濒危植物小黄花茶种群数量结构与生存潜力分析
2022-10-17白小节申开平穆君翁涛臧丽鹏任闻达韩勖李青谭淇毓何跃军
白小节, 申开平, 穆君, 翁涛, 臧丽鹏, 任闻达, 韩勖, 李青, 谭淇毓, 何跃军*
濒危植物小黄花茶种群数量结构与生存潜力分析
白小节1, 申开平2, 穆君2, 翁涛2, 臧丽鹏2, 任闻达2, 韩勖2, 李青2, 谭淇毓2, 何跃军2*
(1. 贵州省赤水桫椤国家级自然保护区管理局,贵州 赤水 564700;2. 贵州大学林学院,森林生态研究中心,贵阳 550025)
为了解濒危植物小黄花茶()的生长特征和生存状况,对贵州赤水市国家级桫椤自然保护区内小黄花茶进行全域资源清查,采用样方法调查小黄花茶每株植物高度和地径,统计分析了种群数量及结构特征,并采用径级对应龄级,以0.5 cm地径间隔划特定年龄分组,共分Ⅰ~XV径级,绘制静态生命表分析种群生存潜力和发展趋势。结果表明,调查共发现小黄花茶545株,不同小生境中小黄花茶数量差异大,最少24株,最多120株;种群地径小于0.5 cm且高度低于0.5 m个体存活数量极少,平均地径0.5~2 cm维持最大种群数量;Ⅰ~Ⅱ龄级(0~1.0 cm)种群死亡率和消失率急剧上升而生命期望值急剧降低,II~XV龄级(≥0.5 cm)种群死亡率和生命期望值趋于平缓,消失率呈现较大波动但总体趋于平缓;种群数量结构和存活曲线表明该种群趋近于Deevey-Ⅰ型,属衰退型种群。小黄花茶种群幼苗数量严重不足,种群自我更新困难,现存种群干扰压力大,种群衰退趋势明显,急需加强种群保育。
小黄花茶;种群结构;静态生命表;存活曲线; 濒危植物
种群数量结构与动态是种群生态学研究核心[1],从种群结构与数量维持角度研究濒危物种的生存潜力,对深入探索物种濒危机制及种群发展动态以及有效保护濒危物种具有重要意义。生命表和存活曲线可反映种群生存状况以及种群与环境间的相互关系[2–4],对世代重叠种群采用静态生命表和存活曲线分析自然种群生存潜力是研究种群结构和发展动态的重要手段。罗西等[5]通过研究濒危植物鹅掌楸()天然种群年龄结构与数量动态及分布格局,辨析了该种群动态;马丹丹等[6]也通过种群年龄结构、生命表、存活曲线等揭示了珍稀濒危植物堇叶紫金牛()种群动态及存活状态。因此,分析珍稀濒危植物种群结构与存活动态对该物种的保护和利用具有重要意义。
张宏达等[7]在贵州西北部赤水金沙公社海拔900~1 060 m的森林或悬崖峭壁上发现1多年生常绿木本灌木新种,其在系统分类上与短柱茶组(sect.)接近,又与金花茶组(sect.)特征完全不同,故命名为小黄花茶组新组(sect.),并将该物种命名为小黄花茶(),属于山茶科(Theaceae)山茶属小黄花茶组。小黄花茶是赤水桫椤国家级保护区旗舰保护植物桫椤的伴生物种,该保护区也是小黄花茶模式物种产地。桫椤()种群及其群落生态的研究受到广泛关注,桫椤群落优势植物种间联结性[8]、桫椤种群结构与分布格局[9]、种群生态位[10]、种内种间竞争[11]等;然而,对桫椤物种的保护,不仅要保护物种本身,更应保护其生存的生境及其伴生物种。
