喀斯特退化天坑不同坡向植物群落系统发育与功能性状结构
2022-10-27刘橼锰张永永孙晓瑞
税 伟,冯 洁,李 慧,江 聪,孙 祥,刘橼锰,张永永,孙晓瑞
1 福州大学环境与安全工程学院, 福州 350116 2 福州大学空间数据挖掘与信息共享教育部重点实验室, 福州 350116 3 北京大学城市与环境学院, 北京 100871
喀斯特天坑是一种具有垂直峭壁的圈闭化“负地形”,坑内形成异于地表的局域小生境,为某些植物的生长繁殖提供了“避难所”,坑内形成的独特地下森林,是一座天然的物种保护库[1—2]。由于天坑不同区域受垂直峭壁影响的程度不同,内部局域环境差异大,植物群落分异明显。在原生天坑坑壁梯度上存在明显的生态交错带[3],在退化天坑底部的草地群落呈圈层分布,群落物种丰富度和多样性呈“逆洼地效应”[4],其倒石坡乔灌植物物种多样性高于坑外[5],不同坡向植物群落类型分异明显,阳坡以壳斗科等耐旱型植物群落为主要建群种,而阴坡随着演替的进行,壳斗科等耐旱型植物优势逐渐减弱[6]。目前天坑植物群落的研究多集中在物种多样性特征、植物群落的空间特征、不同环境梯度下植物功能性状特征等,而结合系统发育和功能性状研究天坑植物群落构建的则鲜有。
群落构建机制受多种条件影响[7],不同群落受地方生物因素和非生物因素影响,导致群落类型和演替速度的不同,其系统发育结构也发生改变[8]。新几内亚低地雨林的次生林系统发育结构聚集,原始林系统发育结构发散,表明随着演替的进行群落构建的主导因素由生境过滤转向生物间的相互作用[9],然而哥斯达黎加东北部演替早期的热带低地潮湿雨林群落的系统发育结构发散,竞争排斥影响较大[10]。可见群落的系统发育结构受研究区域的环境因素影响而不同。植物群落构建在小尺度上的研究,地形和土壤因子是非常重要的影响因素,坡向是小尺度研究中的一个典型梯度,对植物群落构建有重要意义[11—12]。不同坡向其光照强度、土壤温湿度、土壤营养物质含量差异大[13—14],会影响植物群落的物种组成、物种多样性、功能性状多样性等[15],导致其群落构建机制可能会发生变化。随着Blomberg等[16]提出系统发育信号检验的发展,系统发育在群落构建中的研究日渐增多。系统发育的方法可以很好地构建微域的物种库,揭示某地区的物种进化历史、过去物种的生态位进化以及当代物种的共存机制[17]。在天坑圈闭化生境中,发育了多个坡向的倒石坡,具有独特生境的天坑内部又形成了异质性的坡向小生境,因此对坡向植物多样性维持机制的研究,探讨环境作用和生物作用对不同坡向植物群落构建的影响,有助于揭示天坑生物多样性保护库价值,为喀斯特地区植被恢复提供依据。
功能性状是植物与环境之间长期相互影响的结果[6],能够指示环境变化,反映植物生态策略,对揭示植物群落构建机制有重要作用[18—19],本文以云南沾益深陷塘天坑为例,结合植物功能性状和系统发育的方法研究植物群落构建,分析天坑坡向植物群落的功能性状是否具有显著的系统发育信号,通过不同坡向的植物群落系统发育结构和功能性状结构,量化环境过滤、竞争排斥和随机过程在天坑坡向植物群落构建中的作用,为天坑植物多样性的保护提供科学的参考。
