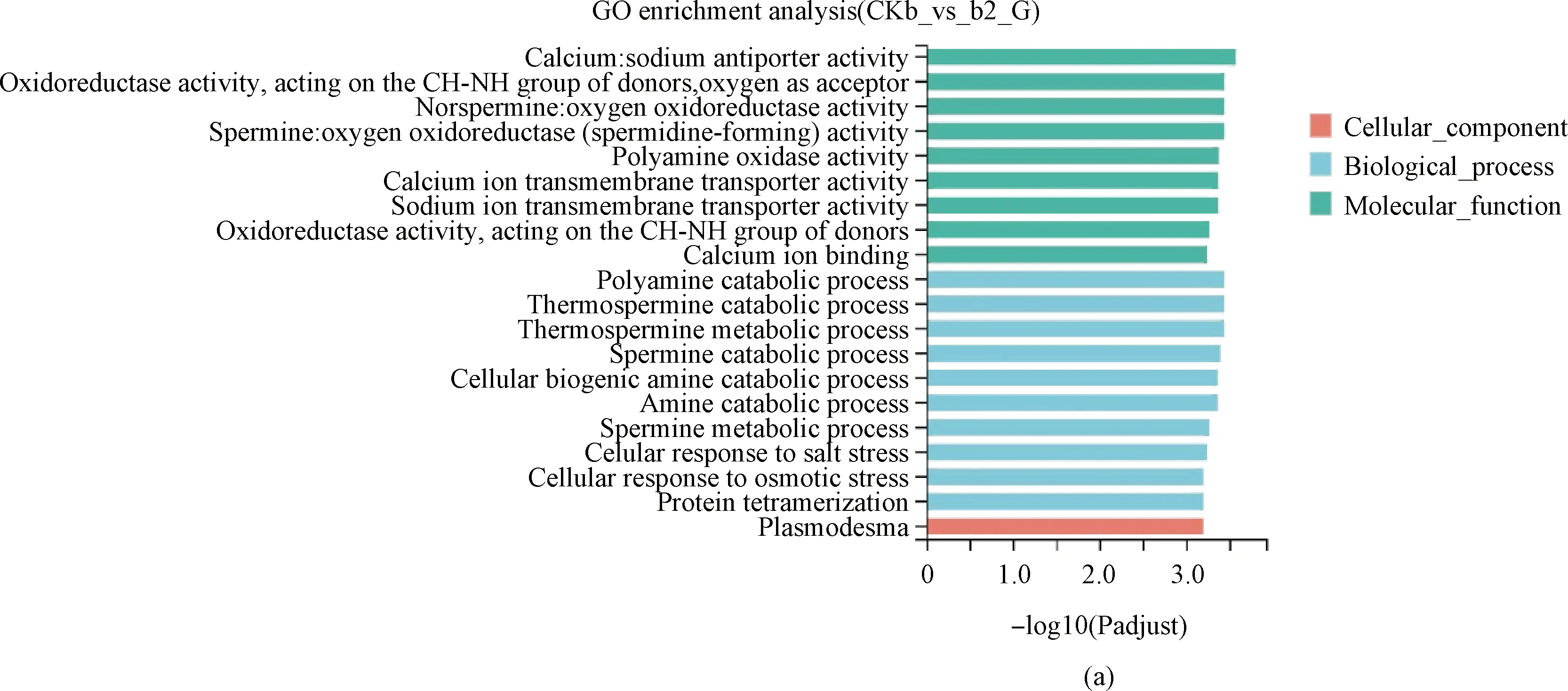
图4 GO功能分类结果图
综上所述,芽孢杆菌DGL1与燕麦根部互作,主要影响燕麦叶部细胞光合器官结构稳定、光合碳同化、氨基酸代谢等途径以此调控燕麦的生长发育[20]。互作2 h时,差异表达基因显著富集在多胺分解代谢过程,多胺与植物生长发育和外界环境的胁迫反应密切相关,推测DGL1可以诱导多胺分解代谢,从而促进燕麦的花器官、根的形成,提高环境胁迫的适应能力;互作4 h,差异基因显著富集在与光合作用相关的代谢通路中,预测DGL1能够促进叶绿体膜等细胞器膜、光合色素、光系统等的合成,加快光合速率,积累光合产物。互作8 h时,差异基因显著富集在与光合作用、细胞壁、细胞器膜的合成过程中,其中磷酸烯醇式丙酮酸羧化酶(Phosphoenolpyruvate carboxyki-nase,PEPC)是参与C4碳同化途径中固定CO2的酶[21],推测DGL1可能通过促进燕麦PEPC酶的活性,促使C3植物燕麦进行一定程度的C4碳同化方式“浓缩”CO2,提高燕麦的光合效率,此外,推测燕麦通过PEPC酶促进草酰乙酸,为保卫细胞提供苹果酸,增强环境胁迫耐受性。互作12 h时,差异基因显著富集在硫胺素、多胺等与生物胁迫及非生物胁迫响应方面,苏氨酸醛缩酶主要参与植物氮素代谢[22],预测DGL1能够通过提高苏氨酸醛缩酶活性,增加蛋白质的含量,提高燕麦产量。
2.4 KEGG富集分析
为了进一步探究菌株DGL1促燕麦生长的分子机制,将4个处理组的差异基因富集到KEGG数据库。分析结果表明:互作2 h,差异基因显著富集在β-丙氨酸代谢途径、精氨酸和脯氨酸代谢途径、甘油脂代谢途径、鞘脂代谢途径、类胡萝卜素生物合成途径等(图5A);互作4 h,差异基因显著富集在光合作用-天线蛋白代谢途径、卟啉和叶绿素代谢途径、β-丙氨酸代谢途径、氰基氨基酸代谢途径、光合生物中的碳同化途径(图5B);互作8 h,差异基因显著富集在光合作用-天线蛋白代谢途径、植物昼夜节律途径、β-丙氨酸代谢途径、精氨酸和脯氨酸代谢途径、硫胺素代谢等(图5C);互作12 h,差异基因显著富集到植物昼夜节律途径、β-丙氨酸代谢途径、硫胺素代谢途径、精氨酸和脯氨酸代谢途径、α-亚麻酸代谢途径等(图5D)。