1988年小黄花茶被列为贵州省省级珍稀濒危保护植物[12]。受人为干扰和自然因素影响,小黄花茶小生境破碎且病害严重,种群数量呈快速下降趋势[13],使得该物种面临灭绝的风险。然而,对该物种的研究是极其滞后的,主要集中在对小黄花茶生物学特性[14–15]、空间分布格局[16]、繁殖技术[17]等,而未见对小黄花茶种群数量结构与生存潜力的研究报道。本研究在对赤水桫椤国家级自然保护区小黄花茶资源清查基础上,对小黄花茶群落及种群进行调查,拟通过小黄花茶种群数量结构和生存潜力分析其发展动态,为该物种的预测保护提供理论参考。
1 材料和方法
1.1 样地概况与群落调查
样地位于贵州赤水桫椤国家级自然保护区葫市镇金沙村闷头溪,保护区面积为133 km2,种群集中分布区地理位置为28°28′01″~28°28′56″ N, 105° 57′35″~105°58′47″ E,海拔352~756 m。本研究针对小黄花茶种群集中分布点采用典型样地法进行群落学调查,本次调查涉及闷头溪下河沟(XHG)、观音岩谷(GYYG)、石缸岩(SGY)、大湾子(DWZ)、黄泥沟(HNG)、陈田(CT)、血台子(XTZ)、半坡头(BPT)共8个调查样点,各样点基本信息见表1。调查样地面积20 m×20 m,每个样地分为4个10 m× 10 m样方,共调查32个样方。按乔木层、灌木层和草本层分别记录每个样方内物种的名称、株数、胸径(地径)、冠幅、枝下高等指标。其中,小黄花茶因平均高度和地径较小,故其地径均在离地面5~10 cm处测量,记录每株的高度、冠幅和生存状态等信息。样方内的主要乔木有毛竹()、慈竹()、香樟()、桫椤()和毛桐()等;主要灌木有小黄花茶、桫椤、杜茎山()、水麻()和柃木()等;主要草本植物有板蓝()、竹叶草()、耳蕨()、里白()和四块瓦()等。
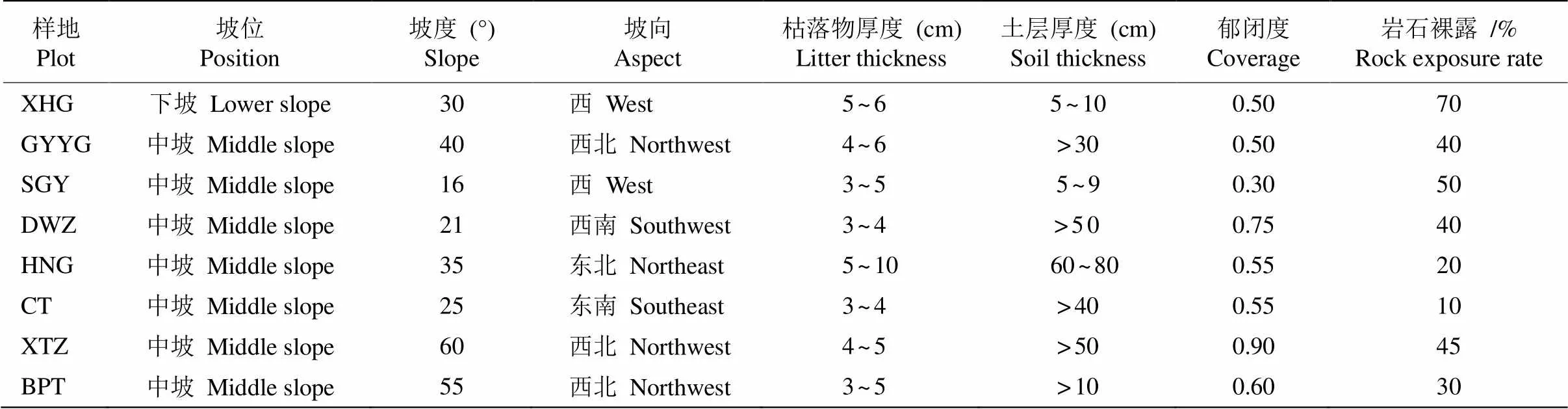
表1 小黄花茶群落样地基本概况
XHG: 下河沟; GYYG: 观音岩谷; SGY: 石缸岩; DWZ: 大湾子; HNG: 黄泥沟; CT: 陈田; XTZ: 血台子; BPT: 半坡头。下图同。
XHG: Xiahegou; GYYG: Guanyinyangu; SGY: Shigangyan; DWZ: Dawanzi; HNG: Huangnigou; CT: Chentian; XTZ: Xuetaizi; BPT: Banpotou. The same is following Figures.
