不同时期高温胁迫对夏玉米物质生产性能及籽粒产量的影响
2022-10-31张川刘栋王洪章任昊赵斌张吉旺任佰朝刘存辉刘鹏
张川,刘栋,王洪章,任昊,赵斌,张吉旺,任佰朝,刘存辉,刘鹏
不同时期高温胁迫对夏玉米物质生产性能及籽粒产量的影响
张川1,刘栋1,王洪章1,任昊1,赵斌1,张吉旺1,任佰朝1,刘存辉2,刘鹏1
1作物生物学国家重点实验室/山东农业大学农学院,山东泰安 271018;2山东省种子管理总站,济南 250100
【目的】黄淮海夏玉米区高温胁迫频发、重发、持续期延长显著影响籽粒产量。本文以不同耐热型玉米品种为材料,探明大喇叭口期(V12)和开花期(VT)高温胁迫对两类品种叶片光合特性、碳同化物积累、分配和籽粒产量的影响。【方法】本研究以耐热型品种郑单958(ZD958)和热敏感型品种先玉335(XY335)为材料,以同时期适宜温度处理(昼32℃12 h/夜22℃12 h)为对照,使用自动控温控湿的高温棚模拟田间自然增温效果,设置V12期、VT期高温胁迫处理(昼38℃12 h/夜28℃12 h),比较高温胁迫后夏玉米叶面积指数(LAI)、碳代谢酶活性、光合速率、碳同化物积累和分配的动态变化特征,明确夏玉米物质生产性能及籽粒产量对高温胁迫的响应机制。【结果】高温胁迫后,两品种的LAI、碳代谢酶活性、净光合速率和干物质积累量均显著降低,ZD958和XY335的LAI、RuBP羧化酶活性、PEP羧化酶活性、净光合速率和干物质积累量比其对照分别降低了2.98%—4.21%、40.38%—54.46%、16.88%—30.60%、18.14%—25.49%、12.83%—19.38%和3.80%—5.07%、56.56%—76.16%、26.33%—33.66%、22.37%—34.62%、22.07%—26.72%,VT期高温胁迫的降幅大于V12期。高温胁迫后,夏玉米叶片蒸腾速率显著升高,但叶片水分利用效率显著下降。高温下两品种的13C同化量均显著降低,V12期高温胁迫后,ZD958和XY335的13C同化量分别降低了18.48%和22.82%,籽粒中13C同化量占比降低。高温胁迫显著降低穗粒数,千粒重虽有小幅提高,但籽粒产量显著降低。与适宜温度相比,V12期高温胁迫后ZD958穗粒数和产量分别降低了62.53%和45.87%;VT期高温胁迫后穗粒数和产量分别降低了70.53%和66.89%;V12期高温胁迫后XY335穗粒数和产量分别降低了70.50%和62.87%;VT期高温胁迫后分别降低了85.41%和80.61%;VT期高温胁迫降幅大于V12期高温胁迫,XY335的降幅大于ZD958。【结论】高温胁迫降低了夏玉米叶面积指数、叶片RuBP和PEP羧化酶活性,显著降低叶片光合速率和干物质生产性能。高温下穗粒数显著减少,抑制了碳同化物从叶片和茎秆向籽粒的转运,最终导致籽粒产量降低。VT期高温胁迫效应大于V12期,热敏感型品种XY335的降幅显著大于耐热型品种ZD958。
夏玉米;高温胁迫;碳代谢酶活性;碳同化物积累与分配;产量
0 引言
【研究意义】玉米是我国重要的粮食、饲料和经济作物,在保障国民经济安全中占有重要地位[1]。黄淮海夏玉米区播种面积占全国总面积的1/3以上,该地区光热资源丰富,但全球气候变化导致高温胁迫频发、重发,高温发生时间正处于黄淮海夏玉米生长发育与产量形成的关键时期,不利于玉米高产稳产[2-3]。高温胁迫后,夏玉米生育期缩短、光合速率降低、开花授粉过程受阻、空秆率和畸形穗率增加,穗粗、穗长、穗粒数和千粒重均显著降低[4],且对籽粒品质造成不利影响[1,5]。碳同化物在植物体内的积累与分配对作物的生长发育和产量形成至关重要,其包括无机碳在叶绿体中通过光合作用转化为有机碳的光合固定代谢过程、碳同化物的运输转化过程及碳同化物的积累与分配过程[6]。因此,研究高温胁迫后夏玉米碳同化物积累与分配的影响,探析其背后的生理生化机理,对于研发相关抵御高温栽培措施具有重要指导意义。【前人研究进展】高温胁迫是指温度升高到一定程度并持续一定时间,对作物生长发育及产量形成造成不可逆转的损伤[7]。作物在生长发育过程中遭遇高温胁迫后,各器官的基因表达、细胞结构、生理活动等均会发生不可逆的改变,造成器官功能部分甚至完全丧失,进而影响作物产量[8]。玉米虽为起源于热带地区的喜温作物,但在实际生产中高温胁迫仍严重威胁玉米籽粒产量和品质[9]。气温≥32℃即对玉米生长发育造成不良影响,气温≥38℃时玉米多数生理指标发生显著改变,生长发育受到严重影响[10]。光合作用是作物产量形成的基础,90%的干物质积累来源于光合作用,但光合作用对高温胁迫极其敏感[11]。高温胁迫后叶片生长受到抑制,叶面积指数下降,群体光能利用率降低[12-13]。RuBP羧化酶和PEP羧化酶是玉米叶片光合作用的两个关键酶,其活性高低直接影响暗反应中CO2的同化效率[14]。高温胁迫后RuBP羧化酶和PEP羧化酶的活性显著降低[15]。环境温度超过30℃时,RuBP羧化酶活性即开始逐渐丧失[16],当温度≥40℃时RuBP羧化酶(Rubisco)和Rubisco活化酶活性受抑制是光合速率降低的重要原因[17-18]。逆境胁迫显著影响玉米碳同化物的积累过程,改变其在营养器官中的分配,减少穗中碳同化物的分配比例[19-21]。研究发现,随干旱胁迫程度的增加,茎、叶等营养器官中干物质分配占比提高,而穗部干物质占比逐渐减小[19]。高温胁迫显著降低玉米植株干物质积累量、结实率和穗粒数,最终导致籽粒产量降低[20-21]。【本研究切入点】过去关于夏玉米高温胁迫的报道多集中于生长发育、光合特性、雌雄穗分化以及产量形成等方面,且以灌浆期高温胁迫研究较多,但对夏玉米大喇叭口期和开花期高温胁迫如何影响其碳同化物积累与分配以及造成产量降低的生理机制的研究报道较少。【拟解决的关键问题】本文通过设置大田试验,采用自动控温控湿高温棚模拟田间高温胁迫,研究大喇叭口期和开花期高温胁迫对夏玉米光合性能、碳同化物积累与分配及最终籽粒产量的影响,为丰富夏玉米高温胁迫机理和研发相关抵御高温栽培措施提供理论依据。
