基于三种分子标记的栽培金鸡菊遗传多样性及亲缘关系分析
2022-12-01孙浩男周欣莹李明阳刘冬云
孙浩男,李 蓉,王 鑫,周欣莹,李明阳,刘冬云*,李 鹤
(1.河北农业大学园林与旅游学院,河北 保定 071000;2.天津泰达园艺有限公司,天津 300000)
栽培金鸡菊(Coreoopsis)大多为一年生或多年生草本,既包括一些自野外长期引种筛选用于栽培的原生种,也包括许多人工培育的新品种,其舌状花呈现白色、黄色、橙色、红色、粉色、紫色及多种复色和变色,花径大小也有不同,具有极高的观赏价值,深受园林景观和家庭园艺的喜爱,大部分栽培金鸡菊的适应性广泛,一些园艺品种的花期可自5—6月开至霜冻,花期极长[1-2]。菊科(Compositae)金鸡菊属植物大多能够自播,主要原产于北美洲和南美洲,我国主栽剑叶金鸡菊(C.lanceolata)、大花金鸡菊(C.grandiflora)和两色金鸡菊(C.tinctoria)等7种,而国外常栽种的还有耳叶金鸡菊(C.auriculata)、莱氏金鸡菊(C.leavenworthii)等[3-4]。金鸡菊属植物形态变异丰富,同种内存在较多变异类型,仅凭形态特征进行鉴定不易,我国一些学者也常将大花金鸡菊、剑叶金鸡菊和金鬃金鸡菊(C.basalis)混淆,而栽培金鸡菊许多园艺品种之间或和原种相比形态也十分接近,一些品种的遗传背景也不清晰。我国的栽培金鸡菊种质主要依靠国外引进,有部分种质来历不明且存在同名异物的现象,同时,由于目前国际上对于栽培金鸡菊也没有较为系统的遗传图谱,许多种质的亲缘关系也不明确,这些都限制了金鸡菊种质资源的利用[5-7]。
栽培金鸡菊的园艺品种大多通过远缘杂交培育,美国Terra nova公司利用轮叶金鸡菊(C.verticillata)、玫红金鸡菊(C.rosea)等北美原生种培育了一系列远缘杂交品种,如花小且繁密的Pie系列,叶片金黄的Lemonade系列,花色丰富且具有一定耐寒性的Hardy Jewel系列等[8-11],这些也是较早引入我国的无性系品种,常被称为“迷你金鸡菊”。个人育种家Probst引入更多原生种,通过6代以上的远缘杂交培育了众多代表性品种,如花大色艳的Big Bang系列和Little Bang系列、叶型纤细且花色丰富的Satin and Lace系列和PermaThread系列、植株紧凑且耐寒性强的Leading Lady系列、耐热性强且适宜盆栽的Coloropsis系列等[1,12-15],这些也是目前国际上较受欢迎的家庭园艺品种。韩国Uri Seed公司利用伽马射线和除草剂对现有品种未生根的插条进行诱变,筛选出了株型紧凑、适宜盆栽且花色和花型组成奇特的Uri Twinklebells系列、Uri Bloomsation系列和Uri Gardenscape系列等[16-19],主要在日本、韩国和欧洲推广。此外,也有一些其它国家的园艺公司在培育不同大花类型的金鸡菊品种,如美国Ball公司的UpTick系列和‘Superstar’、以色列Danziger公司的Solanna系列,荷兰Dümmen Orange公司的‘Moonswirl’等[20-22]。一些由野外引种选育的栽培种质常用于绿化和药用栽培,它们通常具有自播性和入侵性较强的缺点[23]。近年来,我国盆栽观赏和花境设计对栽培金鸡菊品种的类型和数量要求不断加大,应用较多是籽播类型的大花金鸡菊品种如‘Early Sunrise’,‘Sunkiss’等[24-25],并非上述提到的无性系品种。