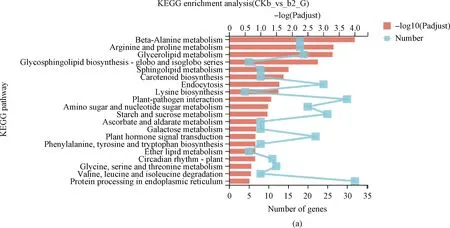
图5 差异基因Pathway富集结果
KEGG富集分析表明,燕麦植株与芽孢杆菌DGL1互作,燕麦叶片组织的代谢通路发生了显著的变化。差异表达基因主要富集在精氨酸、甘油脂等与氨基酸代谢、脂质代谢、多胺合成等相关途径中,推测DGL1促进燕麦蛋白质、脂质的合成及代谢,促进燕麦的生长发育并增强逆境胁迫抗逆性;在互作4 h,8 h时,基因显著富集在光合作用-天线蛋白代谢、卟啉和叶绿素代谢等与光合作用相关的代谢通路中,推测DGL1通过促进燕麦的光合作用,积累碳水化合物、有机酸等光合产物,促进燕麦的生长。
2.5 燕麦应答DGL1相关代谢通路分析
2.5.1植物激素信号通路分析 植物激素作为胞间信号调控植物整个生命周期的生长和发育,并在多种非生物胁迫响应中扮演着重要的角色[23-24]。通过KEGG分析发现,芽孢杆菌DGL1与燕麦互作2 h,4 h,8 h,12 h,诱导了燕麦叶部色氨酸代谢、类胡萝卜素生物合成、光合作用-天线蛋白通路、苯丙氨酸代谢、茉莉酸信号转导途径中相关基因差异表达。筛选出位于细胞膜上的生长素早期响应AUXI蛋白、Aux/IAA蛋白的编码基因在菌株DGL1的作用下,在4个处理组中均上调表达,推测芽孢杆菌DGL1可通过诱导燕麦IAA的形成,促进燕麦细胞的分裂、根的伸长、以及果实的发育。
茉莉酸信号转导途径中的核心受体编码基因COI1在互作2 h上调2.5倍,互作4 h上调3.1倍,互作8 h上调5.7倍,互作12 h上调3.6倍;推测菌株DGL1促进了燕麦的茉莉酸介导的信号转导途径。已有相关报道称,茉莉酸代谢途径能够诱导促分裂原活化蛋白激酶(MAP)级联反应,钙通道以及与乙烯、水杨酸和脱落酸相关激素代谢通路之间的相互作用,提高对逆境的抵抗能力[25]。
类胡萝卜素通过将吸收的光能传递到反应中心叶绿素a分子来促进光合作用,可以扩展光合作用过程中的吸收光谱,同时具有保护强光伤害的功效,通过散热以保护膜和蛋白质免受紫外线的伤害,还为脱落酸的生物合成提供底物[26-28]。通过KEGG富集分析发现,菌株DGL1诱导燕麦ABA受体蛋白编码基因PYL4基因在互作2 h,4 h,8 h,12 h均上调表达,而ABA的合成能够有效地调节植物种类胡萝卜素的合成,据邓昌哲等[29]报道,外源喷施脱落酸,能够使木薯叶片中胡萝卜素含量升高,且涉及类胡萝卜素合成基因上调表达,因此,推测菌株DGL1能够促进燕麦中PYL4基因上调表达,诱导类胡萝卜素代谢通路,从而促进燕麦对光能的转化与吸收,促进光合作用,提高生物量。