1 材料与方法
1.1 试验设计与材料
试验于2019—2020年在山东农业大学黄淮海区域玉米技术创新中心(36°10′N,117°09′E)进行,该地属于半湿润暖温带大陆性季风气候,试验田为棕壤土,耕层0—20 cm含有机质11.2 g·kg-1、全氮1.1 g·kg-1、碱解氮60.4 mg·kg-1、速效磷41.0 mg·kg-1、速效钾75.6 mg·kg-1。两年均于6月5日播种,10月5日收获。肥料用量按照高产田的产量水平确定,其中纯N 270 kg·hm-2、P2O5105 kg·hm-2、K2O 210 kg·hm-2。40%的氮肥(包膜缓控尿素,含N 42%)和全部磷肥(过磷酸钙,含P2O512%)、全部钾肥(硫酸钾,含K2O 51%)作为基肥施用,剩余60%的氮肥于大喇叭口期高温处理前开沟追施。生长期间根据土壤墒情及降水情况采用微喷带统一灌溉,分别于播种前、拔节期和小喇叭口期灌溉80 mm、50 mm和50 mm,之后自然降水满足夏玉米生长所需,未再补充灌溉,高温处理期间无降水。按统一的植保方案进行病虫害防治。
试验选用耐热型品种郑单958(ZD958)和热敏感型品种先玉335(XY335)为材料,种植密度为67 500株/hm2。采用裂-裂区设计,主区为高温处理时期,分别为大喇叭口期(V12期)和开花期(VT期);裂区为不同温度处理,分别为高温胁迫处理和常温对照,依据黄淮海夏玉米区近10年气象资料分析,将高温胁迫处理设置为昼(7:00—19:00)夜(19:00—次日7:00)平均温度为38℃和28℃,常温对照处理设置为昼(7:00—19:00)夜(19:00—次日7:00)平均温度为32℃和22℃;裂-裂区为不同耐热型玉米品种。试验共8个处理,每个处理重复3次,小区面积18 m2。试验选用高透光率树脂薄膜和钢架结构制作控温控湿设施,薄膜厚度0.1 mm,透光率92%以上。棚顶部与底部均留出空隙进行通风,棚内放置自动加热装置(大功率自动加温机)和控湿装置(自动加/除湿机),按照设定温度进行处理,处理结束后拆除薄膜,使其自然生长。高温处理期间,每天上午11:00使用AccuPAR LP-80型植物冠层分析仪(DECAGON,USA)测定棚内光照强度,用EGM-4型(PP-Systems,UK)红外线CO2分析仪测定棚内CO2浓度,连续测定7 d。利用GSP-6型(Elitheh,CN)温湿度计每隔15 min自动记录环境温湿度。高温胁迫对不同处理田间小气候的影响如表1所示,不同处理日平均温度变化如图1所示。
1.2 测定项目及方法
1.2.1 叶面积指数 于高温处理7 d后、开花期(VT)、灌浆期(R2)、乳熟期(R3)、蜡熟期(R5)、完熟期(R6),各处理选取3株长势一致的植株,测量其全部展开叶和未展开叶的叶长、叶宽。完全展开叶叶面积计算公式为:LA=L×W×0.75,未完全展开叶叶面积计算公式为:LA=L×W×0.5。式中,LA为叶面积,L为叶片长度,W为叶片宽度,0.75和0.5为校正系数。单株叶面积为全株所有叶片叶面积之和。叶面积指数的计算公式为:LAI=LA/GA,式中,LA为该土地面积上的总叶面积,GA为土地面积。
1.2.2 净光合速率和蒸腾速率 每个处理选取长势均匀的植株15株,于高温处理7 d后、VT期、R2期、R3期、R5期使用CIRAS-II型便携式光合仪,选择晴天上午10:00至12:00在1 600 µmol·m-2·s-1光照强度、390 µmol·mol-1CO2浓度和80% H2O浓度条件下测定净光合速率(n)和蒸腾速率(E)(V12期测定第12片完全展开叶,VT期及VT期以后测定穗位叶)。叶片水分利用效率为净光合速率与蒸腾速率的比值。

表1 高温胁迫对夏玉米田间小气候的影响
同一年度同列标以不同小写字母的值差异显著(<0.05)。CK:常温对照;HT:高温处理。下同
Values followed by different small letters within a column are significantly different among different treatment in the same year at 0.05 probability level. CK: normal temperature; HT: high temperature. the same as below

图1 高温处理期间不同处理日平均温度变化
1.2.3 碳代谢酶活性测定 在高温处理7 d后、VT期、R2期、R3期、R5期取植株新鲜叶片(V12期取植株第12片完全展开叶,VT期及VT期以后取植株穗位叶),液氮速冻后,-80℃冷冻保存。
酶的提取方法参照张翼飞[22]的方法稍加修改后进行,称取鲜叶片0.500 g左右,置于预冷的研钵中,加适量经酸洗的预冷过的石英砂,加6 mL预冷过的提取缓冲液(提取缓冲液包括0.1 mol·L-1的Tris-HCl缓冲液(pH 8.4)、10 mmol·L-1的MgCl2溶液、1 mmol·L-1的EDTA溶液、7 mmol·L-1的β-巯基乙醇溶液、5%(v/v)甘油、1% PVP),冰浴研磨至匀浆后转移至10 mL离心管中,在4℃下15 000×离心10 min,上清液即酶液。酶活性参照魏爱丽等[23]的分光光度法进行测定。