DNA是研究植物遗传多样性和亲缘关系常用的标记手段,多年以来,有多种分子标记在金鸡菊属植物上得到应用,如AFLP[26-27],RAPD[28],SSR[29-30],ISSR[31-32]等,此外,还有众多学者利用ITS,matK,psbA-trnH,trnL-F,rbcL等序列研究了金鸡菊属和金鸡菊族(Coreopsideae)内的系统发育和遗传变异[7,33]。ISSR是一种不需要知道DNA序列即可用锚定引物扩增的技术,它同时结合了SSR和RAPD两种分子标记的优点,具有成本低、操作便捷、多态性丰富等特点[34]。核基因ITS、叶绿体基因psbA-trnH和trnL-F序列是常见的DNA条形码,可作为快速和准确识别许多植物物种的有效工具用于园林花卉的栽培溯源和品种分类,其中ITS和ITS2基因更是中国DNA条形码植物工作组认定的种子植物核心条形码[35-38]。本研究以课题组收集自国内外的100份金鸡菊种质为试验材料进行遗传多样性及亲缘关系分析,旨在从分子角度上为栽培金鸡菊在分类鉴定、育种及指纹图谱构建等方面提供更多理论。
1 材料与方法
1.1 试验材料
供试金鸡菊种质共100份,包括14个种,78个园艺品种(24个籽播系,54个无性系),3个课题组自选育种质(1个籽播系,2个无性系),主要收集自世界各国,详情见表1。

表1 供试的100份栽培金鸡菊的信息
1.2 DNA提取、引物筛选和PCR扩增
2020年8月初统一采集植株的成熟叶片,植物基因组DNA试剂盒购买自康为世纪公司,具体步骤按说明书进行。使用Nano Drop 2000微量分光光度计检测提取的金鸡菊DNA浓度,并用1.5%琼脂糖凝胶(1×TAE缓冲液,120 V,0.5 h)对DNA母液上样检测,剔除不合格的样品重新提取直到合格。DNA模板及引物分别稀释至30 ng·μL-1,10 μmol·L-1备用。
ISSR引物参考哥伦比亚大学公布的100条ISSR引物(http://www.biotech.ubc.ca/services/naps/primers/Primers.pdf)进行试验,去除无扩增产物及弥散严重的引物,初步筛选出27条,再以17个亲缘关系较远的籽播型金鸡菊DNA模板进行退火温度试验,复筛出扩增产物稳定、清晰且多态性较好的12条引物。反应体系20 μL:DNA模板2 μL、2×Es Taq Master Mix 10 μL、引物2 μL、ddH2O 6 μL。反应程序:94℃预变性4 min、94℃变性45 s、退火45 s(最适温度)、72℃延伸45 s、32个循环、72℃终延伸10 min。PCR扩增产物使用1.5%琼脂糖凝胶进行检测(1×TAE缓冲液,120 V,40 min),使用FR-2000成像系统进行拍照和记录。