1 材料和方法
1.1 区域概况和样方设置
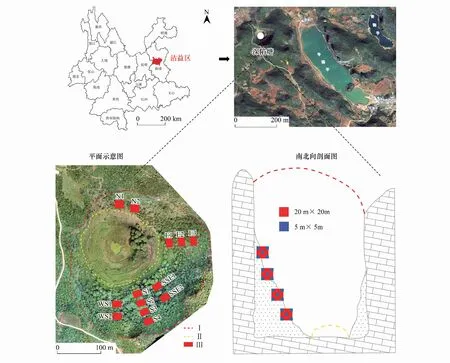
图1 天坑区位图及样方布设示意图Fig.1 Location map of Tiankeng and schematic diagram of plot layoutN: 北 North; E: 东 East; SSE: 东南偏南 south-southeast.; S: 南 South; WS: 西南 southwest; Ⅰ: 天坑上部边界Upper boundary of tiankeng; Ⅱ: 坑坡下部边界Lower boundary of tiankeng slope; Ⅲ: 天坑乔木样方Arbor quadrat in tiankeng
沾益天坑群位于云南省曲靖市(图1),地理坐标在25°35′—25°57′N, 103°29′—103°39′E之间,其中包含原生天坑和退化天坑,具有丰富的物种多样性[20]。该地区属于亚热带高原季风气候,昼夜温差较大,年均温在13.8—14℃,太阳辐射能年总量为518.2 kJ/cm2,天坑群内土壤主要有红壤、黄棕壤、紫色土及石灰土等。经过对云南喀斯特天坑群的调查,本研究以典型的中度退化天坑——深陷塘天坑为研究对象。深陷塘坑底海拔2028—2031 m,坑口长径421.9 m,短径348.7 m,坑深148.7 m[5]。西侧为垂直峭壁,北侧为陡峭退化坑壁,是天坑的入口,植被以稀疏耐旱的灌木为主;东侧上部为垂直峭壁,下部为倒石坡,倒石坡上分布着乔灌相间的植被;南侧是坑内最大的倒石坡,植被以乔木为主。南侧倒石坡是坑内阴坡,生境条件较好,东坡次之,北坡作为阳坡,生境条件最差。根据坑内坡长和植物生长分布的实际情况,在南侧、西南侧、南南东侧、东侧和北侧共布设13个20 m×20 m的乔木样方,在每个乔木样方内按照“梅花型”设置5个5 m×5 m的灌木样方,共计65个灌木样方,分别调查记录每个样方内植物的物种名、株高、胸径、多度,利用GPS记录经纬度和海拔数据。物种多样性对植物群落有重要影响[21],通过计算分析可得不同坡向植物群落α多样性,其中Margalef丰富度为南坡>东坡>北坡,且差异显著,而不同坡向植物群落的物种多样性和均匀度差异不显著。
1.2 功能性状的采集和测定
根据全球植物功能性状标准化测量手册[22],在样方内使用高枝剪采摘每个物种不同植株不同方向的长势良好的3—5片叶子。使用精度为0.02 mm的电子数显卡尺测叶子主脉一侧0.25 cm处,记录三组数据求平均值;使用叶面积仪记录每一片叶子的叶长(LL)、叶宽(LW)、长宽比(LWR)和叶面积(LA);鲜叶采用精度为0.001 g电子天平称量鲜重,叶片带回实验室后放入设置为80℃的烘干箱48小时至恒重,然后称量其干重。比叶面积(SLA)为叶面积与叶干重的比值;叶干物质含量(LDMC)为叶干重与叶鲜重的比值。