1.2.4 干物质积累量 于高温处理7 d后、VT期、R2期、R3期、R5期、R6期,在田间选取3株生长一致的植株地上部,按茎秆、叶片、雄穗、苞叶、籽粒和穗轴分开,105℃杀青30 min后80℃烘干至恒重,称量各部分干重,计算单株干物质积累量。
1.2.513C同位素的标记与测定 采用13CO2标记法,高温处理的第1天上午10:00开始进行标记,V12期高温处理饲喂第12片完全展开叶,VT期高温处理饲喂穗位叶,同化塑料袋容积根据叶片大小进行调整,使用注射器将50 mL13CO2注入袋中,标记1 h,结束后将同化塑料袋撤下,每个处理饲喂5株。在R6期取样,将植株按茎秆、叶片、雄穗、苞叶、籽粒和穗轴分开,105℃杀青30 min,80℃烘至恒重并称重。将13C同位素标记烘干样品磨粉过200目筛,利用锡囊准确称取4 mg左右样品,用Elementar vario MICRO cube元素分析仪(Elementar,德国)测定各器官全碳含量,用Isoprime 100稳定同位素质谱仪(Isoprime,英国)测定各器官13C丰度值。相关计算公式如下:
13C丰度Fi(%)=((δ13C+1000)×RPBD)/((δ13C+1000)×RPBD+1000)×100。
式中,RPBD(碳同位素的标准比值)=0.0112372。
各器官含碳量Ci(mg)=器官总质量(g)×全碳含量(%)/100×1000;
进入各器官的13C量13Ci(mg)=Ci×Fi/100×1000。式中,Ci为各器官含碳量(mg),Fi为各器官13C丰度。整株13C固定量为进入各器官的13C量之和。
13C在各器官的分配率13Ci(%)=13Ci/13C净吸收×100。
1.2.6 籽粒产量的测定 于R6期每个处理选取3个重复,各取30个果穗考种,测定其穗粒数及千粒重,并计算籽粒产量。
1.3 统计与分析
试验数据采用Excel 2016处理,采用DPS 11.0软件进行处理与统计分析并采用LSD法进行差异比较,用Sigmaplot 10.0(Systat Software,San Jose,CA)作图。
2 结果
2.1 高温胁迫对夏玉米叶面积指数的影响
如图2所示,高温胁迫后,两个品种的LAI均低于其相应的对照处理。V12期高温胁迫后,ZD958和XY335的LAI比各自对照分别降低了4.21%、5.07%(2019年)和2.26%、8.66%(2020年);VT期高温胁迫后,ZD958和XY335的LAI分别降低了2.98%、3.80%(2019年)和3.22%、2.89%(2020年)。随着生育进程推进,LAI降幅呈现增加的趋势,下降速率也显著高于对照。不同品种之间比较,XY335的降幅要显著高于ZD958。
2.2 高温胁迫对夏玉米碳代谢酶活性的影响
高温胁迫显著降低了两品种叶片的RuBP和PEP羧化酶活性,VT期高温胁迫的效应大于V12期,对XY335的胁迫效应大于ZD958。高温胁迫后,各处理酶活性均有所回升,但仍低于对照,且随着生育进程推进,各处理酶活性均呈现下降的趋势(图3—4)。2019年V12期高温胁迫后,ZD958的RuBP和PEP羧化酶活性分别降低了40.38%和16.88%;VT期高温胁迫后降幅分别为54.46%和30.60%。V12期高温胁迫后,XY335的RuBP和PEP羧化酶活性分别降低了56.56%和26.33%,VT期高温胁迫后分别降低了76.16%和33.66%。高温胁迫对RuBP羧化酶活性的影响显著高于PEP羧化酶,两年表现趋势一致。

ZD-CK:郑单958常温对照处理;ZD-HT:郑单958高温胁迫处理;XY-CK:先玉335常温对照处理;XY-HT:先玉335高温胁迫处理。V12+7:大喇叭口期高温处理7 d;VT:开花期;VT+7:开花期高温处理7 d;R2:灌浆期;R3:乳熟期;R5:蜡熟期;R6:完熟期。下同
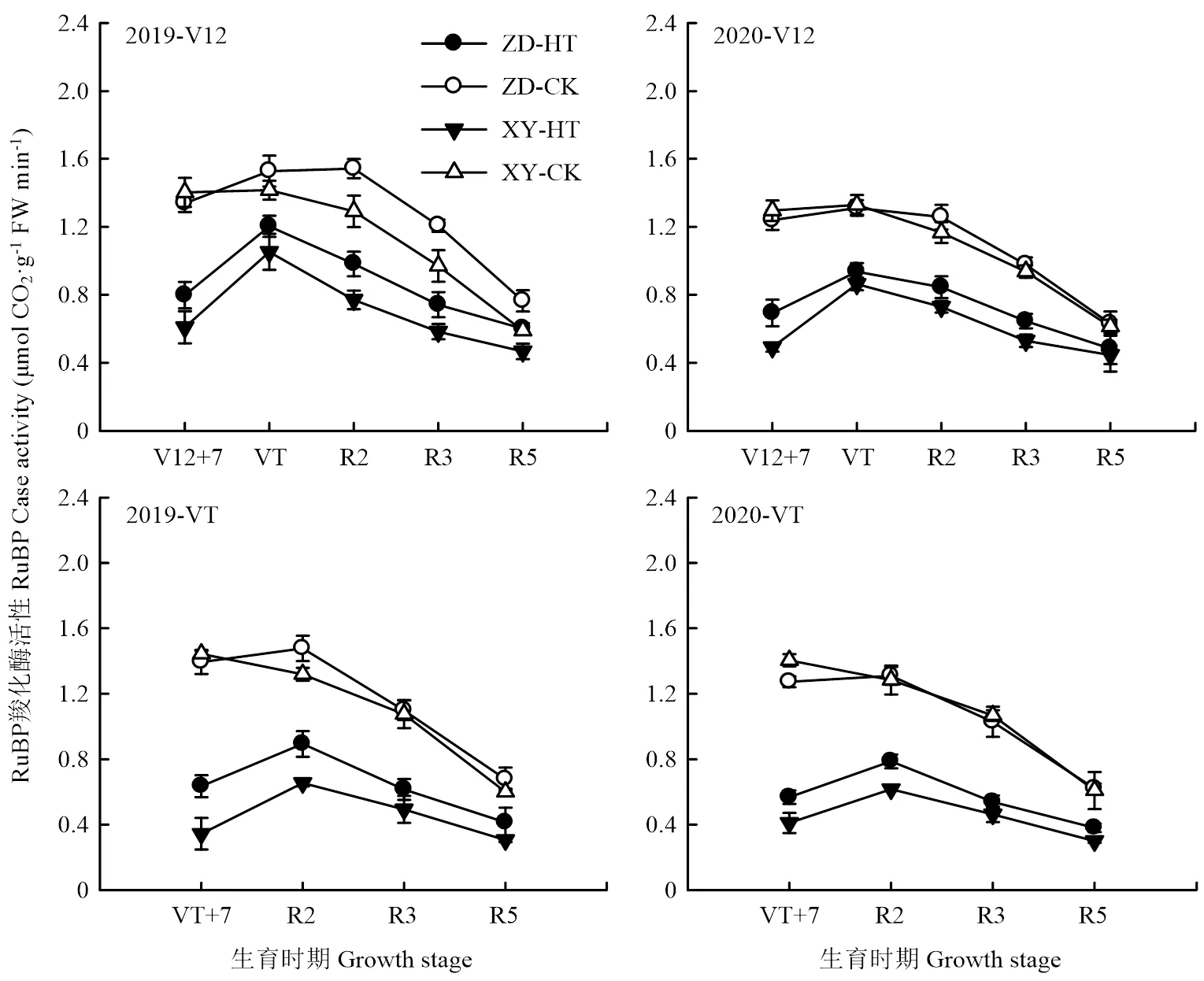
图3 高温胁迫对夏玉米RuBP羧化酶活性的影响
2.3 高温胁迫对夏玉米叶片净光合速率的影响
高温胁迫后,两个品种的穗位叶净光合速率均显著降低,高温处理结束后,对夏玉米光合作用的影响减弱,但仍低于对照(图5)。V12期高温胁迫后,ZD958和XY335净光合速率分别降低了18.14%、22.37%(2019年)和18.11%、22.40%(2020年);VT期高温胁迫后,ZD958和XY335分别降低了25.49%、34.62%(2019年)和25.55%、34.66%(2020年)。两品种VT期比V12期受高温的影响更大,其中XY335比ZD958受高温胁迫的影响更显著。
2.4 高温胁迫对夏玉米叶片蒸腾速率和叶片水分利用效率的影响
高温胁迫后,夏玉米叶片蒸腾速率显著升高,但叶片水分利用效率显著下降(图6—7)。V12期高温胁迫后,ZD958和XY335蒸腾速率分别升高64.24%和81.78%,叶片水分利用效率分别降低4.03%和3.99%;VT期高温胁迫后,ZD958和XY335蒸腾速率分别升高102.19%和58.66%,叶片水分利用效率分别降低5.22%和4.19%。两年表现趋势一致。
2.5 高温胁迫对夏玉米完熟期13C同化物积累与分配的影响