ITS扩增正反向引物分别为ITS5-modified(5′-GGAAGGAGAAGTCGTAACAAGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)[39];psbA-trnH区正反向引物为psbAF(5′-GTTATGCATGAACGTAATGCTC-3′)和trnHR(5′-CGCGCATGGT GGATTCACAATCC-3′)[40];trnL-F区正反向引物为通用引物trnL-c(5′-CGAAATCGGT AG ACGCTACG-3′)和trnL-f(5′-ATTTGAACTGGTGACACGAG-3′)[41]。反应体系25 μL:DNA模板2 μL、2×Es Taq Master Mix 12.5 μL、正反向引物各1 μL、ddH2O 8.5 μL。反应程序:94℃预变形4 min、94℃变性45 s、退火45 s(ITS为64℃、psbA-trnH和trnL-F为60℃)、72℃延伸1 min、35个循环、72℃终延伸10 min。检测合格的PCR产物进行切胶回收和双向测序。
1.3 数据处理
ISSR电泳图上清晰的条带记为“1”,同位置无条带或不易分辨的弱带记为“0”,以此建立原始数据矩阵。计算各引物的扩增总带数(TB)、多态性带数(PB)、多态性百分比(PPB)、利用Popgene 32软件分析群体总观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性指数(H)和Shannon’s信息指数(I)。使用NTsys 2.10e软件,绘制UPMGA聚类图。利用Popgene 32软件计算各材料间的遗传相似系数和遗传距离[42]。
ITS,psbA-trnH和trnL-F基因参照各测序峰图,利用BioEdit软件查看测序结果,并对序列进行人工校对和拼接,修剪两端后计算序列长度、平均序列长度、GC含量、平均GC含量,由于ITS基因测序结果不佳,仅截取ITS2区进行分析。根据NCBI网站上已发表的金鸡菊相关基因进行Blast比对,并利用ClustalX 1.08和MEGA 7.0软件将多序列进行对位排列,对齐和修剪后的序列导入DNAMAN 8和DNASP 5软件突变总数(Total number of mutations,Eta)变异位点(Variable sites,V)、单一突变位点(Singleton sites,S)、简约信息性位点(Parsimony-informative sites,Pi)、核苷酸多样性(Nucleotide diversity,π)、平均核苷酸差异数(k)、单倍型数目(Number of haplotypes,h)、单倍型多样性(Haplotype diversity,Hd)、单倍型多样性方差(Variance of haplotype diversity,Vh)、单倍型多样性标准差(Standard deviation of haplotype diversity,Sh)。最终以同为金鸡菊族的红花大丽花(Dahliacoccinea)为外类群,构建系统发育树[43-45]。
2 结果与分析
2.1 ISSR分析
2.1.1遗传多样性 由图1和表1可知,100个栽培金鸡菊种质的多态性较高,总共扩增出清晰的条带153个,多态性条带(PB)149个,多态性条带百分比(PPB)为97.39%,每条引物扩增的条带8~16个不等,平均扩增条带12.75个,最多的是UBC825,最少的是UBC879。群体总观测等位基因数(Na)为1.973 9,有效等位基因数(Ne)为1.486 5、Nei’s基因多样性指数(H)为0.288 6和Shannon’s信息指数(I)为0.440 6。