1.3 土壤因子的测定
土壤理化性质指标依托于福建省农科院土壤肥料研究所测定。土壤含水量(SWC, %)采用烘干法,用环刀采集深度为0—20 cm土壤样品,称量其湿重M,在温度设置为105℃的烘箱中6—8 h,烘干至恒重Ms,计算土壤含水量SWC=(M-Ms)/M;用土壤酸度计测定pH,用重铬钾氧化法测定土壤有机质(SOM),碱解扩散法测定土壤碱解氮(AN),土壤全氮(Total nitrogen, TN)的测定采用半微量开氏法,用钼锑抗比色法测定全磷(TP)和有效磷(AP),用火焰光度计法测定全钾(TK)和速效钾(AK)。
1.4 功能性状的系统发育信号检测
本文根据Blomberg等[16]提出的K值检验法分析系统发育信号。K=1定义为Brownian motion的进化模型;K>1,表明功能性状具有比Brownian motion模型进化更强的系统发育信号;反之,K<1,表明功能性状的系统发育信号较弱。通过实际K值与零模型的比较,进而判断功能性状系统发育信号的显著性。当系统发育树分支末端物种随机置换999次生成的零模型K值小于实际K值(P<0.05),功能性状具有显著的系统发育信号,否则说明系统发育信号不显著[15]。
1.5 群落系统发育和功能性状结构
系统发育树的构建是采用物种的科属信息经过R语言的“plantlist”程序包处理,输入网页版phylomatic软件中,得到物种的系统发育的拓扑结构,该结构是以被子植物分类系统Ⅲ(APGⅢ, Angiosperm Phylogeny Grup Ⅲ)为基础构建。然后经过phylocom软件的BLADJ算法,结合分子及化石定年数据拟合系统发育拓扑结构的分支结构,采用R语言“picante”程序包计算得到系统发育树。本文研究中的净亲缘关系指数(Net relatedness index, NRI)是群落中所有物种的平均系统发育距离(Mean pairwise distance, MPD)的标准效应值,最近亲缘关系指数(Nearest taxon index, NTI)是平均最近相邻谱系距离(Mean nearest taxon distance, MNTD)的标准效应值。根据调查样方群落得到实际的MPD或MNTD,把物种随机置换999次构建零模型,得到随机状态下的MPD或MNTD,经过标准化得到NRI或NTI[23—24]。
采用平均配对性状距离(Mean pairwise trait distance, traitMPD)和平均最近相邻性状距离(Mean nearest taxon trait distance, traitMNTD)计算功能性状结构。TraitSESMPD(Standardized mean pairwise trait distance)和traitSESMNT(Standardized mean nearest taxon trait distance)是基于功能性状聚类树计算求得。