高温胁迫后ZD958和XY335的植株13C同化量均显著降低,两年表现趋势一致(表2)。2019年V12期高温胁迫后,ZD958和XY335的13C同化量分别降低了18.48%和22.82%,其中,ZD958茎秆和叶片13C同化量占比相较于CK分别提高了13.57%和13.08%,籽粒13C同化量占比降低了26.77%;XY335茎秆和叶片13C同化量占比分别提高了18.70%和18.74%,籽粒13C同化量占比降低了37.50%,XY335的13C同化量降低幅度和各器官中13C同化量占比的变化幅度均高于ZD958。VT期高温胁迫后两个敏感型不同的品种表现趋势与V12期基本一致,但其变化幅度要高于V12期高温胁迫,其中,ZD958的13C同化量及籽粒13C同化量占比分别下降了20.61%和32.42%; XY335的13C同化量及籽粒13C同化量占比分别下降了33.40%和48.09%。

图4 高温胁迫对夏玉米PEP羧化酶活性的影响

图5 高温胁迫对夏玉米叶片净光合速率的影响

图6 高温胁迫对夏玉米叶片蒸腾速率的影响

图7 高温胁迫对夏玉米叶片瞬时水分利用效率的影响
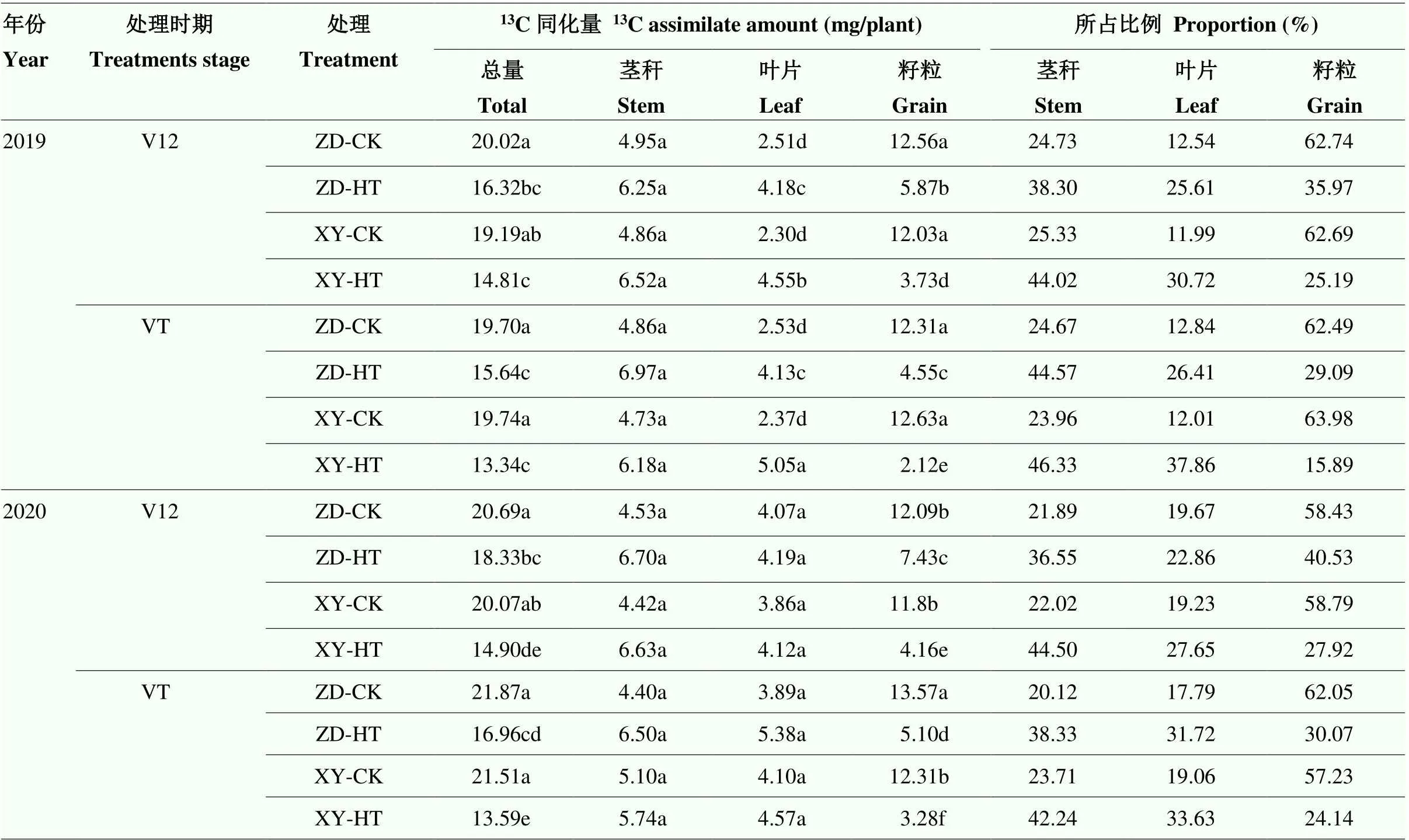
表2 高温胁迫对夏玉米完熟期13C同化物积累与分配的影响
ZD-CK:郑单958常温对照处理;ZD-HT:郑单958高温胁迫处理;XY-CK:先玉335常温对照处理;XY-HT:先玉335高温胁迫处理。同年同列数据后不同小写字母表示处理间差异达0.05显著水平。下同
ZD-CK: ZD958 normal temperature treatment; ZD-HT: ZD958 high temperature stress treatment; XY-CK: XY335 normal temperature treatment; XY-HT: XY335 high temperature stress treatment. Values followed by different letters within the same column in the same year are significantly different between treatments at<0.05. The same as below
2.6 高温胁迫对夏玉米干物质积累及籽粒产量的影响
由表3可知,高温胁迫后夏玉米干物质积累量显著降低,且随生育进程的推进,其降幅呈增加趋势。2019年V12期高温胁迫后,ZD958和XY335干物质积累量分别降低19.38%和26.72%,至R6期降幅进一步增大,分别为29.48%和31.43%;VT期高温胁迫后,ZD958和XY335分别降低12.83%和22.07%,至R6期降幅增至32.88%和37.54%,两年趋势表现一致。高温胁迫后夏玉米产量显著降低,V12期高温胁迫后,ZD958和XY335分别降低了45.87%、62.53%(2019年)和42.53%、60.05%(2020年);VT期高温胁迫后,ZD958和XY335分别降低了66.89%、80.61%(2019年)和53.99%、66.51%(2020年)。相比于V12期高温处理,VT期高温处理产量损失高出6.46%—21.02%。XY335产量降低幅度比ZD958高出12.52%—17.52%。高温胁迫显著减少了穗粒数,V12期高温胁迫后,ZD958和XY335分别减少了62.87%、70.50%(2019年)和56.73%、65.52%(2020年);VT期高温胁迫后,ZD958和XY335分别降低了70.53%、85.41%(2019年)和64.21%、72.40%(2020年),其表现趋势和产量一致。高温胁迫后,夏玉米粒重显著增加,具体表现为每1000粒粒重增加2.9—12.07 g,且XY335粒重大于ZD958。高温下穗粒数的降低幅度远大于粒重的增加幅度。此外,高温显著降低了夏玉米收获指数,高温胁迫后夏玉米收获指数仅为0.14—0.37,而常温对照收获指数为0.48—0.58。
3 讨论
3.1 高温胁迫对夏玉米光合特性的影响
叶片是植物主要的光合作用器官,绿叶面积大小和高值持续期长短直接影响光合性能,进而影响作物产量[24]。前人研究发现,高温胁迫后玉米幼苗瘦弱、叶片生长速率变缓并很快变黄出现早衰[25]。高温处理会降低叶面积,进而影响植株干物质量[26-27]。本研究结果发现,高温胁迫后ZD958和XY335的LAI均显著降低,且XY335的降幅较大;且随着生育进程推进,高温胁迫处理的LAI下降速率也显著高于对照。由此可见,高温处理后夏玉米较小的LAI及其快速下降可能是夏玉米干物质生产性能降低的重要原因(图2)。此外,高温还会直接抑制植物的光合作用,进而影响碳水化合物的积累与转运[28]。高温下ZD958和XY335穗位叶的净光合速率均显著降低,其中XY335比ZD958反应更加敏感,开花期比大喇叭口期的高温胁迫效应增加(图5)。高温结束后各处理的净光合速率均有一定幅度的上升,但仍然低于对照,由此可知,高温胁迫造成的光合作用损伤难以恢复,高温下光合速率降低是干物质减少的直接原因。高温胁迫后夏玉米叶片蒸腾速率显著升高是植株对高温逆境做出的主动调节过程,植物通过提高蒸腾速率散发更多的水分来降低叶片温度,减少叶片损伤,进而降低叶片水分利用效率(图6—7)。
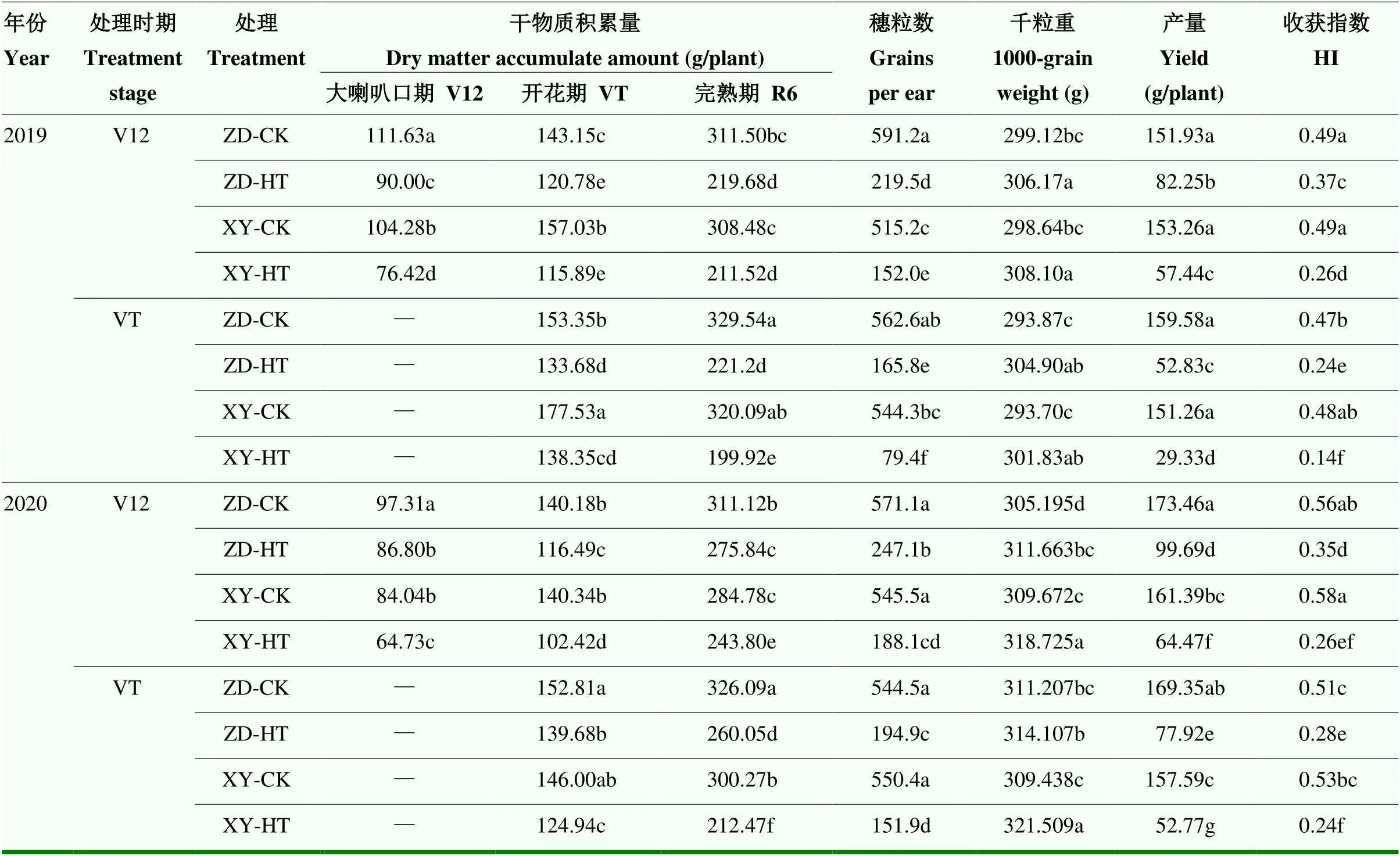
表3 高温胁迫对夏玉米干物质积累及籽粒产量的影响