表2 12条ISSR引物的基本信息统计

图1 ISSR引物UBC825和UBC868部分PCR扩增电泳图谱
2.1.2聚类分析 遗传相似系数介于0.413 8~1之间,平均为0.713 2,有13组达到了1,如‘中央舞台’和‘精彩表演’、‘南瓜派’和‘樱桃派’等,这些组合或是具有相同的父母本,或是源于同一品种的变异。沙漠金鸡菊与大花金鸡菊‘太阳之子’、‘阳光’、‘圣达菲’的遗传相似系数最小,为0.542 2,亲缘关系最远。遗传距离整体介于0~0.960 0,平均为0.383 5。有13组为0,如Lemonade系列、‘甜梦’与‘天堂之门’等,不能区分部分同系列的种质,也不能用于鉴别变异品种。
根据ISSR的UPGMA聚类图(图2),在L1(D=0.664)处,100个金鸡菊种质可划分为4类,与金鸡菊属下的分组有一定关联,在L2(D=0.664)处可进一步划分为14个小组,可进一步划分种一级的种质。第Ⅰ类由部分金鸡菊组种质组成,分别为金鬃金鸡菊(A)和皇冠金鸡菊(B);第Ⅱ类的89个材料可分为9组,前5组包括两色组的两色金鸡菊(C)和莱氏金鸡菊(G)、歧序组的玫红金鸡菊(D),还有不可思议系列、Coloropsis系列、Lemonade系列、Pie系列、Uri Dream系列等品种,F组包括了‘月光’和所有轮叶金鸡菊杂交品种,第Ⅱ类后4组包括除第Ⅰ类其余金鸡菊组的种质和Big Bang系列、Little Bang系列、Solanna系列、‘雪莓’等。第Ⅲ类包括仅包括轮叶组的大叶金鸡菊、三叶金鸡菊、掌叶金鸡菊、轮叶金鸡菊;第Ⅳ类为异果组的沙漠金鸡菊。
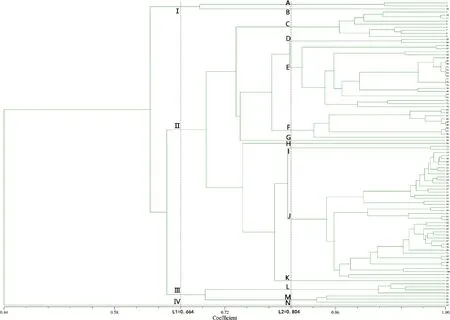
图2 基于ISSR的100份金鸡菊种质资源的UPGMA聚类图
2.2 psbA-trnH,trnL-F及ITS2序列分析
2.2.1序列及多态性分析 根据部分电泳结果(图3),psbA-trnH序列大小在250~500 bp之间,其中轮叶组的轮叶金鸡菊-德国、大叶金鸡菊、掌叶金鸡菊、三叶金鸡菊和异果组的沙漠金鸡菊的序列明显较长;trnL-F和ITS序列大小在700~800 bp之间,序列之间大小差异不明显。部分ITS序列出现非特异性条带,为通用性引物导致,但都有一条明亮主带,切取这部分条带进行回收测序。测序结果中,ITS序列GC碱基富集,部分位点出现套峰,部分片段底峰较高,因此仅截取测序结果较好的ITS2区进行序列处理和分析。将psbA-trnH,trnL-F及ITS2共300个序列上传至NCBI获得登录号(表1):ON887337~ON887436,ON887437~ON887536,ON887537~ON887636。
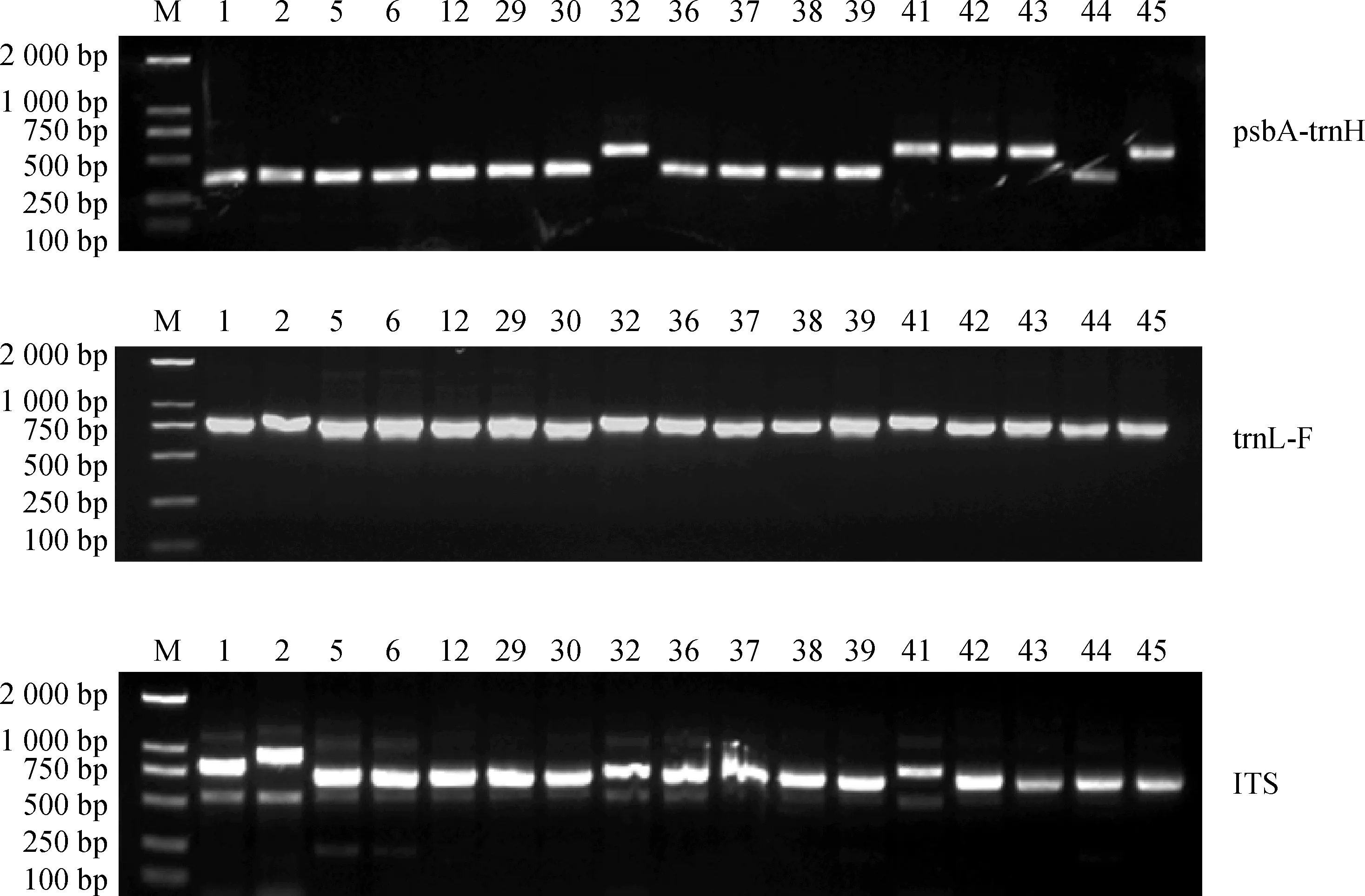
图3 部分ITS、psbA-trnH和trnL-F序列电泳图谱
