萝卜硫素下调结直肠癌中eIF4F翻译起始复合物的表达
2022-11-03周大臣喻宗繁
邢 运,周大臣,贺 良,崔 笑,喻宗繁
结直肠癌(colorectal cancer, CRC)是指结肠和直肠黏膜上皮和腺体发生的恶性肿瘤。2018年的全球癌症统计结果表明,CRC总发病人数185万,占比10.2%,居全球癌症发病率第3位。死亡人数88.1万(9.2%),居第2位[1]。在细胞翻译过程的起始阶段,真核细胞启动因子4F(eIF4F)翻译起始-复合物作为限速步骤发挥着重要作用[2]。萝卜硫素(sulforaphane,SFN)是一种异硫氰酸酯,天然存在于大多数十字花科蔬菜中。1992年首次从西兰花中提取,并定义为II相解毒酶诱导剂[3]。SFN已被证实具有多种抗癌特性,特别是对CRC[4]。该研究从CRC患者组织标本中检测eIF4F翻译起始复合物相关蛋白的表达,进一步探讨SFN影响CRC细胞增殖的具体分子生物学机制,为SFN治疗CRC提供理论依据。
1 材料与方法
1.1 材料
1.1.1组织标本 收集12例安徽医科大学第二附属医院普外科CRC患者的组织标本,分别收取肿瘤组织与癌旁组织,放入装有RNA组织保存液的标本管中,并且所有标本在收集和运输过程中均置于冰上,并于-80 ℃冰箱中长期保存,将收集到的12例组织样本制作为组织芯片。
1.1.2细胞与动物 原代CRC细胞系HT-29购于上海生命科学院细胞典藏库。健康裸鼠,4~6周龄,18~22 g,购自江苏南京集萃药康生物有限公司,动物生产许可证号:SCKK(苏)2018-0008。饲养于20~25 ℃、40%~60%湿度、照明与黑暗各12 h交替环境中,自由进水、饮食,1周后行后续实验。
1.1.3试剂和仪器 SFN、MTT购自美国Sigma公司;胎牛血清购自美国Gibco公司;青霉素/链霉素、BCA试剂盒和4%多聚甲醛购自上海碧云天生物技术有限公司;DMEM购自美国Hycolne公司;0.25%胰酶购自美国Wisen公司;β-actin和eIF4F复合物相关抗体购自美国Cell Signaling Technology公司;山羊抗兔或抗鼠二抗购自北京中杉金桥有限公司;二甲基亚砜(DMSO)和0.1%结晶紫购自北京索莱宝科技有限公司。细胞培养箱购自美国 Thermo Fisher公司,荧光倒置显微镜购自日本OLYMPUS公司,酶标仪购自上海科华生物工程股份有限公司。
1.2 方法
1.2.1细胞培养 原代CRC细胞系HT-29细胞,DMEM高糖培养基+10%血清+双抗(青霉素、链霉素)中培养,细胞培养箱培养条件为:5%CO2,37 ℃。每3 d换液1次,每5 d传代1次,传代次数不超过10代,1 500 r/min离心5 min,取生长状态良好的处于对数生长期的HT-29细胞进行实验。
1.2.2细胞活力测定(MTT) 将细胞接种于密度为3×103个/孔的96孔板中。二甲基亚砜(DMSO,对照组)和不同浓度的SFN处理48 h。MTT比色法测定细胞活力。在每个时间点加入5 mg/ml 的MTT溶液10 μl,37 ℃孵育4 h。移除培养基并添加200 μl异丙醇以溶解甲醛沉淀物。在570 nm处测量细胞匀浆的光密度,620 nm作为参比滤光片。DMSO作为载体对照,对细胞活力无影响。
1.2.3Western blot实验 处理后的HT-29细胞PBS洗2次,加入蛋白裂解液(RIPA ∶蛋白酶抑制剂 ∶磷酸酶抑制剂比例为1 ∶100 ∶100),待细胞裂解完全后吸取悬液,4 ℃、13 200 r/min离心30 min,吸取蛋白上清液,储存于-80 ℃冰箱,用BCA法测蛋白浓度并进行蛋白质定量,加上样缓冲液煮沸10 min变性,以蛋白质30 μg进行SDS-PACE电泳。转膜后,用含5% BSA的TBST室温慢摇封闭1.5 h,分别加入β-actin和相应一抗(1 ∶1 000),4 ℃孵育过夜,加入相应的辣根过氧化物酶标记的羊抗鼠/兔二抗孵育1 h,洗膜后用显影仪检测蛋白显影。