RuBP羧化酶和PEP羧化酶是植物碳代谢过程中两个重要的关键酶。RuBP羧化酶催化1,5-二磷酸核酮糖(RuBP)合成三磷酸甘油酸,其活性高低直接影响暗反应中CO2的同化效率,与PSⅡ反应中心的光化学效率密切相关[14,29]。PEP羧化酶催化磷酸烯醇式丙酮酸(PEP)合成草酰乙酸,在C4植物叶片中起着固定原初CO2的作用[30]。植物体内RuBP羧化酶和PEP羧化酶活性均受到品种特性与环境因子的影响,并且对逆境胁迫有一定的适应性[31]。本研究结果表明,高温胁迫显著降低了两个品种叶片的RuBP羧化酶和PEP羧化酶活性。不同处理间比较,VT期高温处理碳代谢酶活性的降幅大于V12期,XY335叶片酶活性的降幅大于ZD958。高温胁迫下耐热型品种ZD958更高的碳代谢酶活性可能是其保持较高光合性能、积累更多干物质的主要原因(图3—4)。
3.2 高温胁迫对夏玉米碳同化物积累与分配及产量的影响
作物高产不仅要求功能叶片有较强的光合生产能力,而且要求光合器官中形成的光合产物能够合理分配、高效运输[32]。高温胁迫显著影响夏玉米植株的正常生长发育和形态建成,不同器官对高温响应不同,抽雄期高温胁迫处理的叶片干物质量比对照有所增加,而根和茎的干物质量却比对照有所下降[33]。穗期阶段是玉米营养器官生长与生殖器官分化发育同时并进阶段,此时遭遇逆境胁迫会导致干物质向茎和叶片中的分配比例增加,向穗中的转运积累减少[19]。前人研究指出,高温胁迫会引起玉米叶片光合物质生产能力下降、营养器官早衰,同时伴随着呼吸消耗增加,导致净同化物减少、同化物供应不足,穗粒数和粒重降低,进而导致玉米的产量降低[34]。本研究发现,高温胁迫显著降低了夏玉米干物质积累量,表明其生长发育受到抑制,植株生长缓慢,至完熟期与对照间的差异进一步增大。结果同时表明,VT期高温胁迫效应大于V12期,对XY335干物质积累量的影响大于ZD958(图6)。
碳素是决定植物生长发育的基本元素,其在植物体内的转运和分配决定着植物的生长过程及其生产力,也关系到植物对环境胁迫的适应能力[35]。13C同位素示踪分析表明,高温不仅降低了ZD958和XY335的13C积累量,而且也改变了13C同化物在各器官中的分配比例。高温胁迫后,茎秆和叶片中13C同化物占比提高,籽粒中13C同化物占比降低,说明高温抑制了碳同化物从茎秆和叶片向籽粒的运输,阻碍了13C同化物向籽粒的分配(表1)。由此可见,高温胁迫后籽粒产量的显著降低一方面是因为高温降低了夏玉米干物质积累量,另一方面是因为高温抑制了营养器官中碳同化物向籽粒的转运。此外,高温胁迫后穗粒数显著降低,粒重显著增加,但远不足以弥补库容减少所带来的产量损失,说明高温胁迫处理下穗粒数显著减少是产量降低的主要原因(表2)。前人研究结果表明,高温胁迫后夏玉米穗粒数减少的主要原因是高温抑制了雌雄穗的生长发育,导致散粉量减少、散粉持续期缩短、雌雄间隔增大,无法完成授粉受精过程[20-21]。穗粒数的降低即籽粒库容的减少降低了对碳同化物的供应需求,这也可能是高温抑制了叶片和茎秆中碳同化物向籽粒转运的重要原因。耐热型品种ZD958的穗粒数、碳同化物积累量及其向籽粒的分配比例高于热敏感型品种XY335,其受高温胁迫的减产效应也显著低于XY335。
4 结论
高温胁迫后,夏玉米LAI、叶片RuBP和PEP羧化酶活性均显著降低,致使叶片光合速率显著降低,严重降低干物质生产性能。高温下穗粒数显著减少,抑制了碳同化物从叶片和茎秆向籽粒的转运,最终导致籽粒产量降低。VT期高温胁迫对夏玉米干物质生产性能和籽粒产量的影响显著大于V12期;两个时期热敏感型品种XY335的降幅显著大于耐热型品种ZD958。
[1] 李少昆, 赵久然, 董树亭, 赵明, 李潮海, 崔彦宏, 刘永红, 高聚林, 薛吉全, 王立春, 王璞, 陆卫平, 王俊河, 杨祁峰, 王子明. 中国玉米栽培研究进展与展望. 中国农业科学, 2017, 50(11): 1941-1959.
LI S K, ZHAO J R, DONG S T, ZHAO M, LI C H, CUI Y H, LIU Y H, GAO J L, XUE J Q, WANG L C, WANG P, LU W P, WANG J H, YANG Q F, WANG Z M. Advances and prospects of maize cultivation in China. Scientia Agricultura Sinica, 2017, 50(11): 1941-1959. (in Chinese)
[2] 任寒, 刘鹏, 董树亭, 张吉旺, 赵斌. 高温胁迫影响玉米生长发育的生理机制研究进展. 玉米科学, 2019, 27(5): 109-115.
REN H, LIU P, DONG S T, ZHANG J W, ZHAO B. Research advancements of effect of high temperature stress on growth and development of maize. Journal of Maize Sciences, 2019, 27(5): 109-115. (in Chinese)
[3] 王群, 赵向阳, 刘东尧, 闫振华, 李鸿萍, 董朋飞, 李潮海. 淹水弱光复合胁迫对夏玉米根形态结构、生理特性和产量的影响. 中国农业科学, 2020, 53(17): 3479-3495.
WANG Q, ZHAO X Y, LIU D Y, YAN Z H, LI H P, DONG P F, LI C H. Root morphological, physiological traits and yield of maize under waterlogging and low light stress. Scientia Agricultura Sinica, 2020, 53(17): 3479-3495. (in Chinese)