序列基本信息见表3,三个序列按序列平均大小排列为trnL-F>psbA-trnH>ITS2,其中,ITS2和trnL-F序列大小较为稳定,ITS2的GC含量存在较大的变异范围,两个叶绿体序列的GC含量较为稳定。ITS2的平均GC含量最大,为57.65%,而psbA-trnH最低,为28.51%。在psbA-trnH序列中,所有材料都存在一段长短不一的poly-T结构,共有8个材料在此结构后与其余92个材料的序列同源性较低,有大量碱基无法对齐,不仅包括电泳结果中pbsA-trnH序列较长的5个材料外,还包括3个轮叶金鸡菊的园艺品种,但后三者部分序列片段丢失。在后续分析中修剪掉无法对齐的碱基做后续分析,ITS2在突变总数(Eta)、变异位点(V)、单一突变位点(S)等8个指标上远大于两个叶绿体序列,变异最为丰富,但在单倍型多样性方差(Vh)和单倍型多样性标准差(Sh)上低于二者。trnL-F在突变总数(Eta)、变异位点(V)、单一突变位点(S)上虽略大于psbA-trnH,但在简约信息性位点(Pi)、核苷酸多样性(π)等指标上更低。

表3 供试金鸡菊的ITS,psbA-trnH和trnL-F序列分析
2.2.2基于psbA-trnH及trnL-F序列的母系溯源 将psbA-trnH和trnL-F两段序列进行拼接,合并后的序列在多样性分析中共定义了9个单倍型(表1,表3,表4,图4),其中由野外引种长期筛选栽培的种质在单倍型上能够完成部分种一级或组一级的划分:异果组的沙漠金鸡菊属于HA9型;轮叶组的种质属于HA5型;两色组种质中,两色金鸡菊及其品种属于HA2型,莱氏金鸡菊属于HA8型。金鸡菊组的种质分布在HA1,HA3,HA4和HA6型中:皇冠金鸡菊为HA6型;金鬃金鸡菊、剑叶金鸡菊为HA1型;大花金鸡菊、耳叶金鸡菊‘金色精灵’和短毛金鸡菊‘阳光超人’为HA3型,但短毛金鸡菊‘虹光’和歧序组的玫红金鸡菊同为HA7型。大花金鸡菊和剑叶金鸡菊的单倍型有区别,在园艺品种上也存在一定差异,大花金鸡菊‘和谐’和大花金鸡菊‘旭日’与剑叶金鸡菊同为HA1型,而剑叶金鸡菊‘斯丹泰勒’、‘索拉纳金杯’和其余大花金鸡菊品种单独成HA4型。剩余的种质仅分布在HA2型和HA7型中,除2个不可思议系列的种质属于籽播系品种,其余全部为无性系品种,与两色金鸡菊及玫红金鸡菊具有较强的母系遗传关系。

表4 基于psbA-trnH和trnL-F序列的9个金鸡菊单倍型类型及差异位点
以红花大丽花(psbA-trnH:AY215528;trnL-F:MT214749,图中为101)为外类群构建的MP(Maximum Parsimony)系统发育树,将一些单位型进行了整合(图4,图5)。MP系统发育树显示大多无性系的品种与两色组和歧序组有较强的母系遗传关系,在组一级上的母系溯源更加明确。Ⅲ类和Ⅵ类之间存在7~9个碱基的差异,而母系与金鸡菊组相关的Ⅳ类和Ⅴ类之间存在7~8个碱基的差异,可见金鸡菊组相关的栽培金鸡菊母系变异更为丰富。轮叶组(Ⅰ类)和异果组(Ⅱ类)的8个材料较早独立于其它4类,它们也是测序时发现psbA-trnH序列poly-T结构后同源性较低的8个材料。综合单倍型网络图和MP系统发育树来看,异果组的沙漠金鸡菊与其余栽培金鸡菊的遗传距离和亲缘关系最远,其次为轮叶组的大叶金鸡菊、掌叶金鸡菊、三叶金鸡菊和轮叶金鸡菊(包括园艺品种)。舌状花主花色为红色、橙色、粉色、紫色、白色和浅黄色的种质在母系溯源中全部与两色组与歧序组有关。通常被当做轮叶金鸡菊和玫红金鸡菊杂交得来的Uri Dream系列、‘中央舞台’、‘精彩表演’等园艺品种的母系为两色组,但舌状花不具两色金鸡菊的花眼。通常被当做轮叶金鸡菊品种的‘月光’,‘焦糖布丁’和‘甜蜜果酱’母系全部为歧序组,涉及到远缘杂交。