1.2 种群数量统计与生存潜力分析
采用径级对应龄级的时空替代法统计各龄级株数,根据种群的龄级结构编制静态生命表,绘制存活曲线[18];静态生命表中主要参数计算参照江洪[19]的方法。种群存活曲线是将小黄花茶存活个体数量以0.5 cm地径间隔划特定年龄分组,共分Ⅰ~XV径级,按标准化存活数为纵坐标绘制出存活曲线, 参考Deevey[20]提出的存活曲线3种类型,即Deevey-Ⅰ (衰退型种群)、Deevey-Ⅱ (稳定型种群)和Deevey-Ⅲ型(增长型种群),并参考Hett等[21]的检验方法对存活数和龄级关系进行拟合,依据决定系数R和检验值确定最优模型,进而判断种群动态。以死亡率、消失率和期望值为纵坐标,以龄级为横坐标绘制死亡率曲线、消失率曲线和期望值曲线, 预测种群动态变化。
2 结果和分析
2.1 种群数量分布
整体上,调查样地内统计到的小黄花茶数量为545株(图1)。小黄花茶的数量在调查的几个小生境间差异很大,调查的8个样地中,半坡头、陈田、下河沟、血台子样地的小黄花茶数量较多,而观音岩谷、石缸岩、大湾子和黄泥沟样地小黄花茶数量较少。
2.2 种群地径、高度、冠幅和枝下高
由图2可见,下河沟和陈田样地小黄花茶的平均地径较低,分别为2和1.4 cm,而观音岩谷最高,为4.7 cm。高度和地径的变化在几个小生境间表现出一致的规律,下河沟和陈田样地的平均高度较低,分别为2.5和1.6 m;观音岩谷的平均高度最高为5.4 m。在陈田样地的平均冠幅最低,仅为0.7 m2,在观音岩谷的最高,为6 m2。平均枝下高在陈田样地最低,仅为0.4 m,在大湾子样地最高,为1.4 m。总体而言,观音岩谷、大湾子、血台子、黄泥沟、半坡头等样地的小黄花茶地径、高度、冠幅和枝下高均高于下河沟、陈田等样地。

图1 样地内小黄花茶数量
2.3 种群径级和高度级数量特征
由图3可见,小黄花茶的地径呈连续分布,以Ⅱ~Ⅳ级(0.5~2 cm)的个体数量最多,占整个小黄花茶种群的50.14%,而Ⅰ级(<0.5 cm)的个体数量最少,仅占0.55%。此外,小黄花茶的高度也呈连续分布, Ⅳ级(1.5~2 m)的个体数量最多,占整个小黄花茶种群的27.50%,而Ⅰ龄级(<0.5 m)的个体最少,仅占0.83%。由此可见,在小黄花茶种群中,幼苗和老年个体的数量较少,主要由中年个体组成。从年龄结构角度来看,小黄花茶种群幼苗个体数量严重不足,种群自我更新困难,年龄结构总体处于衰退状态。

图2 小黄花茶种群的生长结构
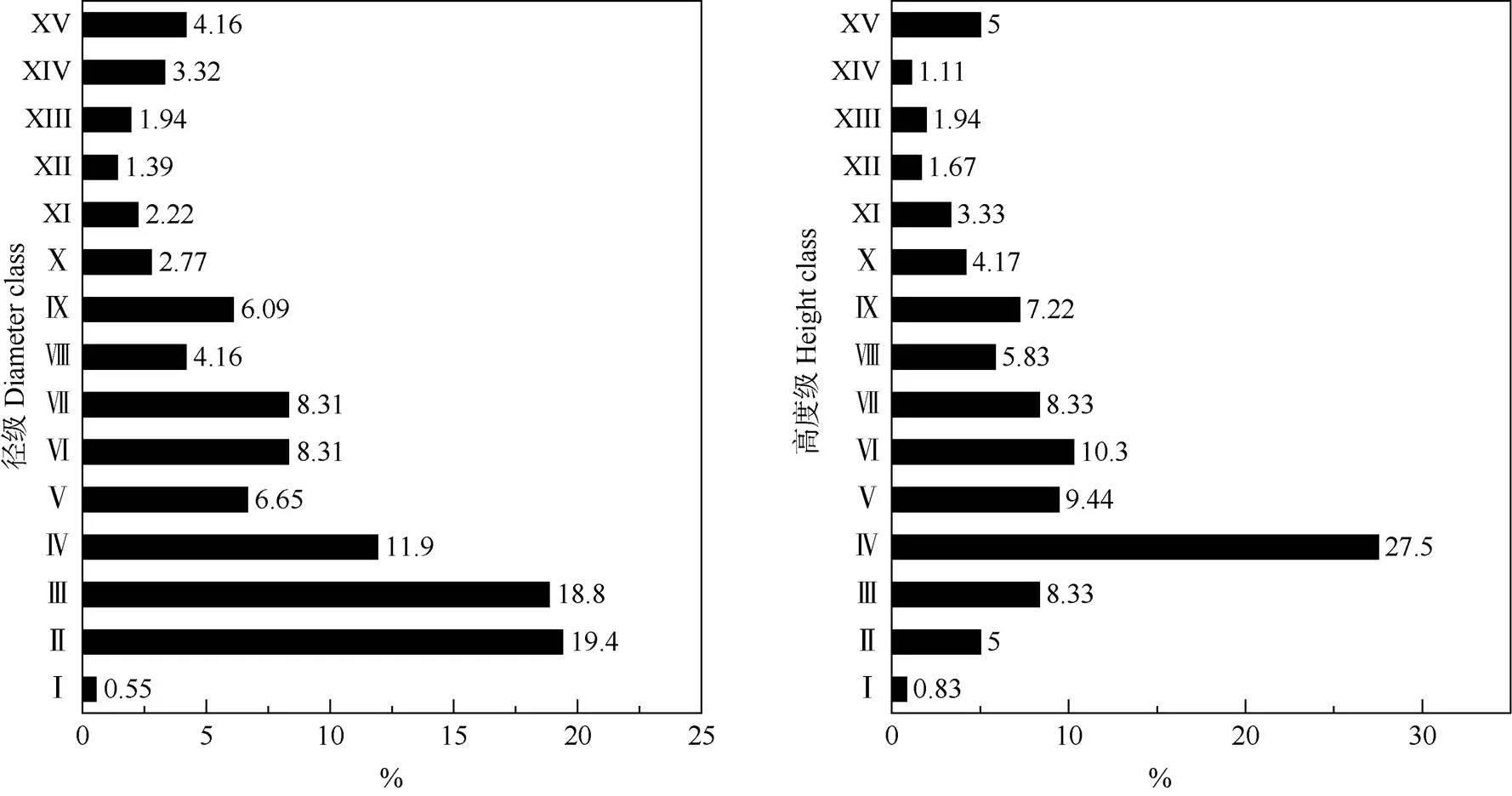
图3 小黄花茶种群的径级和高度级分布。径级: Ⅰ: <0.5 cm; Ⅱ: 0.5~1 cm; Ⅲ: 1~1.5 cm; Ⅳ: 1.5~2 cm; Ⅴ: 2~2.5 cm; Ⅵ: 2.5~3 cm; Ⅶ: 3~3.5 cm;Ⅷ: 3.5~4 cm; Ⅸ: 4~4.5 cm; Ⅹ: 4.5~5 cm; XI: 5~5.5 cm; XII: 5.5~6 cm; XIII: 6~6.5 cm; XIV: 6.5~7 cm; XV: ≥7 cm。