[4] ZHANG Q, YANG Z Q. Impact of extreme heat on corn yield in main summer corn cultivating area of China at present and under future climate change. International Journal of Plant Production, 2019, 13(4): 267-274.
[5] JHA U C, BOHRA A, SINGH N P. Heat stress in crop plants: Its nature, impacts and integrated breeding strategies to improve heat tolerance. Plant Breeding, 2014, 133(6): 679-701.
[6] 苌建峰. 不同基因型玉米碳氮代谢差异研究[D]. 郑州: 河南农业大学, 2007.
CHANG J F. Study on differences of carbon and nitrogen metabolism in different genotype maize[D]. Zhengzhou: Henan Agricultural University, 2007. (in Chinese)
[7] LEMMA A T. Action and reaction of plants to high temperature: improving response of wheat to heat stress. Current Journal of Applied Science and Technology, 2021, 40(10): 62-70.
[8] ZHEN F X, ZHOU J J, MAHMOOD A, WANG W, CHANG X N, LIU B, LIU L L, CAO W X, ZHU Y, TANG L. Quantifying the effects of short term heat stress at booting stage on nonstructural carbohydrates remobilization in rice. The Crop Journal, 2020, 8(2): 194-212.
[9] 张保仁, 董树亭, 胡昌浩, 王空军. 高温对玉米籽粒淀粉合成及产量的影响. 作物学报, 2007, 33(1): 38-42.
ZHANG B R, DONG S T, HU C H, WANG K J. Effect of high air temperature during different growth stage on starch synthesis in grain and yield in maize (L.). Acta Agronomica Sinica, 2007, 33(1): 38-42. (in Chinese)
[10] 王海梅. 高温胁迫对河套灌区玉米生理指标及产量构成要素的影响. 干旱气象, 2015, 33(1): 59-62.
WANG H M. Temperature stress on physiological indexes and yield components of maize in Hetao irrigation district. Journal of Arid Meteorology, 2015, 33(1): 59-62. (in Chinese)
[11] XU Y F, CHU C C, YAO S G. The impact of high temperature stress on rice: Challenges and solutions. The Crop Journal, 2021, 9(5): 963-976.
[12] 赵龙飞, 李潮海, 刘天学, 王秀萍, 僧珊珊. 花期前后高温对不同基因型玉米光合特性及产量和品质的影响. 中国农业科学, 2012, 45(23): 4947-4958.
ZHAO L F, LI C H, LIU T X, WANG X P, SENG S S. Effect of high temperature during flowering on photosynthetic characteristics and grain yield and quality of different genotypes of maize (L.). Scientia Agricultura Sinica, 2012, 45(23): 4947-4958. (in Chinese)
[13] 孙宁宁. 玉米叶、粒对高温胁迫的响应[D]. 郑州: 河南农业大学, 2017.
SUN N N. Responses of maize (L.) leaf and kernel to heat stress[D]. Zhengzhou: Henan Agricultural University, 2017. (in Chinese)
[14] 陈传晓. 不同积温带春玉米碳代谢机理及化学调控效应的研究[D]. 保定: 河北农业大学, 2013.