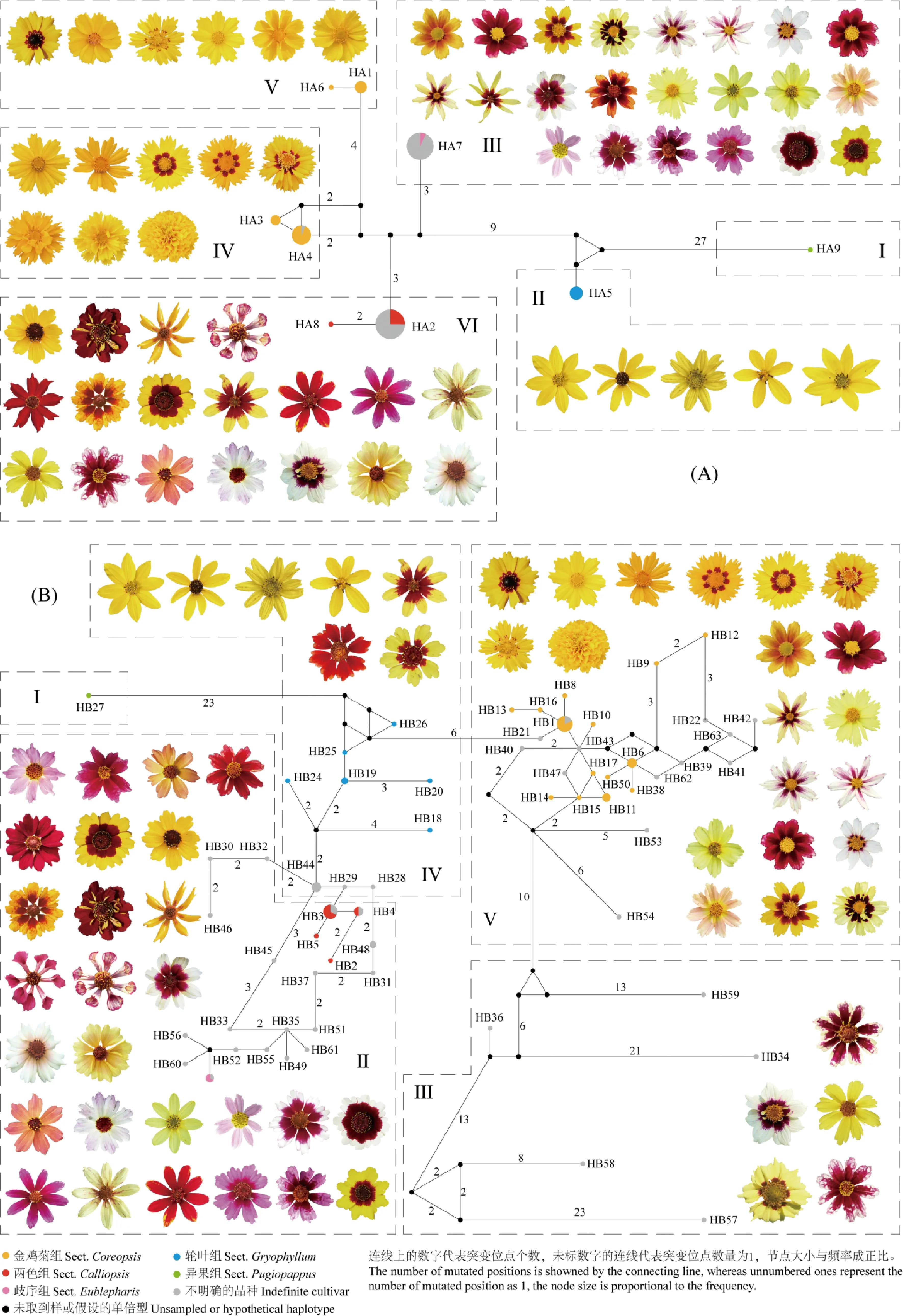
图4 栽培金鸡菊在叶绿体基因psbA-trnH+trnL-F(A),核基因ITS2(B)上的单倍型网络图及相关种质的头状花形态

图5 基于psbA-trnH和trnL-F序列分析的100个金鸡菊种质资源的MP系统发育树
2.2.3基于ITS2序列的聚类分析 100个ITS2序列中共出现63个单倍型(表1,表3,图4),大多单倍型仅包括1个品种,一些在ISSR标记和叶绿体标记中较难区分的种质得到了分型,如‘焦糖布丁’、‘甜蜜果酱’、Lemonade系列和Pie系列等,有部分品种共享1个单倍型,如轮叶金鸡菊‘黄金雨’和轮叶金鸡菊‘大花’同为HB19,‘美国梦’和‘天堂之门’同为HB29,‘日落梦想’和‘粉色梦想’同为HB48等,可能为同一种质或发生突变的种质。部分源于同一杂交组合的品种得到了区分,如‘秋之诗韵’和‘雪莓’,但无法区分‘中央舞台’和‘精彩表演’。除大花金鸡菊‘叠日’和大花金鸡菊‘欢歌’外,其它源于美国的大花金鸡菊品种均为HB1型,未能得到区分,而除大花金鸡菊‘太阳之子’外,所有源于德国的大花金鸡菊品种均能通过单倍型分析得到区分。
通过K2P模型计算ITS2序列的遗传距离介于0~0.553 3之间,平均遗传距离为0.080,有122个组合的遗传距离为0,最大的为沙漠金鸡菊和‘雪莓’。以红花大丽花(ITS2:AY561279;图中为101)为外类群构建NJ(Neighbor-Joining)系统发育树(图6),100个金鸡菊种质可划分为5类:异果组的沙漠金鸡菊最早独立为第Ⅰ类,与ISSR和叶绿体母系溯源的结果一致;第Ⅲ类包括‘秋之诗韵’、‘雪莓’、‘蔓越莓沙冰’以及Hardy Jewel系列中的‘红宝石霜’和‘黄水晶’。第Ⅴ类的44个种质主要与金鸡菊组有关,包括所有金鸡菊组的种质‘焦糖布丁’、‘甜蜜果酱’以及除Ⅲ类外所有金鸡菊组远缘杂交的品种。第Ⅱ类主要与两色组与歧序组有关,包括两色金鸡菊、莱氏金鸡菊、玫红金鸡菊、‘甜梦’、‘天堂之门’、‘月光’、‘精彩表演’、‘中央舞台’等36个种质;第Ⅳ类包括所有轮叶组的种质、还有Pie系列中的‘菠萝派’和‘樱桃派’、Satin and Lace系列等14个种质。结合图4和图6可知,第Ⅱ类、Ⅳ类和Ⅴ类中相邻的单倍型大多仅有1~3个碱基的差异,‘焦糖布丁’(HB53)和‘甜蜜果酱’(HB54)不仅未与轮叶组划为一组,也未与母系的歧序组划为一组,而是由某一假设的(或未取到样)与金鸡菊组相关紧密的单倍型突变5~6个碱基得来,此外,此单倍型也经过较为复杂的突变形成了HB34,HB36,HB57,HB58和HB59,这5个单倍型单独聚为Ⅲ类,其头状花形态虽与Ⅴ类较为接近,但在NJ系统发育树中较Ⅳ类更早地独立出Ⅴ类,存在更远的遗传距离和亲缘关系。

图6 基于ITS2序列分析的100个金鸡菊种质资源的NJ系统发育树
3 讨论