2.5.2次生代谢产物通路分析 苯丙烷类生物代谢途径是合成植物生长发育所需要的黄酮、木质素、花青素等次生代谢产物主要方式之一,参与植物细胞分化,色素、细胞壁的形成等生理过程[30]。芽孢杆菌DGL1与燕麦互作,诱导燕麦叶部的苯丙烷生物代谢途径,发现该通路的关键酶苯丙氨酸解氨酶编码基因PAL在4个处理组均上调表达,作为苯丙烷类生物代谢中最重要的酶,其活性的高低能够成为衡量植物抗病性的指标[31],因此,推测菌株DGL1通过促进合成苯丙氨酸解氨酶,从而促进次生代谢物的合成,诱导了燕麦的抗病能力,间接促进了燕麦的生长。
2.5.3光合作用相关通路分析 光合作用-天线蛋白通路途径检测到光系统Ⅰ中的光合蛋白编码基因Lhca1,Lhca2,Lhca3,Lhca4,与光系统Ⅱ中的捕光色素蛋白编码基因Lhcb1,Lhca2,Lhcb3,Lhcb4,Lhcb5,Lhcb6均上调表达。捕光色素蛋白在光能的捕获、传递、分配两个光系统PSI和PSII中的能量平衡,还能维持光合作用主要细胞器膜类囊体膜的结构[32](图6),推测DGL1能够通过诱导燕麦细胞叶绿体中光合链相关组分及光系统中相关蛋白编码基因的表达,稳定和增强叶绿体的结构和功能,促进燕麦光合性能、提高光合速率,促进燕麦生物量的增加。
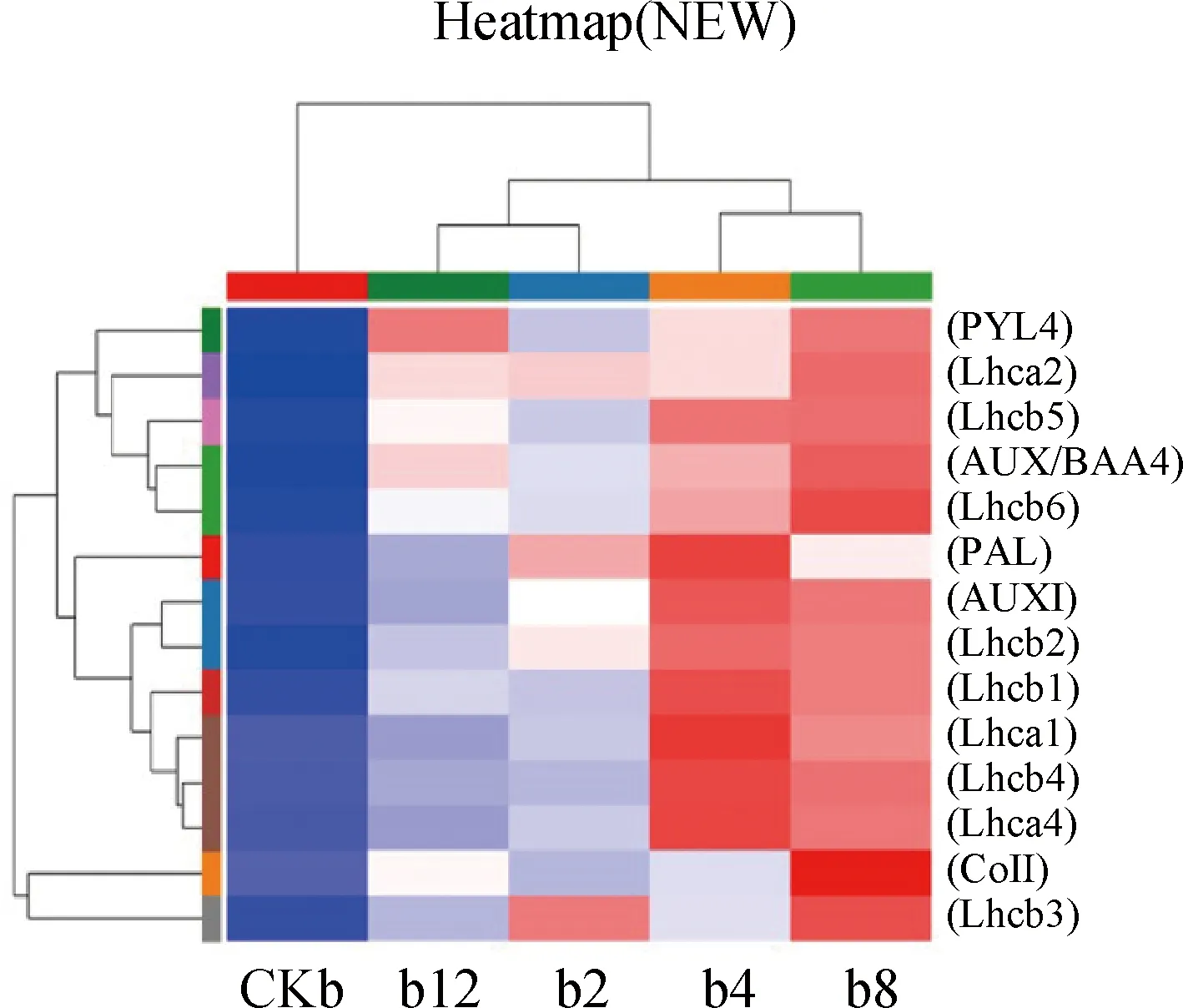
图6 差异基因Pathway富集结果
2.6 促生相关基因分析
芽孢杆菌DGL1与燕麦互作,筛选出多个与促生相关差异表达基因显著上调。
2.6.1信号转导应答 生物及非生物胁迫应答方面,合成丝氨酸/苏氨酸的蛋白磷酸酶基因PP2C在4个处理组均过度表达,在互作2 h上调7.9倍,互作4 h上调4.2倍,互作8 h上调6.8倍,互作12 h上调5.4倍;已有研究表明PP2C蛋白在植物激素ABA信号转导途径具有重要的作用,能够参与调控种子萌发、营养器官发育以及增加植物抗逆性[33]。