系统发育指数(NRI和NTI)和功能性状结构指数(traitSESMPD和traitSESMNTD)等于0时,表示谱系结构和功能性状结构自由扩散;指数大于0时,表示谱系结构和功能性状结构聚集;指数小于0时,表示谱系结构和功能性状结构发散。
式中,MPDobs、MNTDobs、traitMPDobs、traitMNTDobs是实际观测值;MPDrand、MNTDrand、traitMPDrand、traitMNTDrand是零模型下随机零群落MPD、MNTD、traitMPD、traitMNTD的值;mean(MPDrand)、mean(MNTDrand)、mean(traitMPDrand)、mean(traitMNTDrand)是通过零模型计算的平均值;sd(MPDrand)、sd(MNTDrand)、sd(traitMPDrand)、sd(traitMNTDrand)是零模型下随机零群落的各指数的标准差。
1.6 数据处理
植物功能性状的种内变异是物种性状的标准差与平均值的比值。重要值是用综合性数量指标来反映植物物种在群落中的相对重要性[25],本文以重要值的大小作为群落主要优势物种的测度指标[26—27]。乔木层重要值=(相对密度+相对频度+相对显著度)/3,灌木层重要值=(相对密度+相对频度+相对显著度)/3。根据单因素方差分析判断不同坡向的系统发育指数、功能性状结构指数的显著性差异。数据整理和多样性指数计算在Excel 2016中进行,系统发育和功能性状结构指数的计算在R 4.0.2中实现,相关制图均在R 4.0.2和Origin 2018中绘制。
2 结果与分析
2.1 不同坡向土壤环境差异
深陷塘天坑的土壤有机质、有效磷和土壤含水量从北坡到南坡逐渐增大,土壤pH从北坡到南坡逐渐减小。全氮含量在北坡(2.03 g/kg)最低,而东坡和南坡差异不大;土壤碱解氮含量东坡(185.2 mg/kg)最高,北坡(105.3 mg/kg)最低;土壤全磷在北坡含量(0.67 g/kg)最高,东坡和南坡相差不大;土壤全钾在南坡含量(14.33 g/kg)最高,东坡含量(5.10 g/kg)最低;土壤速效钾在北坡含量(139.50 mg/kg)最高,东坡含量(87.93 mg/kg)最低(表1)。从北坡—东坡—南坡,土壤含水量及土壤营养物质含量有逐渐增大的趋势,东坡土壤含水量和土壤营养物质介于北坡和南坡之间,且变异系数较大,可能是因为东坡处于阴坡和阳坡的交界处,造成不同坡位样方的土壤环境差异较大。
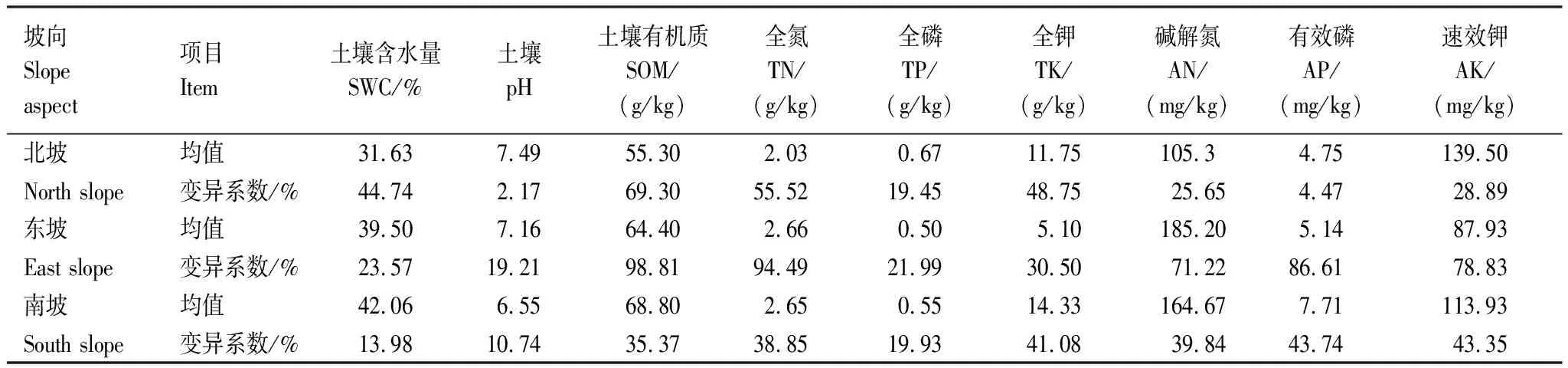
表1 深陷塘不同坡向土壤环境因子
2.2 不同坡向植物群落组成