前人的研究表明,金鸡菊属植物大多自交不亲和[5-6,46-47],高异交率是金鸡菊产生高遗传多样性的主要原因[23,48],野生金鸡菊的遗传多样性主要集中在种群内部,而种群间的较少[26-28]。栽培植物在驯化过程中使用的原生种个体通常有限,人们对良种的选择、长期的无性繁殖等,均会导致栽培植物的遗传多样性降低[43,49]。栽培金鸡菊具有复杂的遗传背景,在形态学(未发表)和细胞学[2]上也具有较高的遗传多样性,但基于ISSR的分子标记发现,100个栽培金鸡菊种质的平均Nei’s基因多样性指数(H)为0.288 6,而前人报道同属的两色金鸡菊栽培种质为0.149 8[32],均低于牡丹(Paeoniasuffruticosa)0.308 5[50]、矮牵牛(Petuniahybrida)0.374 4[51]、萱草(Hemerocallishybrids)0.455 7[52]等常见观赏花卉,这可能与其籽播系种质来源较为单一,且一些无性系品种的杂交路线趋同有关,但也可能是分子标记类型、所选引物或统计方法不同等导致的。
在研究被子植物系统发育时,由于谱系不完全分选和叶绿体捕获,母系遗传的叶绿体基因并不总是与核基因的系统发育相对应,后者发生在叶绿体基因的定向基因流下,而前者是一个随机的过程[53-55]。群体之间通过杂交和反复回交,部分基因会在不同亲本中交换发生基因渐渗,而原生叶绿体基因组很容易被外来叶绿体基因组所取代发生叶绿体捕获,此类种间杂交后代的细胞核和细胞质通常来自不同的物种[56]。本研究发现,基于叶绿体基因psbA-trnH和trnL-F的系统发育树与基于核基因ITS2的系统发育树存在较大分歧,舌状花主花色为红色、橙色、粉色、紫色、白色和浅黄色的种质在母系溯源中全部与两色组与歧序组有关,而在ITS2的系统发育树和ISSR聚类中,此类种质部分与轮叶组和金鸡菊组有着更近的亲缘关系,可能是在杂交和回交过程中发生了叶绿体捕获。在ITS2系统发育树和单倍型网络图中发现,‘黄水晶’、‘红宝石霜’、‘秋之诗韵’、‘雪莓’和‘蔓越莓沙冰’这五个材料从Ⅴ类中独立成Ⅲ类,它们所对应的HB34,HB36Z,HB57,HB58和HB59单倍型彼此之间具有较大的碱基差异,结合母系溯源和已发表的品种专利来看[10-11,57-60],这五个品种都是以金鸡菊组为父本,两色组为母系的种质。通常被认为是轮叶金鸡菊的‘焦糖布丁’和‘甜蜜果酱’[61-62]归为Ⅴ类,其亲本包含金鸡菊组,二者所对应的HB53和HB54单倍型与相邻单倍型之间有5~6个碱基差异,而其余相邻单倍型之间仅有1~3个碱基差异,这可能是由于二者引入了轮叶组的基因。ITS2中的Ⅲ类和Ⅴ类种质的育种目标多为培育大花型和色彩丰富的多年生金鸡菊品种,它们通过引入两色组和歧序组改良花色,在此基础上与金鸡菊组杂交,因此整体上与金鸡菊组的亲缘关系更近,它们在花部性状上也更偏向于金鸡菊组,但也有部分性状介于Ⅴ类和Ⅱ类之间,如头状花直径大小连续、花盘上可同时出现4裂和5裂的管状花(金鸡菊组和轮叶组为5裂,两色组和歧序组为4裂)等,这些性状可能为双亲遗传。