高度级: Ⅰ: <0.5 m; Ⅱ: 0.5~1 m; Ⅲ: 1~1.5 m; Ⅳ: 1.5~2 m; Ⅴ: 2~2.5 m; Ⅵ: 2.5~3 m; Ⅶ: 3~3.5 m; Ⅷ: 3.5~4 m; Ⅸ: 4~4.5 m; Ⅹ: 4.5~5 m; XI: 5~5.5 m; XII: 5.5~6 m; XIII: 6~6.5 m; XIV: 6.5~7 m; XV: ≥7 m。以下图表同。
2.4 种群静态生命表
由表2可见,不同龄级小黄花茶的存活个体数波动较大,总体上,随着龄级的增加,个体存活数量呈先上升而后逐渐降低的趋势,植株个体期望寿命降低。从龄级Ⅱ~Ⅴ的存活数随着径级的增加而降低,生命期望值也表现出在幼苗时期(Ⅰ)最高,而随着龄级的增加其生命期望值降低,且在龄级XIV时最低,这说明小黄花茶在幼龄时期的生理活动相对旺盛,老年时期个体生命力变弱。小黄花茶种群个体在龄级XI时死亡数、死亡率以及消失率均为0,说明此时小黄花茶种群对环境的适应能力较强。此外,龄级Ⅰ的死亡率为负数,这说明小黄花茶的幼苗严重不足。静态生命表反映了小黄花茶种群的基本生存状态。
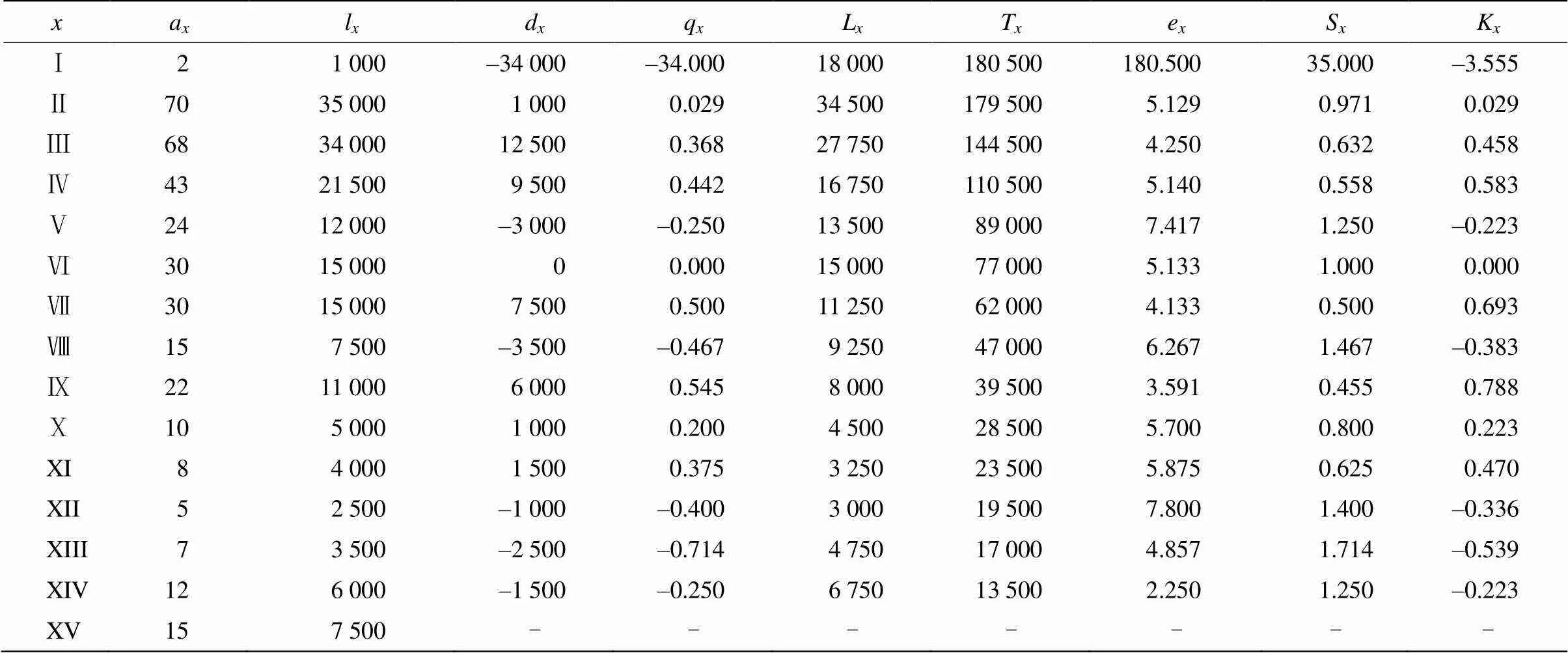
表2 小黄花茶种群的静态生命表
: 龄级;a: 存活数;l: 标准化存活数;d: 死亡数;q: 死亡率;L: 平均存活数;T: 存活总数;e:期望寿命;S: 存活率;K: 消失率。
: Age class;a: Survival number;l: Standardized survival number;d: Mortality;q: Mortality rate;L: Average survival number;T: Total survival number;e: Life expectancy;S: Survival;K: Extinction rate.
2.5 种群生存潜力分析
小黄花茶种群的生存潜力分析见图4。存活曲线表明小黄花茶种群在Ⅰ龄级(<0.5 cm)时存活数量最低,从Ⅱ~XV龄级开始存活率呈现明显下降趋势, 经数学模型检验,种群内线性函数模型的检验值和判定系数R均大于指数函数和幂函数,且具有显著性(表3),可见种群的存活曲线更趋近于Deevey-Ⅰ型。结合死亡率曲线、生命期望值曲线和消失率曲线可以看出,从龄级Ⅰ~Ⅱ,死亡率和消失率急剧升高而生命期望值急剧降低,而从龄级Ⅱ~XV,死亡率和生命期望值趋于平缓,消失率则呈现较大波动,但总体趋于平缓。总的来看,小黄花茶属于衰退型种群,其死亡率、生命期待值以及消失率呈现出较大波动。

表3 小黄花茶种群存活曲线的检验模型

图4 小黄花茶种群的存活曲线(lx)、死亡率曲线(qx)、生命值曲线(ex)和消失率曲线(Kx)
3 结论和讨论