丝裂原活化蛋白激酶MAPKKK27表现出在互作2 h上调6.6倍,互作4 h上调7.1倍,互作8 h上调6.8倍,互作12 h上调8.3倍,当植物遭受逆境胁迫时,MAPK级联激活调控植物激素信号转导与多种生物、非生物胁迫信号转导途径[34]。编码细胞色素b5蛋白的基因Cytb5在互作2 h上调1.1倍,互作4 h上调1.0倍,互作8 h上调1.6倍,互作12 h上调2.3倍,细胞色素b5蛋白是存在于内质网上的血红素蛋白,能够参与植物细胞中的各种氧化还原反应,通过调节活性氧(ROS)的平衡,提高植物抗逆境胁迫能力[35]。
2.6.2糖类代谢 在糖类代谢方面,调控多糖β-葡聚糖合成的CSLH基因在4个处理组均上调表达,互作2 h上调1.3倍,互作4 h上调1.1倍,互作8 h上调1.8倍,互作12 h上调1.9倍;编码葡萄糖-1-磷酸腺苷酸转移酶的基因glgC互作2 h上调1.8倍,互作4 h上调3.1倍,互作8 h上调3.0倍,互作12 h上调3.1倍(图7)。
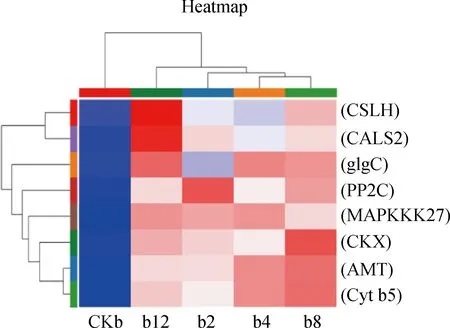
图7 差异基因Pathway富集结果
2.6.3氮素代谢、生长发育 此外,氮素代谢、生长发育相关的基因显著上调。植物氮同化代谢中起到重要作用的编码铵转运蛋白基因AMT在4个处理组中均上调表达,在互作2 h上调1.7倍,互作4 h上调2.2倍,互作8 h上调2.3倍,互作12 h上调1.6倍;位于细胞膜上的铵转运蛋白,能够促进植物对氮素的吸收,提高植物的产量[36]。合成胼胝质合成酶的编码基因CALS2互作2 h上调6.4倍,互作4 h上调6.1倍,互作8 h上调6.4倍,互作12 h上调7.1倍;胼胝合成酶在花粉发育和植物营养物质运输具有重要的作用。
综上所述,芽孢杆菌DGL1诱导了燕麦体内PP2C,MAPKKK27,Cytb5基因的表达,这些蛋白质与植物应答生物、非生物胁迫的耐受反应相关,因此,推测这些蛋白的诱导表达可能会增加燕麦对环境应激的适应能力;基因CSLH,glgC与植物β-葡聚糖、淀粉的合成相关,推测这些多糖编码基因的诱导表达可能会增加燕麦体内储存的养分,为燕麦细胞正常生命活动提供能源;基因AMT与铵转运蛋白的合成相关,因此,AMT的诱导表达可能增加燕麦对氮素的营养吸收,促进蛋白质的合成;胼胝质合成酶是存在植物细胞壁上的特殊成分,能够调节胞间营养物质的输送,DGL1诱导基因CALS2上调表达可能促进了燕麦细胞间的运输能力,为生长发育输送更多的营养物质。