本研究共调查26科40属47种物种(图2),其中北坡15种,东坡20种,南坡45种。本文取各个坡向重要值最大的前三个物种来表征该坡向的优势种,因此北坡优势种有黄背栎、长圆叶梾木(Swidaoblonga)、球核荚蒾(Viburnumpropinquum),东坡优势种有黄背栎、栓皮栎(Quercusvariabilis)、清香木 (Pistaciaweinmannifolia),南坡优势种有头状四照花 (Dendrobenthamiacapitata)、青冈、构树(Broussonetiapapyrifera)(表2)。
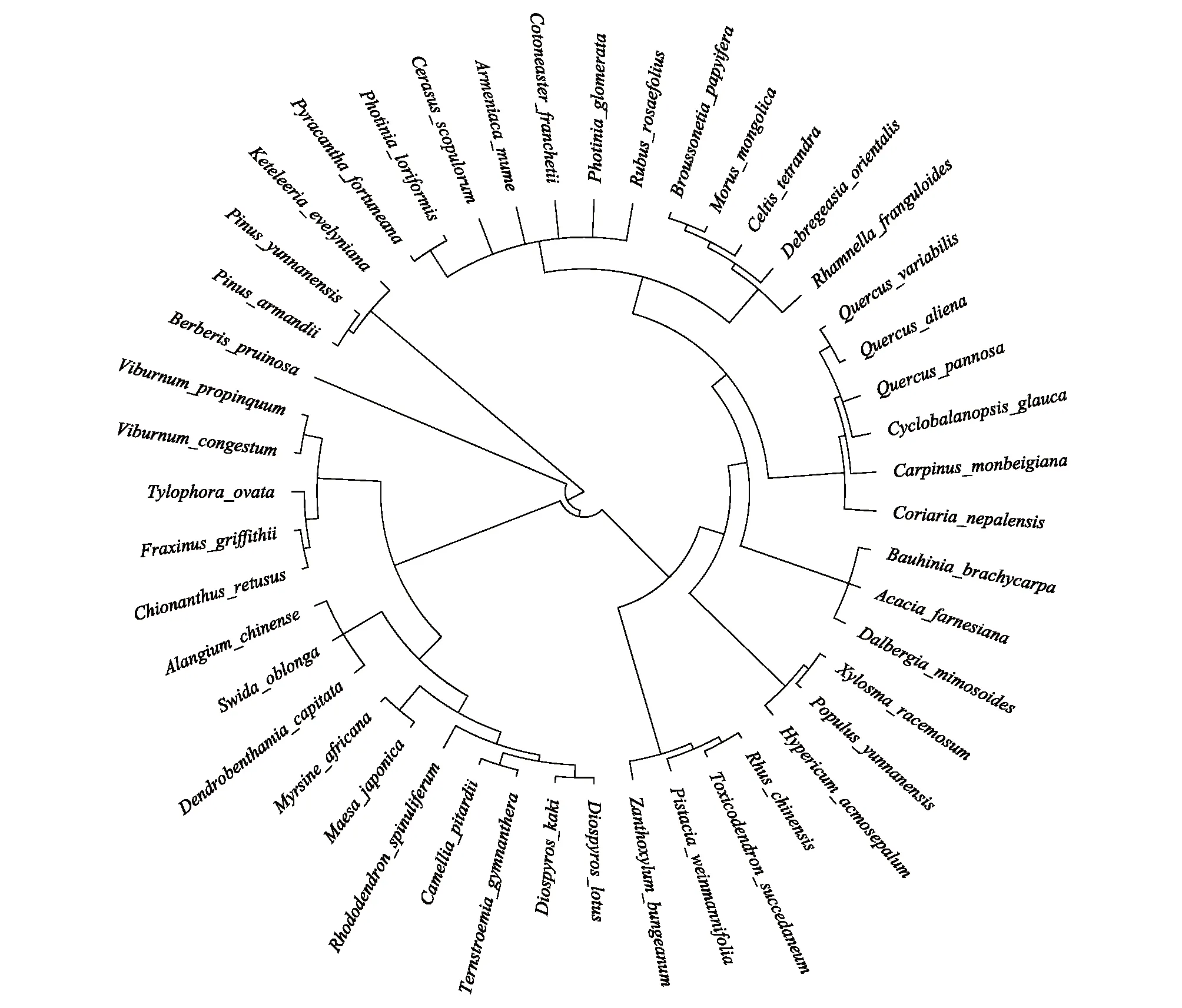
图2 系统发育树Fig.2 Phylogenetic treeBerberis pruinosa: 粉叶小檗; Toxicodendron succedaneum: 野漆; Rhus chinensis: 盐肤木; Pistacia weinmanniifolia: 清香木; Zanthoxylum bungeanum: 花椒; Hypericum acmosepalum: 尖萼金丝桃; Populus yunnanensis: 滇杨; Xylosma racemosum: 柞木; Dalbergia mimosoides: 象鼻藤; Acacia farnesiana: 金合欢; Bauhinia brachycarpa: 鞍叶羊蹄甲; Coriaria nepalensis: 马桑; Carpinus monbeigiana: 云南鹅耳枥; Quercus aliena: 槲栎; Quercus variabilis: 栓皮栎; Cyclobalanopsis glauca: 青冈; Quercus pannosa: 黄背栎; Rhamnella franguloides: 猫乳; Celtis tetrandra: 四蕊朴; Morus mongolica: 蒙桑; Broussonetia papyrifera: 构树; Debregeasia orientalis: 水麻; Photinia loriformis: 带叶石楠; Pyracantha fortuneana: 火棘; Cotoneaster franchetii: 西南栒子; Armeniaca mume: 梅; Photinia glomerata: 球花石楠; Cerasus scopulorum: 崖樱桃; Fraxinus griffithii: 光蜡树; Chionanthus retusus: 流苏树; Tylophora ovata: 娃儿藤;Rubus rosaefolius: 空心泡; Viburnum propinquum: 球核荚蒾; Viburnum congestum: 密花荚蒾; Rhododendron spinuliferum: 爆杖花; Camellia pitardii: 西南红山茶; Ternstroemia