本研究中,小黄花茶数量在8个小生境中仅存活545株,小生境间的数量差异较大,且不同生境条件下生存状况不同。研究表明,地径和高度与树龄呈正相关[22],径级结构、高度级结构是植物个体的生存能力与环境相互作用的结果,其与个体存活数量、生长指标等均可以反映种群与环境的适和度[23–25]。小黄花茶种群地径和高度均呈先升高后降低趋势,年龄结构中幼苗和老年个体数量较少,中年个体数量较多,因此推测出目前小黄花茶种群年龄结构呈衰退趋势。王君等[26]研究也认为小黄花茶幼苗阶段受到人为等环境因素的影响较大,并受其他优势树种的竞争压力,导致其幼苗的数量较少, 种群处于衰退状态。然而,张华雨等[27]研究却认为小黄花茶种群处于稳定类型,一个可能的原因是, 近5 a来,小黄花茶生境受外界干扰增强,种群已经从稳定型转变为衰退型。此外,本研究发现小黄花茶种群在Ⅱ~XV龄级其种群存活数急剧降低,死亡率呈微小平稳波动态势,种群消失率则呈较大的波动。从生命表、存活曲线、死亡率曲线、生命值曲线和消失率曲线中可以看出,小黄花茶种群的存活个体数、死亡率以及生命期望值波动均比较大; 当濒危植物面临强烈的资源竞争以及其它外界因素的影响时,植物幼苗存活率通常很低,种群存活状态差[28–30]。因此,本研究中的小黄花茶可能面临着伴生种(如竹子)强烈的资源竞争和人为干扰(如竹林经营),从而导致幼苗存活率极低,种群处于不稳定状态。此外,幼苗的存活率低也可能与种子萌发率低或被捕食率高有关,如吕晓梅等[14]和刘海燕等[15]通过研究小黄花茶种子特征发现,小黄花茶种子外壳坚硬,种子病虫率较高,从而导致种子出芽率较低,且容易遭遇动物取食,造成自然生境中的种子损失、种源稀少、繁衍困难而使该幼苗存活数量极少[14–15],这些因素都是导致小黄花茶种群衰退的重要原因。
本研究中小黄花茶分布于8个不同的小生境,它们构成了在空间上相互隔离但又存在功能联系的8个局域种群。局域种群间可以通过花粉或种子交流,进而发生功能上的联系[31],8个小生境局域种群组成了该保护区小黄花茶复合种群[32–33]。通常情况下生境面积越大越有利于维持局域种群数量并增加局域种群间迁移率繁殖,促进种群续存[34–35]。小黄花茶的衰退原因也可能是人为干扰导致生境破碎化,各小生境之间相隔距离较远,小黄花茶局域种群交流受阻,从而加剧其灭绝的风险。此外, 局域种群数量衰减,如观音岩谷仅存余24株,进一步降低复合种群数量维持,使小黄花茶可能发展成为极危种群或极小种群。极小种群特指分布地域狭窄, 长期受到外界因素干扰胁迫而呈现种群退化和个体数量持续减少, 以致种群和个体数量已低于稳定存活界限的最小可存活数量,而具有随时面临灭绝风险的种群[36]。然而,小黄花茶至今尚未纳入国家级极小种群物种保护名录。基于本研究结果,我们建议将小黄花茶纳入国家级极小种群物种保护名录。为缓解小黄花茶受竹林经营干扰对种群数量维持的影响,对小黄花茶采取有效措施加强种质资源保护已迫在眉睫,如通过排除竹林经营干扰恢复原生境,采取迁地和就地保护促进种群复壮,采取人工繁育种苗进行种群回归保护等。此外,应加强小黄花茶种群生态学过程研究,重点在种群遗传、种群繁殖过程与机理,种群扩散限制与数量维持, 复合种群生态过程,群落物种多样性,种间关系等,并从分子生物学和生理生态学角度探索种群濒危机制;进一步研发小黄花茶种群生态保护技术体系如迁地和就地保护技术、种苗繁育技术、病虫害防控技术、种群复壮与种群回归技术等。