CHEN C X. Studies on the mechanism of carbon metabolism of spring maize and chemical regulation effects under different accumulated temperature zones[D]. Baoding: Hebei Agricultural University, 2013. (in Chinese)
[15] 孙胜楠, 王强, 孙晨晨, 刘丰娇, 毕焕改, 艾希珍. 黄瓜幼苗光合作用对高温胁迫的响应与适应. 应用生态学报, 2017, 28(5): 1603-1610.
SUN S N, WANG Q, SUN C C, LIU F J, BI H G, AI X Z. Response and adaptation of photosynthesis of cucumber seedlings to high temperature stress. Chinese Journal of Applied Ecology, 2017, 28(5): 1603-1610. (in Chinese)
[16] 张英华, 杨佑明, 曹莲, 郝杨凡, 黄菁, 李金鹏, 姚得秀, 王志敏. 灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响. 作物学报, 2015, 41(1): 136-144.
ZHANG Y H, YANG Y M, CAO L, HAO Y F, HUANG J, LI J P, YAO D X, WANG Z M. Effect of high temperature on photosynthetic capability and antioxidant enzyme activity of flag leaf and non-leaf organs in wheat. Acta Agronomica Sinica, 2015, 41(1): 136-144. (in Chinese)
[17] MAESTRI E, KLUEVA N, PERROTTA C, GULLI M, NGUYEN H T, MARMIROLI N. Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Molecular Biology, 2002, 48(5): 667-681.
[18] MORALES D, RODRIGUEZ P, DELL'AMICO J, NICOLAS E, TORRECILLAS A, SANCHEZ-BLANCO M J. High temperature preconditioning and thermal shock imposition affects water relations, gas exchange and root hydraulic conductivity in tomato. Biologia Plantarum, 2003, 47(2): 203-208.
[19] 贾双杰, 李红伟, 江艳平, 赵国强, 王和洲, 杨慎骄, 杨青华, 郭家萌, 邵瑞鑫. 干旱胁迫对玉米叶片光合特性和穗发育特征的影响. 生态学报, 2020, 40(3): 854-863.
JIA S J, LI H W, JIANG Y P, ZHAO G Q, WANG H Z, YANG S J, YANG Q H, GUA J M, SHAO R X. Effects of drought on photosynthesis and ear development characteristics of maize. Acta Ecologica Sinica, 2020, 40(3): 854-863. (in Chinese)
[20] 穆心愿, 马智艳, 张兰薰, 付景, 刘天学, 丁勇, 夏来坤, 张凤启, 张君, 齐建双, 赵霞, 唐保军. 不同耐/感玉米品种的叶片光合荧光特性、授粉结实和产量构成因素对花期高温的反应. 中国生态农业学报(中英文), 2022, 30(1): 57-71.
MU X Y, MA Z Y, ZHANG L X, FU J, LIU T X, DING Y, XIA L K, ZHANG F Q, ZHANG J, QI J S, ZHAO X, TANG B J. Responses of the photosynthetic fluorescence characteristics, pollination and yield compositions of different tolerant/susceptible maize varieties to high temperature during flowering. Chinese Journal of Eco-Agriculture, 2022, 30(1): 57-71. (in Chinese)
[21] 闫振华, 刘东尧, 贾绪存, 杨琴, 陈艺博, 董朋飞, 王群. 花期高温干旱对玉米雄穗发育、生理特性和产量影响. 中国农业科学, 2021, 54(17): 3592-3608.
YAN Z H, LIU D Y, JIA X C, YANG Q, CHEN Y B, DONG P F, WANG Q. Maize tassel development, physiological traits and yield under heat and drought stress during flowering stage. Scientia Agricultura Sinica, 2021, 54(17): 3592-3608. (in Chinese)
[22] 张翼飞. 施氮对甜菜氮素同化与碳代谢的调控机制研究[D]. 哈尔滨: 东北农业大学, 2013.
ZHANG Y F. Study on regulation mechanism of nitrogen application on nitrogen assimilation and carbon metabolism in sugar beet (L.)[D]. Harbin: Dongbei Agricultural University, 2013. (in Chinese)
[23] 魏爱丽, 张英华, 黄琴, 王志敏. 小麦不同绿色器官光合速率与碳同化酶活性及其基因型差异研究. 作物学报, 2007, 33(9): 1426-1431.
WEI A L, ZHANG Y H, HUANG Q, WANG Z M. Dynamic characteristics of photosynthetic rate and carbon assimilation enzyme activities of different green organs in different genotypes of wheat. Acta Agronomica Sinica, 2007, 33(9): 1426-1431. (in Chinese)
[24]黄振喜, 王永军, 王空军, 李登海, 赵明, 柳京国, 董树亭, 王洪军, 王军海, 杨今胜. 产量15000 kg·hm-2以上夏玉米灌浆期间的光合特性. 中国农业科学, 2007, 40(9): 1898-1906.
HUANG Z X, WANG Y J, WANG K J, LI D H, ZHAO M, LIU J G, DONG S T, WANG H J, WANG J H, YANG J S. Photosynthetic characteristics during grain filling stage of summer maize hybrids with high yield potential of 15000 kg·ha-1. Scientia Agricultura Sinica, 2007, 40(9): 1898-1906. (in Chinese)
[25] 方芳, 何序晨, 张志豪, 张勤, 关亚静, 胡晋, 胡伟民. 玉米自交系苗期对高温胁迫的响应机制及其抗逆性. 浙江农业学报, 2019, 31(7): 1045-1056.
FANG F, HE X C, ZHANG Z H, ZHANG Q, GUAN Y J, HU J, HU W M. Response mechanism and stress resistance of maize inbred lines to high temperature stress at seedling stage. Acta Agriculturae Zhejiangensis, 2019, 31(7): 1045-1056. (in Chinese)
[26] 曲明南. CO2升高和短期高温胁迫对玉米幼苗生理生化指标的影响[D]. 沈阳: 沈阳农业大学, 2013.
QU M N. Effects of elevated CO2and short term heat stress on physiological and biochemical variables in maize seedlings[D]. Shenyang: Shenyang Agricultural University, 2013. (in Chinese)
[27] 付景, 孙宁宁, 刘天学, 杨豫龙, 赵霞, 李潮海. 高温胁迫对玉米形态、叶片结构及其产量的影响. 玉米科学, 2019, 27(1): 46-53.
FU J, SUN N N, LIU T X, YANG Y L, ZHAO X, LI C H. Effect of high temperature stress on morphology, leaf structure and grain yield of maize. Journal of Maize Sciences, 2019, 27(1): 46-53. (in Chinese)
[28] 陈岩, 岳丽杰, 杨勤, 张会玲, 柯国华, 刘永红. 高温热害对玉米生长发育的影响及研究进展. 耕作与栽培, 2019, 39(1): 26-31.
CHEN Y, YUE L J, YANG Q, ZHANG H L, KE G H, LIU Y H. Effects of high temperature on maize development and its research advances. Tillage and Cultivation, 2019, 39(1): 26-31. (in Chinese)
[29] FUKAYAMA H, UEGUCHI C, NISHIKAWA K, KATOH N, ISHIKAWA C, MASUMOTO C, HATANAKA T, MISOO S. Overexpression of rubisco activase decreases the photosynthetic CO2assimilation rate by reducing rubisco content in rice leaves. Plant & cell physiology, 2012, 53(6): 976-986.