gymnanthera: 厚皮香; Diospyros kaki: 野柿; Diospyros lotus: 君迁子; Myrsine africana: 铁仔; Maesa japonica: 杜茎山; Alangium chinense: 八角枫; Swida oblonga: 长圆叶梾木; Dendrobenthamia capitata: 头状四照花; Pinus yunnanensis: 云南松; Pinus armandii: 华山松; Keteleeria evelyniana: 云南油杉
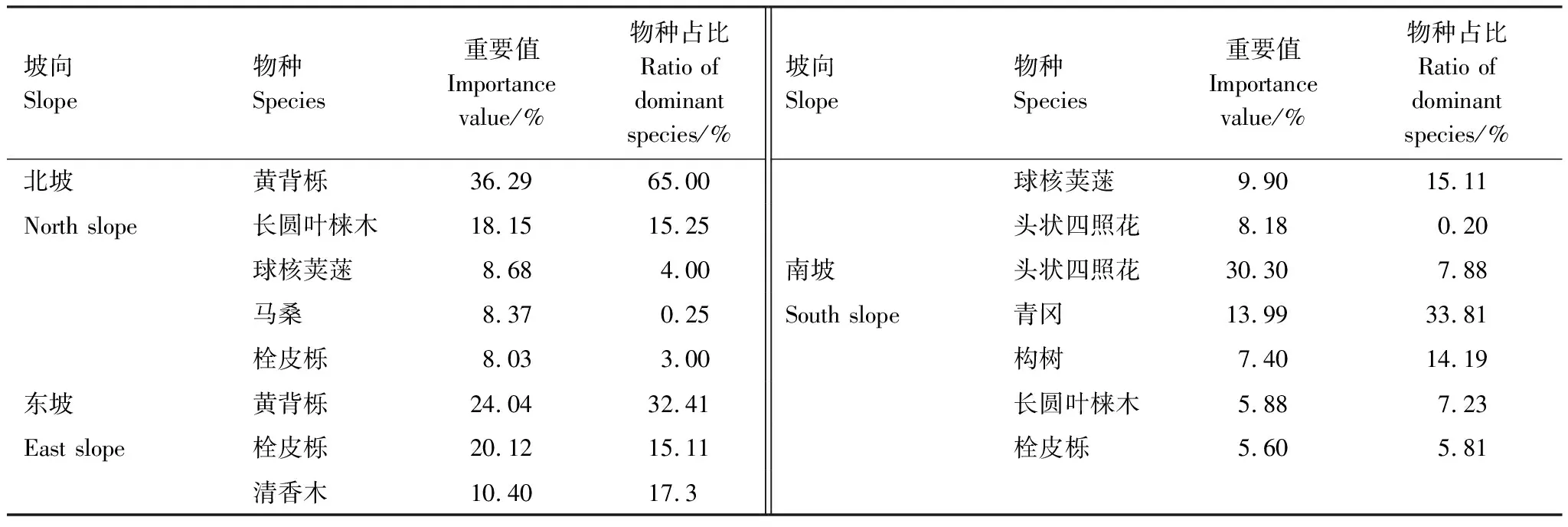
表2 不同坡向重要值前5的物种
2.3 退化天坑不同坡向优势种功能性状变异特征
深陷塘各坡向优势种性状总体变异程度中等,不同性状之间存在差异。从表3来看,在北坡,黄背栎功能性状变异程度最大的是叶面积,最小是长宽比,而长圆叶梾木叶厚度变异程度最大,比叶面积变异程度最小,球核荚蒾也是叶面积变异程度最大,但叶宽变异程度最小;东坡黄背栎和栓皮栎都是叶面积变异程度最大,叶长最小,清香木比叶面积变异程度最大,叶宽最小;南坡头照四状花和构树都是叶干重变异程度最大,叶长最小,而青冈叶干物质含量变异程度最大,叶宽最小。三个坡向的优势种中,东坡清香木叶面积变异程度最大(54.26%),而变异程度最小(8.66%)的则是南坡青冈的叶宽。整体上,南坡优势物种变异程度最大,东坡次之,北坡优势物种变异程度最小。
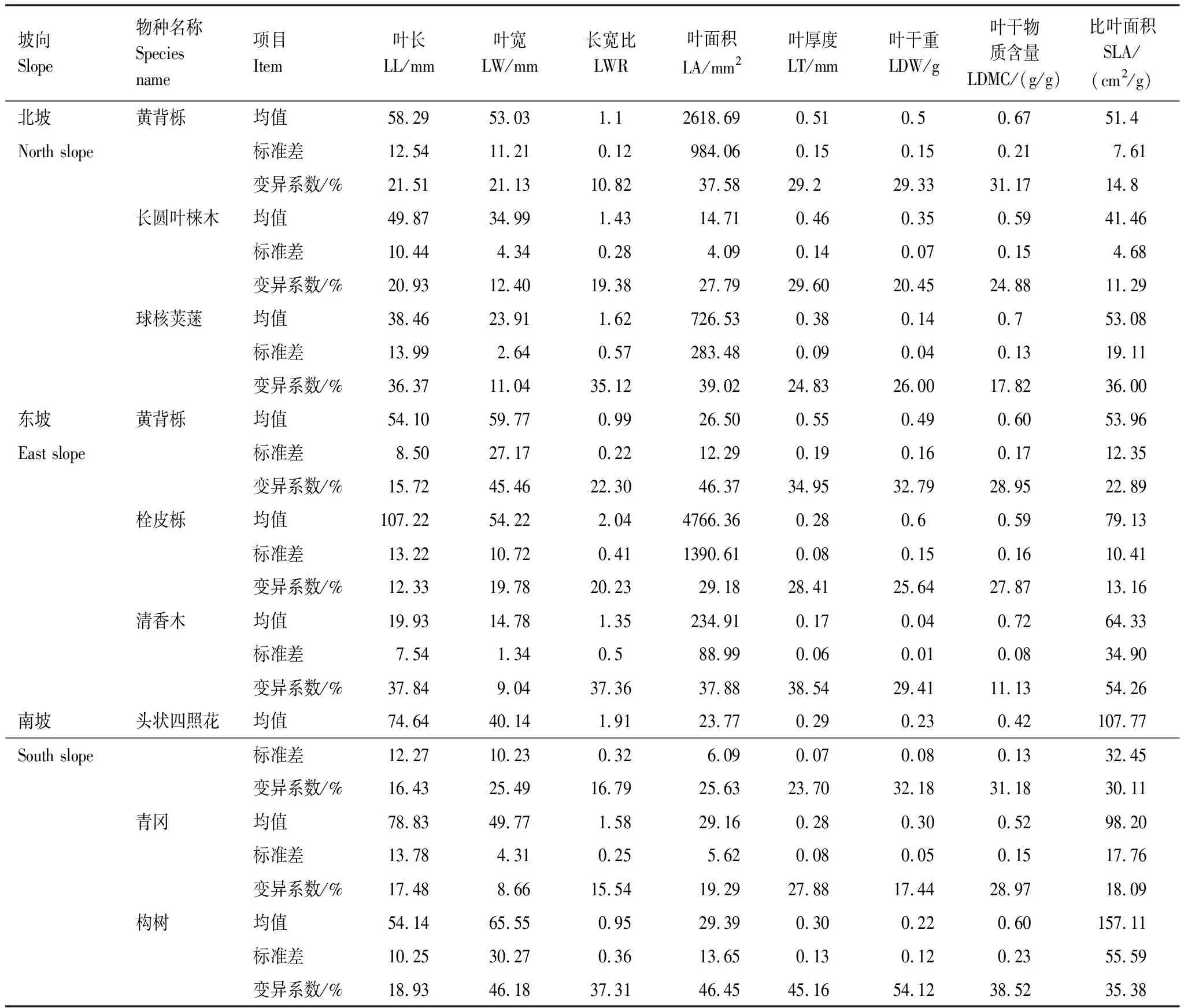
表3 深陷塘不同坡向优势物种功能性状变异特征
2.4 退化天坑不同坡向植物功能性状的系统发育信号检验
植物群落构建过程中聚集是环境过滤占主导作用,而发散主要由竞争排斥作用主导。喀斯特天坑深陷塘坡向植物群落的叶长、叶面积、叶干重、叶干物质含量、最大树高的K值小于1,说明功能性状表现出的系统发育信号强度弱,这5种功能性状都没有较强的系统发育保守性,性状在进化过程中受生境影响更大;而叶宽、比叶面积、长宽比、叶厚度的K值大于1,说明功能性状表现出的系统发育信号强,受遗传因素影响显著(P<0.05)(表4),表明谱系距离近的物种,功能性状的相似度高。
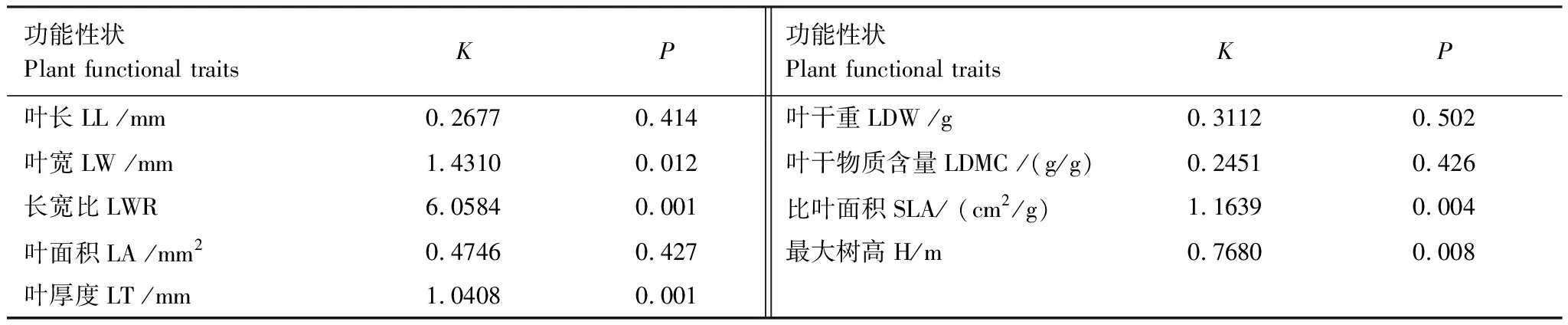
表4 深陷塘坡向植物功能性状系统发育信号检验
2.5 退化天坑不同坡向群落的系统发育和功能性状结构的差异
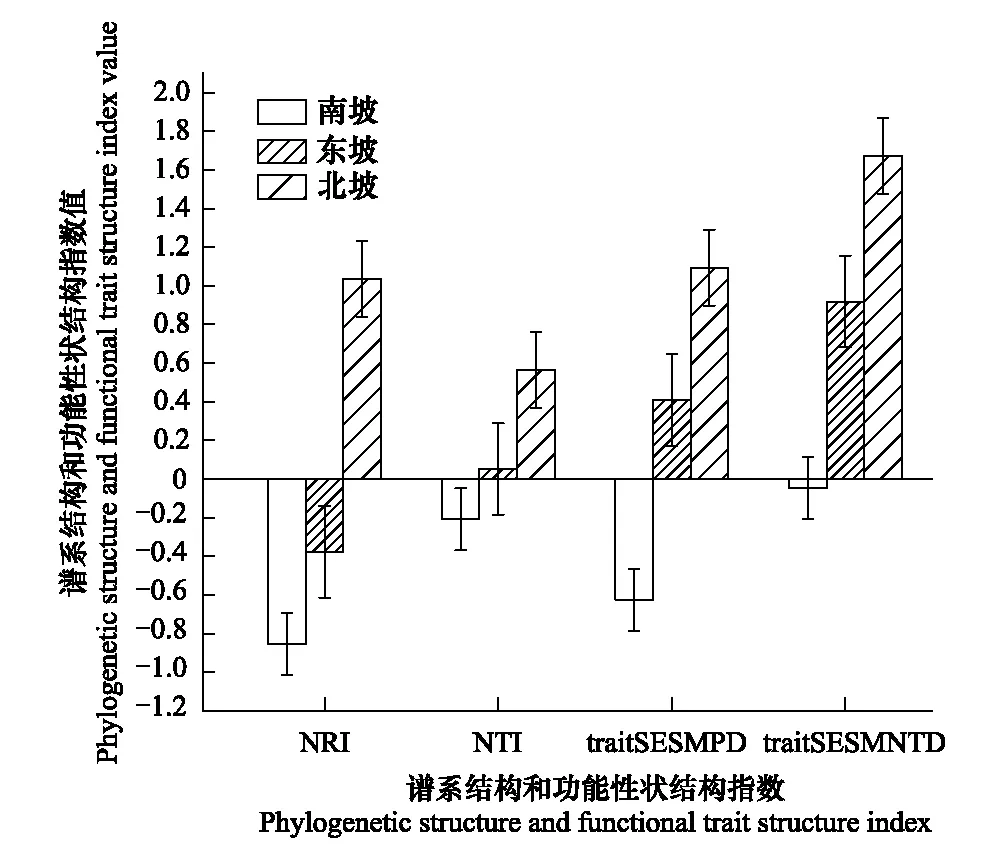