3 讨论
在生长发育过程中,植物根系与根际促生菌PGPR保持着共生关系,植物为微生物生存与繁殖提供营养物质,微生物也通过直接和间接作用促进植物生长,增加植物对生物和非生物胁迫的适应能力[37]。解淀粉芽孢杆菌DGL1能够促进冷地早熟禾(Poacrymophila)、燕麦(Avenasativa)、紫羊茅(Festucarubra)等植物生长,并对多个植物病原真菌具有拮抗活性[38]。为了探究芽孢杆菌DGL1促燕麦生长的分子机制,利用转录组测序技术分析DGL1与燕麦互作后,燕麦叶片对菌株的应答反应。结果表明,互作后差异基因主要富集在影响燕麦细胞生长发育、代谢、光合作用、信号转导、防御反应等相关途径中。
据报道,芽孢杆菌可通过产生植物生长类激素供给宿主来促进植物生长,或者通过调节宿主植物的生长类激素相关基因的表达,调节植物生长发育,诱导植物产生耐逆性[39]。菌株DGL1与燕麦互作后,燕麦色氨酸代谢途径中Aux/IAA编码基因上调表达,Aux/IAA编码的蛋白能够与生长素响应因子ARF结合,调节植物生长素编码基因的表达[40],促进燕麦的生长发育。在茉莉酸信号转导途径中,COI1基因在4个处理组均上调表达,COI1蛋白是茉莉素受体复合体核心成员,具有调控植物发育与诱导植物系统抗性的重要作用,该蛋白的缺失及突变将导致茉莉酸转导途径的受阻[41]。王文静等发现甘蓝型油菜COI1蛋白功能缺失将导致花丝、花药发育受损,出现雄性不育系的表型[42]。菌株DGL1还能够激活燕麦叶部细胞中类胡萝卜素代谢通路,通过合成光和色素促进燕麦对光能的吸收、传递及转化,促进燕麦的光合作用,提高生物量。
通过KEGG富集分析发现,燕麦叶中编码铵转运蛋白的编码基因AMT在4个处理组中均上调表达,可通过表达铵转运蛋白提高燕麦对外界环境中铵态氮的吸收,增加对氮素的利用率;吴贤鑫等发现铵转运蛋白通过促进氮同化和乙烯信号传导来增加水稻对鞘枯病的抵抗力[43],因此推测基因AMT的上调表达,能够促进燕麦对氮素的吸收利用,从而促进生长发育,并提高耐逆境胁迫能力。在次级代谢物的生成途径中,植物根系能够分泌经苯丙烷代谢途径产生的次级代谢物改变土壤微生物的菌群生态,从而促进植物生长,提高植物抵抗不利环境的适应能力[44]。