[30] LIAN L, LIN Y L, WEI Y D, HE W, CAI Q H, HUANG W, ZHENG Y M, XU H B, WANG F X, ZHU Y S, LUO X, XIE H A, ZHANG J F. PEPC of sugarcane regulated glutathione S-transferase and altered carbon–nitrogen metabolism under different N source concentrations in. BMC Plant Biology, 2021, 21(1): 287.
[31] 毕焕改, 董绪兵, 王美玲, 艾希珍. 钙和水杨酸对亚适温弱光下黄瓜幼苗光合酶活性和基因表达的影响. 园艺学报, 2015, 42(1): 56-64.
BI H G, DONG X B, WANG M L, AI X Z. Foliar spray calcium and salicylic acid improve the activities and gene expression of photosynthetic enzymes in cucumber seedlings under low light intensity and suboptimal temperature. Acta Horticulturae Sinica, 2015, 42(1): 56-64. (in Chinese)
[32] 罗宏海, 李俊华, 张宏芝, 何在菊, 勾玲, 张旺锋. 源库调节对新疆高产棉花产量形成期光合产物生产与分配的影响. 棉花学报, 2009, 21(5): 371-377.
LUO H H, LI J H, ZHANG H Z, HE Z J, GOU L, ZHANG W F. Effects of source and sink manipulation on transportation and allocation of leaf photosynthetic products during flowering and boll setting stage in high yield cotton of Xinjiang. Cotton Science, 2009, 21(5): 371-377. (in Chinese)
[33] 于康珂, 刘源, 李亚明, 孙宁宁, 詹静, 尤东玲, 牛丽, 李潮海, 刘天学. 玉米花期耐高温品种的筛选与综合评价. 玉米科学, 2016, 24(2): 62-71.
YU K K, LIU Y, LI Y M, SUN N N, ZHAN J, YOU D L, NIU L, LI C H, LIU T X. Screening and comprehensive evaluation of heat tolerance of maize hybrids in flowering stage. Journal of Maize Sciences, 2016, 24(2): 62-71. (in Chinese)
[34] 高英波, 张慧, 单晶, 薛艳芳, 钱欣, 代红翠, 刘开昌, 李宗新. 吐丝前高温胁迫对不同耐热型夏玉米产量及穗发育特征的影响. 中国农业科学, 2020, 53(19): 3954-3963.
GAO Y B, ZHANG H, SHAN J, XUE Y F, QIAN X, DAI H C, LIU K C, LI Z X. Effects of pre-silking high temperature stress on yield and ear development characteristics of different heat-resistant summer maize cultivars. Scientia Agricultura Sinica, 2020, 53(19): 3954-3963. (in Chinese)
[35] 许振柱, 周广胜. 全球变化下植物的碳氮关系及其环境调节研究进展-从分子到生态系统. 植物生态学报, 2007, 31(4): 738-747.
XU Z Z, ZHOU G S. Relationship between carbon and nitrogen and environmental regulation in plants under global change-From molecule to ecosystem. Journal of Plant Ecology, 2007, 31(4): 738-747. (in Chinese)
Effects of high temperature stress in different periods on dry matter production and grain yield of summer maize

1State Key Laboratory of Crop Biology/College of Agronomy, Shandong Agricultural University, Taian 271018, Shandong;2Shandong Seed Management station, Ji’nan 250100
【Objective】Frequent, recurrent and prolonged high temperature stress had significant effects on grain yield of summer maize in Huang-Huai-Hai region. In this study, we investigated the effects of high temperature stress at the V12 stage and VT stage on leaf photosynthetic characteristics, carbon assimilate accumulation, distribution and grain yield of maize varieties with different heat tolerance.【Method】In this study, heat resistant maize variety Zhengdan 958 (ZD958) and heat sensitive maize variety Xianyu 335 (XY335) were used as materials. The normal temperature treatments (day 32℃12 h /night 22℃12 h) were set as the control at the same time. High temperature greenhouse equipped with automatic temperature and humidity control facilities was used to simulate the effect of natural field high temperature, the high temperature stress treatments (day 38℃12 h/night 28℃12 h) were set at V12 and VT stage, respectively. The dynamic characteristics of leaf area index (LAI), carbon metabolism enzyme activities, photosynthetic rate and carbon assimilate accumulation and allocation were compared after high temperature stress, aimed to determine the response mechanism of dry matter production performance and grain yield to high temperature stress. 【Result】After high temperature stress, LAI, carbon metabolism enzyme activities, net photosynthetic rate and dry matter accumulation of two cultivars were significantly decreased. LAI, RuBP carboxylase activity, PEP carboxylase activity, net photosynthetic rate and dry matter accumulation of ZD958 and XY335 decreased by 2.98%-4.21%, 40.38%-54.46%, 16.88%-30.60%, 18.14%-25.49%, 12.83%-19.38% and 3.80%-5.07%, 56.56%-76.16%, 26.33%-33.66%, 22.37%-34.62%, 22.07%-26.72%, respectively. The decrease range of high temperature stress in VT stage was larger than that in V12 stage. After high temperature stress, transpiration rate of summer maize leaves increased, while leaf water use efficiency decreased significantly. Under high temperature stress,13C assimilation of ZD958 and XY335 decreased by 18.48% and 22.82%, respectively, and the proportion of13C assimilation in grains decreased. The high temperature stress significantly decreased grain number per spike and grain yield, although 1000 grain weight increased slightly. Compared to the optimum temperature, after V12 high temperature stress, the grain number per spike and yield of ZD958 decreased by 62.53% and 45.87%. After VT high temperature stress, grain number per spike and yield decreased by 70.53% and 66.89%. After V12 high temperature stress, the grain number per spike and yield of XY335 decreased by 70.50% and 62.87%. After VT high temperature stress, grain number per spike and yield decreased by 85.41% and 80.61%. The decrease range of high temperature stress in VT stage was larger than that in V12 stage, and XY335 decreased more than ZD958. 【Conclusion】The high temperature stress reduced LAI, RuBP carboxylase and PEP carboxylase activities, and significantly reduced photosynthetic rate and dry matter production performance of summer maize. Under high temperature stress, the grain number per spike decreased significantly, which inhibited the transportation of carbohydrate from leaf and stem to grain, resulting in lower grain yield. The effects of high temperature stress on dry matter performance and grain yield of summer maize in VT stage was significantly greater than that in V12 stage. The decrease of heat sensitive variety XY335 was significantly greater than that of heat resistant variety ZD958 in two periods.
summer maize; high temperature stress; carbon metabolism enzyme activity; carbon assimilate accumulation and distribution; yield

10.3864/j.issn.0578-1752.2022.19.003
2021-12-05;
2022-06-29
国家自然科学基金(32071959)、山东省重点研发计划(LJNY202103)、山东省现代农业产业技术体系(SDAIT02-08)、国家重点研发计划(2016YFD0300106)
张川,E-mail:zhangc9813@126.com。通信作者刘存辉,E-mail:cunhuiliu@163.com。通信作者刘鹏,E-mail:liupengsdau@126.com
(责任编辑 杨鑫浩)