图3 不同坡向的谱系结构和功能性状结构 Fig.3 Phylogenetic structures and functional structures of different slope NRI: 净种间亲缘关系指数 Net relatedness index; NTI: 最近种间亲缘关系指数 Nearest taxon index; trait SESMPD: 标准化种间平均配对性状距离 Standardized mean pairwise trait distance; trait SESMNTD: 标准化种间平均最近相邻性状距离 Standardized mean nearest taxon trait distance
喀斯特天坑深陷塘北坡植物群落NRI、NTI和traitSESMPD、traitSESMNTD均大于0,表明系统发育结构和功能性状结构均聚集。而南坡植物群落的NRI、NTI指数和traitSESMPD、traitSESMNTD指数均小于0,表明功能性状结构和功能性状结构均发散。东坡植物群落系统发育两指数结果不一致,NRI指数小于0,说明系统发育结构发散;NTI指数大于0,表明系统发育结构聚集;功能性状结构指数traitSESMPD和traitSESMNTD均是大于0,说明功能性状结构聚集。NRI、NTI和traitSESMPD、traitSESMNTD沿着坡向变化(北坡-东坡-南坡)存在相同的变化趋势,即随着坡向的变化系统发育结构和功能性状结构都是从集聚逐渐转变为发散(图3)。
3 讨论
3.1 天坑植物群落系统发育与功能性状结构指数随坡向的变化特征
深陷塘天坑受西面垂直峭壁的遮挡影响,不同方位日辐射量不同,坑内湿热环境异于地表,坑内不同坡向生境中植物群落类型不同,其群落构建机制过程存在差异。江聪等[28]在研究紫茎泽兰(Eupatoriumadenophorum)入侵巴家陷塘天坑时发现,紫茎泽兰在坑坡灌草地较为集中,主要分布在西南坡,而东北坡基本没有分布,从西南坡到东北坡,植物群落由阳生植物演替为阴生植物。本研究表明,深陷塘各坡向系统发育指数和功能性状结构指数有相同的变化趋势,即从北坡到南坡,系统发育指数减小,功能性状结构指数也减小。由图3可知,北坡的植物群落的系统发育结构和功能性状结构均为聚集,而南坡植物群落的系统发育结构和功能性状结构均为发散,且从北坡到南坡发散程度逐渐上升。在东坡,两种功能性状结构指数均为聚集,而两种系统发育指数结果不同,NRI指数表明东坡系统发育结构发散,NTI指数表明东坡系统发育结构聚集。车应弟等[29]在分析亚高寒草甸群落构建时发现西坡两种系统发育指数也不一致。东坡两种系统发育指数结果不同,可能有两种解释:一是两种系统发育指数侧重点不同,NRI亲缘指数是基于群落整体水平的,反映的是整个系统发育树的发育模式,而NTI亲缘关系是基于系统进化树末端的最近的系统发育关系[30];二是由于深陷塘西面是垂直峭壁,东坡属于北坡和南坡的坡向过渡地带,环境条件和种间竞争共同影响了该坡的群落构建。