经过菌株DGL1的处理,燕麦叶部的丙氨酸解氨酶编码基因PAL在4个处理组均上调表达,丙氨酸解氨酶是参与木质素合成的关键酶,在防御病害、维持植物直立、输送水分及营养物质具有重要的作用,在植物受到低温胁迫时,丙氨酸解氨酶也能够诱导黄酮类物质的产生以缓解氧化损伤,从而抵御低温伤害[45]。在糖类代谢方面,调控多糖β-葡聚糖合成的CSLH基因、编码葡萄糖-1-磷酸腺苷酸转移酶的基因glgC均上调表达,赵凯琴等报道称glgc基因转入油菜基因组并成功表达,能够促进油菜(Brassicanapus)种子中多糖淀粉的积累[46];崔雪琼等报道重组glgc基因能够增加马铃薯块茎中淀粉的含量[47]。推测菌株DGL1通过诱导CSLH,glgc基因的表达,促进燕麦多糖的合成,积累能源物。
通过转录组测序发现,差异表达基因显著富集在光系统、对光刺激反应、叶绿素生物合成及叶绿体类囊体膜等与光合作用相关的途径中,并发现在光合作用-天线蛋白通路中光系统Ⅰ中的光合蛋白编码基因Lhca1,Lhca2,Lhca3,Lhca4,与光系统Ⅱ中的补光色素蛋白编码基因Lhcb1,Lhca2,Lhcb3,Lhcb4,Lhcb5,Lhcb6均上调表达。因此,推测菌株DGL1能够通过促进燕麦叶部的光合作用实现光能的转换,并将无机物转换为有机物,促进燕麦的生长,这与报道过的芽孢杆菌能够促进植物光合作用的结论一致,史应武等发现接种芽孢杆菌菌剂S-7后,使得甜菜叶片气孔导度、净光合速率、蒸腾速率等指标显著提高[48]。同时有报道称,有益芽孢杆菌能够减少叶绿体中活性氧的含量,增加抗氧化酶类含量,减轻植物在逆境胁迫中产生过多的活性氧造成对叶绿体的损伤,以此保护植物光合效率[49-50]。
4 结论
综上所述,本研究通过RNA-Seq分析芽孢杆菌DGL1与燕麦不同互作时间燕麦叶部差异表达基因及相关代谢通路,探究了芽孢杆菌促进燕麦生长的分子机制,发现芽孢杆菌参与调控与燕麦生长、应答胁迫反应、参与光合作用相关蛋白的编码基因,同时营养吸收的转运蛋白的编码基因在不同互作时间发生了差异表达,推测菌株DGL1对燕麦的促生机制是通过促进燕麦光合作用、激素代谢、次生代谢物合成、氨基酸代谢等多个途径相互协调的结果,本研究为青藏高原极端生境根际促生菌促燕麦生长提供了理论基础。