[1] ZHANG X P, YU L Z, YANG X Y, et al. Population structure and dynamics ofseedlings regenerated from seeds in a montane region of eastern Liaoning Province, China [J]. Chin J Appl Ecol, 2022, 33(2): 289–296. doi: 10.13287/j.1001-9332.202202.001.
张晓鹏,于立忠,杨晓燕,等. 辽东山区天然更新红松幼苗种群结构与动态 [J]. 应用生态学报, 2022, 33(2): 289–296. doi: 10.13287/j. 1001-9332.202202.001.
[2] YU S C, ZHANG W H, LI G, et al. Effects of different crown densities on structure ofpopulations in Huanglong Mountains, northwest China [J]. Acta Ecol Sin, 2017, 37(5): 1537– 1548. doi: 10.5846/stxb201509251971.
于世川,张文辉,李罡,等. 黄龙山林区不同郁闭度对辽东栎种群结构的影响 [J]. 生态学报, 2017, 37(5): 1537–1548. doi: 10.5846/ stxb201509251971.
[3] JIANG Z M, HE Z S, SU H, et al. Population structure and dynamic characteristics of endangeredHemsl. [J]. Acta Ecol Sin, 2018, 38(7): 2471–2480. doi: 10.5846/stxb201704250753.
姜在民,和子森,宿昊,等. 濒危植物羽叶丁香种群结构与动态特征[J]. 生态学报, 2018, 38(7): 2471–2480. doi: 10.5846/stxb201704 250753.
[4] ZHAO Y, CAO X W, LI B, et al. Structural characteristics of 4 natural populations in the southern forest region of Gansu Province [J]. Sci Silv Sin, 2020,56(9): 21–29. doi: 10.11707/j.1001-7488.20200903.
赵阳,曹秀文,李波,等. 甘肃南部林区4种天然林种群结构特征[J]. 林业科学, 2020,56(9): 21–29. doi: 10.11707/j.1001-7488.2020 0903.