受垂直峭壁的遮挡影响,阴坡和阳坡生境分异明显,深陷塘北坡为阳坡,土壤含水量低,土壤养分含量少,植被主要以耐旱的黄背栎等灌木为主,环境对物种的生长影响较大,这与刘旻霞等[31]的研究结果一致,即环境过滤是阳坡群落构建的主要作用;而深陷塘南坡为阴坡,土层深,土壤含水量高,养分丰富,总生态位宽,植被以头状四照花和构树等阔叶乔木为主,物种多样性高,加剧了一些功能性状相似的物种因生态位重叠范围较大而引起的竞争,这与朱云云等[32]的研究结果一致,即竞争排斥是阴坡群落构建的主要作用;东坡作为北坡和南坡的过渡地带,乔灌混杂,受环境过滤和竞争排斥的共同作用。从北坡到东坡,再到南坡的植物群落构建过程中,环境过滤作用逐渐减小,竞争排斥作用逐渐增大,植被群落由耐旱性的灌木,逐渐演替为高大的乔木,这三个坡向植物组成构成了一个群落演替系列。
3.2 天坑坡向植物群落构建中的生物过滤机制
群落构建在生物过滤和环境过滤的长期影响下,物种逐渐趋于局域环境物种库的过程[33],其中种间竞争和种内性状变异是生物过滤的主要生态学过程[34]。种间竞争是群落结构和植物形态的重要影响因素之一,导致物种间形成生态排序,竞争能力弱的物种逐渐从群落中淘汰,最后形成的共存物种有均等的适合度[35—36],植物群落逐渐趋于稳定。种内变异多是因为植物功能性状的生态适应性策略,植物功能性状的变异受环境因子(气候、地形、土壤因子等)的影响,但功能性状之间的相互作用也是重要因素[34]。Ackerly等[37]的研究表明由于南北坡光照、湿度等存在差异,导致不同坡向植物的种内功能性状出现变异。本研究中,北坡是阳坡,光照强度大,土壤水分含量低,营养物质相对贫瘠,恶劣的环境使得北坡植物群落的谱系结构聚集,促进了植物功能性状趋同进化。而南坡是阴坡,土壤水分含量高,营养物质丰富,植物群落复杂,属于针阔混交林。林下灌丛茂密,植物群落间竞争排斥占主导作用,导致植物功能性状趋异进化。南坡优势种通过增大种内变异程度来缓冲物种间的竞争作用,提高南坡植物群落的稳定性。这与刘旻霞等[38]和盘远方等[39]的研究结果一致,表明由于阴坡和阳坡的生境差异,导致坡向变化是植物功能性状结构存在差异的主要原因。东坡是连接北坡与南坡的生态交错带[40],生境条件次于南坡,却高于北坡;东坡植物的叶长、叶宽、长宽比、叶面积和叶干重有较小的种内变异,可能是因为东坡以黄背栎和栓皮栎为主要优势建群种,其多度较大,种内变异程度与物种的多度有负相关关系[34],因为常见种在资源获取的能力方面差异不大,所以种内变异程度较小。
4 结论
本文通过对深陷塘退化天坑不同坡向植物功能性状和系统发育的研究发现:深陷塘天坑不同坡向的系统发育和功能性状结构具有不一致性,但总体是受生态位过程主导。环境过滤是北坡物种共存的主要机制,竞争排斥是南坡物种共存的主要机制,对于东坡这种生境过渡区域,受到环境过滤和竞争排斥的共同作用。坑内植物功能性状受遗传因子和生境的共同作用,在不同的坡向梯度生境中,采取不同的生存策略,揭示了坡向生境对植物群落功能性状的影响。未来可以结合土壤、光照、水分等环境因子,进一步量化环境因子在不同坡向植物群落构建中的作用,为云南喀斯特天坑植物资源保护和繁育、天坑群生态保护和恢复工作提供理论支撑。