1.2.4动物实验(裸鼠成瘤) 12只健康适龄裸鼠随机均分为两组,将0.1 ml(细胞数约1×106)处于对数生长期的HT-29细胞沿皮下缓慢注入裸鼠腋窝。3 d后,实验组每天腹腔注射溶解于DMSO的SFN(0.5 mg/kg),对照组每天腹腔注射等浓度的溶剂DMSO(0.5 mg/kg),共注射14 d。14 d后处死裸鼠,取出肿瘤,测量体积大小,将肿瘤标本制成石蜡切片进行苏木精-伊红(HE)染色和免疫组织化学染色。
1.2.5HE染色法 石蜡切片置于65 ℃恒温箱中烘焙60 min,于二甲苯中脱蜡,依次在无水乙醇、95%、70%、50%乙醇溶液中脱蜡处理。将切片依次放入苏木精、伊红染色槽中,用自来水洗涤,再置于50%、70%、95%乙醇溶液、无水乙醇中脱水处理,干燥后用中性树脂密封,镜下观察细胞结构与形态。
1.2.6免疫组织化学染色法 石蜡切片置于65 ℃恒温箱中烘焙60 min,于二甲苯中脱蜡,依次在无水乙醇、95%、70%、50%乙醇溶液中脱蜡处理。再放入枸橼酸钠溶液中高温下抗原修复10 min。用纯水冲洗2次,每次5 min,油性笔画圈,10%过氧化氢去离子水 ∶甲醇(1 ∶9) 孵育10 min,以灭活内源性过氧化物酶活性,TBST冲洗3次,每次5 min,破膜液(含0.25% Triton X-100的TBS) 10 min,TBST冲洗,滴加10%山羊血清封闭液,室温孵育2 h,滴加一抗,4 ℃过夜。回收一抗,TBST冲洗,滴加反应增强液,清洗,滴加相应辣根过氧化物酶标记的二抗,纯水冲洗,DAB显色后自来水冲洗终止反应,苏木精染核20 s,纯水冲洗。再置于50%、70%、95%乙醇溶液、无水乙醇中脱水处理、固定,中性树胶封片后镜下观察相关蛋白表达情况。
1.3 统计学处理使用GraphPad Prism 9.0软件对实验数据进行统计学分析。细胞相关实验均分别重复3次以上。用ImageJ软件测量蛋白灰度值以及免疫组化光密度值,并对光密度(optical density,OD)值、裸鼠成瘤中的肿瘤体积结果进行t检验分析,P<0.05表示差异有统计学意义。
2 结果
2.1 CRC患者标本中eIF4F复合物相关蛋白在肿瘤组织中的表达高于癌旁组织CRC患者标本的免疫组化染色结果显示,eIF4F复合物相关蛋白在肿瘤组织中的表达明显高于癌旁组织(图1A)。OD值分析显示:肿瘤组织4EBP1(t=4.235,P<0.001)、P-4EBP1(t=2.974,P<0.01)、eIF4E(t=3.820,P<0.001)、P-eIF4E(t=3.308,P<0.01)、eIF4G(t=4.252,P<0.001)表达均高于癌旁组织,差异有统计学意义(图1B)。

图1 CRC患者肿瘤组织与癌旁组织中eIF4F复合物相关蛋白的表达差异 ×200A:肿瘤组织与癌旁组织免疫组化染色结果;B:各组OD值差异分析;与癌旁组织比较:**P<0.01,***P<0.001
2.2 SFN可抑制HT-29细胞的增殖用MTT比色法检测SFN作用48 h后HT-29细胞存活率的变化,结果显示:SFN对HT-29细胞活性有抑制作用,并且呈剂量依赖性。SFN对HT-29细胞的半抑制浓度(half maximal inhibitory concentration, IC50)为15.85 μmol/L(图2A)。裸鼠成瘤实验结果显示,实验组肿瘤体积低于对照组,差异有统计学意义(t=3.645,P<0.05)(图2B)。HE染色结果显示,与对照组比较,实验组组织细胞内空泡更多(图2C)。
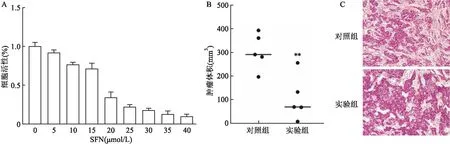
图2 SFN抑制HT-29细胞的增殖 ×200A:不同浓度SFN处理HT-29细胞48 h后的细胞活性;B:裸鼠皮下移植瘤的体积比较;C:裸鼠皮下移植瘤的HE染色;与对照组比较:**P<0.01
2.3 SFN通过PI3K/AKT/mTOR/4EBP1信号通路影响HT-29细胞的增殖Western blot实验结果显示,SFN处理HT-29细胞后,eIF4F翻译起始复合物相关蛋白的表达降低。作为其信号通路的上游,AKT和mTOR表达量也同时降低,P-AKT和P-mTOR的表达有所上升,而PI3K的表达量无明显差异(图3A)。裸鼠皮下移植瘤的免疫组织化学染色显示,与对照组相比,实验组中eIF4F翻译起始复合物相关蛋白的表达量降低,AKT的表达降低,P-AKT与P-mTOR的表达增加(图3B),这与Western blot检测结果相吻合。