[5] LUO X, GUO Q J, YAO L, et al. Characteristics of natural population structure of endangered plant[J]. J CS Univ For Technol, 2021, 41(7): 115–123. doi: 10.14067/j.cnki.1673-923x.2021. 07.014.
罗西,郭秋菊,姚兰,等. 濒危植物鹅掌楸的天然种群结构特征[J]. 中南林业科技大学学报, 2021, 41(7): 115–123. doi: 10.14067/j.cnki. 1673-923x.2021.07.014.
[6] MA D D, KU W P, XIA G H, et al. Structure and dynamics of rare and endangered plantnatural population [J]. J Nanjing For Univ (Nat Sci), 2021, 45(3): 159–164. doi: 10.12302/j.issn.1000-2006. 202006007.
马丹丹,库伟鹏,夏国华,等. 珍稀濒危植物堇叶紫金牛种群结构及动态分析[J]. 南京林业大学学报(自然科学版), 2021, 45(3): 159– 164. doi: 10.12302/j.issn.1000-2006.202006007.
[7] ZHANG H D, ZENG F A., a new section of[J]. Acta Sci Nat Univ Sunyatseni, 1982, 21(3): 74–75.
张宏达,曾范安. 山茶属一新组——小黄花茶组[J]. 中山大学学报(自然科学版), 1982, 21(3): 74–75.
[8] JIANG C H, QI J H, WENG T, et al. Vertical variations of soil nutrients and organic matter incommunity of Danxia landform in north Guizhou [J]. J Mount Agric Biol, 2021, 40(4): 12– 17. doi: 10.15958/j.cnki.sdnyswxb.2021.04.002.
蒋长洪,漆基海,翁涛,等. 丹霞地貌桫椤群落土壤养分及有机质的垂直分异[J]. 山地农业生物学报, 2021, 40(4): 12–17. doi: 10.15958/ j.cnki.sdnyswxb.2021.04.002.
[9] XU D J, GUO N B, WANG P P, et al. Study on the population structure and distribution pattern ofin Xishui Nature Reserve [J]. J SW Univ (Nat Sci), 2014, 36(11): 93–98. doi: 10.13718/ j.cnki.xdzk.2014.11.014.
徐德静,郭能彬,王鹏鹏,等. 习水自然保护区桫椤种群结构与分布格局研究[J]. 西南大学学报(自然科学版), 2014, 36(11): 93–98. doi: 10.13718/j.cnki.xdzk.2014.11.014.
[10] XU D J, WANG P P, HE Y J, et al. Niche characteristics of dominant plant populations ofcommunity of Danxia land- form in north Guizhou [J]. Bull Bot Res, 2014, 34(5): 612–618. doi: 10.7525/j.issn.1673-5102.2014.05.006
徐德静,王鹏鹏,何跃军,等. 黔北丹霞地貌桫椤群落优势种群生态位研究[J]. 植物研究, 2014, 34(5): 612–618. doi: 10.7525/j.issn. 1673-5102.2014.05.006
[11] HE Y J, LIU J M, ZHONG Z C, et al. Intraspecific and interspecific competition ina community [J]. J SW Univ (Nat Sci), 2004, 26(5): 589–593. doi: 10.3969/j.issn.1673-9868.2004.05.019
何跃军,刘济明,钟章成,等. 桫椤群落的种内种间竞争研究[J]. 西南大学学报(自然科学版), 2004, 26(5): 589–593. doi: 10.3969/j.issn. 1673-9868.2004.05.019
[12] BAI X J, ZHANG H Y, WANG X, et al. Flora characteristics of community of endangered plantLi ex H. T. Chang [J]. Guangdong Agric Sci, 2017, 44(1): 94–99. doi: 10.16768/j.issn. 1004-874X.2017.01.014
白小节,张华雨,王鑫,等. 濒危植物小黄花茶群落区系特征研究[J]. 广东农业科学, 2017, 44(1): 94–99. doi: 10.16768/j.issn.1004- 874X.2017.01.014
[13] LIU Q B, LIU B Y, LIANG S. Discussion on the endangered reasons and countermeasures of[J]. Environ Prot Technol, 2005(3): 24–26. doi: 10.3969/j.issn.1674-0254.2005.03.003.
刘清炳,刘邦友,梁盛. 小黄花茶濒危原因及对策探讨[J]. 环保科技, 2005(3): 24–26. doi: 10.3969/j.issn.1674-0254.2005.03.003.
[14] LÜ X M, LIU H Y, HUANG L H, et al. Biological characteristics of five rare and endangeredseeds in Guizhou Province [J]. Guihaia, 2017,37(1): 109–117. doi: 10.11931/guihaia.gxzw201609002.