三种标记均一致显示沙漠金鸡菊与其它栽培金鸡菊种质的亲缘关系较远,部分学者通过形态学和分子证据也认为沙漠金鸡菊和木本金鸡菊(C.gigantea)等与其余金鸡菊属植物相比亲缘关系较远,可划分为Leptosyne属[63],结合本次试验结果,沙漠金鸡菊可作为一个栽培金鸡菊分类的外类群。ITS2序列和ISSR标记在对栽培金鸡菊的划分也有所差异,与EST-SSR标记[30]的结果相比也存在不同。ISSR标记显示,歧序组、两色组、‘月光’和所有轮叶金鸡菊杂交品种划为一类,ITS2将其中一部分轮叶金鸡菊杂交品种与轮叶组划为一类,而EST-SSR标记将‘月光’、轮叶金鸡菊和轮叶金鸡菊杂交品种聚一类,最后才与其他轮叶组种质聚为一类。Pie系列三个品种在ISSR标记中与两色组和歧序组聚为一类,和EST-SSR结果相同[30],但‘菠萝派’和‘樱桃派’在ITS2标记中与轮叶组聚为一类,推测它们的亲本‘Little Penny’(‘RP1’)和‘Rum Punch’由轮叶组、两色组和歧序组的种质共同参与了杂交[9,64-65]。
叶绿体标记可为一些具有特殊花色的类群在母系溯源上提供有力证据[66]。舌状花是栽培金鸡菊的主要观赏部位,丰富的舌状花主花色是栽培金鸡菊远缘杂交育种所期望的目标性状之一,在叶绿体溯源研究中发现,两色组和歧序组是栽培金鸡菊重要的母系材料,与二者相关的种质占整体的66%,具有特殊舌状花主花色的种质在母系溯源中全部与两色组与歧序组相关,本研究也首次通过分子手段证明了‘月光’和短毛金鸡菊‘虹光’为远缘杂交品种,其母系为歧序组,支持了Coombs的推测[1]。孙浩男等[2]发现,栽培金鸡菊中可能存在大量非整倍体和异源多倍体,本试验发现,以x=12,13,14呈整数倍叠加的非整倍体和异源多倍体品种在母系溯源中主要与两色组相关,这些品种可能也包括歧序组基因,但歧序组不为母系,而不以整数倍叠加的非整倍体品种母系全部为歧序组,这可能与单倍型HA7的‘甜梦’与‘天堂之门’参与杂交有关[2]。
本次试验还欠缺个别表现优异,但目前并未在商业上得到大面积推广的种质,如沼生金鸡菊(C.palustris)、长叶金鸡菊(C.gladiata)及全缘叶金鸡菊(C.integrifolia)等[1],它们大多源于歧序组,但鲜有用于杂交育种的报道,目前已有报道确认的栽培金鸡菊育种亲本仅有10余种[1-2,30,67],整体上看,金鸡菊属内还有相当一部分物种有待开发,有关栽培金鸡菊舌状花花色、头状花类型等遗传规律也有待进一步研究。
4 结论
三种分子标记均显示栽培金鸡菊具有一定的遗传多样性,沙漠金鸡菊与常见的栽培金鸡菊种质亲缘关系较远。本研究筛选出了12条适用于栽培金鸡菊的ISSR引物,在遗传距离为0.664时可将整体分为4类。栽培金鸡菊ITS2,psbA-trnH和trnL-F序列大小分别为171~175 bp,252~426 bp和698~703 bp,ITS2和trnL-F序列长度较为稳定,ITS2序列较psbA-trnH和trnL-F序列具有更丰富的变异。叶绿体基因psbA-trnH和trnL-F母系溯源显示,两色组及歧序组是栽培金鸡菊重要的母系资源,有66%的种质与两者相关,序列合并后定义了9个单倍型,HA2和HA7单倍型的种质适宜作为母系培育具有特殊舌状花花色的新品种,同时,本研究首次从分子角度确认了‘月光’及短毛金鸡菊‘虹光’等为歧序组的远缘杂交品种。ITS2序列可将整体分为5类,共定义了63个单倍型,ITS2有望作为栽培金鸡菊品种鉴定的DNA条形码。本研究为栽培金鸡菊在分类鉴定、亲本溯源、杂交育种及指纹图谱的构建等方面提供了有力的分子证据。