图3 SFN降低相关通路蛋白及eIF4F复合物相关蛋白的表达 ×200A:SFN处理HT-29细胞后相关蛋白的Western blot结果;B:皮下移植瘤相关蛋白的免疫组化结果
3 讨论
本研究显示,eIF4F翻译起始复合物在人类CRC组织中上调,这是翻译起始的关键组成部分。同时,从十字花科蔬菜中提取的天然异硫氰酸酯SFN可通过抑制PI3K/AKT/mTOR/4EBP1信号通路,抑制原代CRC细胞中eIF4F复合物的上调。
mRNA翻译(蛋白质合成)是基因表达中最耗能的步骤,并且受到高度调控。翻译调控是细胞对环境的快速适应性反应,对维持细胞内蛋白质的稳态至关重要。因此,翻译过程在人体生长发育中起着重要的作用,mRNA翻译成蛋白质更是基因表达调控中的关键[5]。蛋白质翻译包括起始、延伸、终止和核糖体回收。起始是限速阶段,并受到eIF4F翻译起始复合物合成速度的限制[6]。eIF4F由支架蛋白eIF4G、mRNA5′Cap结合亚单位eIF4E和RNA解旋酶eIF4A三个部分组成。在组成蛋白质翻译起始复合物的所有起始因子中,由于eIF4E被认为是最不丰富的翻译起始因子,所以,eIF4E 与eIF4G 结合形成eIF4F复合物的过程,也就成为了翻译开始的限速步骤和控制蛋白质翻译、表达的关键点[7]。
eIF4E:eIF4G的形成受到小的翻译抑制因子4E结合蛋白(4EBPs)的严格调控,4EBPs与eIF4G竞争形成eIF4E:4EBP1复合物,并抑制eIF4E:eIF4G的形成[7]。eIF4E上4EBPs的结构结合位点与eIF4G重叠,从而使eIF4E能够同时与eIF4G或4EBP1独占结合。哺乳动物有3种4EBPs:4EBP1、4EBP2和4EBP3,其中4EBP1发挥着比其他因子更重要的生物学功能[8]。4EBP1与帽结合eIF4E的结合比与无帽eIF4E结合更快、更紧密。4EBP1和eIF4E之间的这种相互作用受到4EBP1磷酸化的严格调控,这反过来又会使4EBP1失活,并解离eIF4E:4EBP1相互作用。4EBP1的过度磷酸化在大多数人类癌症中被观察到,并且经常与肿瘤分级和不良预后相关。
文献[9]报道,PI3K/AKT/mTOR通路在细胞增殖、凋亡、代谢和血管生成等方面都发挥着重要作用。另外,PI3K/AKT/mTOR信号通路与许多人类疾病密切相关,包括缺血性脑损伤、神经退行性疾病和肿瘤[10]。PI3K/AKT/mTOR分为两部分:磷脂酰肌醇3-激酶(PI3K)及其下游分子丝氨酸/苏氨酸蛋白激酶B(PKB,又称AKT)。该通路被RTK和细胞因子受体激活。酪氨酸残基被磷酸化,为PI3K转位到膜上提供锚点,参与细胞外基质分子和细胞因子的转导。这一信号通路对细胞也有重要的生物学效应,如促进细胞活性和延缓衰老和死亡,与各类生理活动相联系。这一通路的功能障碍不仅与肿瘤的发生和发展有关,还与许多其他人类疾病有关,如白血病、糖尿病和精神分裂症。雷帕霉素(TOR)是真核细胞丝氨酸/苏氨酸激酶靶标,是细胞生长代谢的关键因子[11]。哺乳动物mTOR能感知细胞水平的代谢物(如氨基酸、葡萄糖、氧气和生长因子),调节从蛋白质合成到自噬的一系列过程。并在不同的复合体中起作用,即mTOR复合体1(mTORC1)和mTOR复合体2(mTORC2)[12]。
本研究显示,SFN可以通过抑制PI3K/AKT/mTOR/4EBP1信号通路来抑制eIF4F翻译起始复合物的形成,其原因可能是mTOR磷酸化引起的信号反馈导致AKT过度磷酸化和4EBP1去磷酸化。反过来,4EBP1被SFN去磷酸化并激活,其与eIF4E稳定结合,导致蛋白停止翻译。mTOR通过调节4EBP1的磷酸化在mRNA的翻译过程中起着关键作用。然而,即使AKT磷酸化被激活,PI3K的表达仍无显著变化。因此,可以发现mTOR磷酸化可诱导反馈,激活AKT磷酸化,影响其下游的4EBP1,从而影响蛋白质的翻译过程。