吕晓梅,刘海燕,黄丽华,等. 山茶属五种稀有濒危植物种子的生物学特性研究[J]. 广西植物, 2017,37(1): 109–117. doi: 10.11931/ guihaia.gxzw201609002.
[15] LIU H Y, ZOU T C, ZHOU H Y, et al. A study on seedling propagation and narrow limited distribution mechanism of 10 endemic species in Guizhou [J]. Guizhou Sci, 2010, 28(4): 11–16. doi: 10.3969/j.issn. 1003-6563.2010.04.002.
刘海燕, 邹天才, 周洪英,等. 10种贵州特有植物的种苗繁殖试验及其分布机理研究[J]. 贵州科学, 2010, 28(4): 11–16. doi: 10.3969/j. issn.1003-6563.2010.04.002.
[16] HE Q Q. The distribution pattern ofY. K. Li population [J]. Environ Prot Technol, 2012, 18(3): 28–30. doi: 10.3969/ j.issn.1674-0254.2012.03.008.
何琴琴. 小黄花茶种群空间分布格局研究[J]. 环保科技, 2012, 18(3): 28–30. doi: 10.3969/j.issn.1674-0254.2012.03.008.
[17] ZHANG T, ZHOU X L, LIU H Y, et al. Study on cottage propagation technology ofY. K. Li [J]. Seed, 2010, 29(4): 86–89. doi: 10.3969/j.issn.1001-4705.2010.04.027.
张婷,周晓玲,刘海燕,等. 小黄花茶扦插繁殖技术研究[J]. 种子, 2010, 29(4): 86–89. doi: 10.3969/j.issn.1001-4705.2010.04.027.
[18] CHEN K Y, ZHANG H R, ZHANG B, et al. Population structure and dynamic characteristics of typical constructive species in natural secondary forest on the northern slope of Changbai Mountain [J]. Acta Ecol Sin, 2021, 41(13): 5142–5152. doi: 10.5846/stxb202101050049.
陈科屹,张会儒,张博,等. 长白山北坡天然次生林典型建群种的种群结构及动态特征[J]. 生态学报, 2021, 41(13): 5142–5152. doi: 10.5846/stxb202101050049.
[19] JIANG H. Population Ecology ofMact [M]. Beijing: China Forestry Publishing House, 1992: 1–175.
江洪. 云杉种群生态学[M]. 北京: 中国林业出版社, 1992: 1–175.
[20] DEEVEY JR E S. Life tables for natural populations of animals [J]. Quart Rev Biol, 1947, 22(4): 283–314. doi: 10.1086/395888.
[21] HETT J M, LOUCKS O L. Age structure models of balsam fir and eastern hemlock [J]. J Ecol, 1976, 64(3): 1029–1044. doi: 10.2307/ 2258822.
[22] LIANG J P, CHENG F, WU M, et al. Distribution of highness and diameter classes and correlation between tree height and DBH ofnatural population in Tianyang, Guangxi [J]. Guangxi For Sci, 2017, 46(2): 134–139. doi: 10.3969/j.issn.1006-1126. 2017.02.003.
梁建平,程飞,吴敏,等. 广西田阳望天树天然种群高度级、径级分布及树高-胸径相关生长关系[J]. 广西林业科学, 2017, 46(2): 134–139. doi: 10.3969/j.issn.1006-1126.2017.02.003.
[23] KANG D, GUO Y, REN C, et al. Population structure and spatial pattern of main tree species in secondaryforest in Ziwuling Mountains, China [J]. Sci Rep-UK, 2014, 4: 6873. doi: 10. 1038/srep06873.
[24] REN Y H, LUO D Q, FANG J P, et al. Structure and dynamics ofvar.population in eastern slope of the Sejila Mountains [J]. J NW Agric For Univ (Nat Sci), 2021, 49(7): 59–68. doi: 10.13207/j.cnki.jnwafu.2021.07.007.
任毅华,罗大庆,方江平,等. 色季拉山东坡急尖长苞冷杉种群结构与动态[J]. 西北农林科技大学学报(自然科学版), 2021, 49(7): 59–68. doi: 10.13207/j.cnki.jnwafu.2021.07.007.
[25] XIE T T, SU P X, ZHOU Z J, et al. Structure and dynamic chara- teristics ofpopulation in the desert-oasis ecotone [J]. Acta Ecol Sin, 2014, 34(15): 4272–4279. doi: 10.5846/ stxb201212101776.
解婷婷,苏培玺,周紫鹃,等. 荒漠绿洲过渡带沙拐枣种群结构及动态特征[J]. 生态学报, 2014, 34(15): 4272–4279. doi: 10.5846/stxb 201212101776.
[26] WANG J, LIU H Y, ZOU T C. Population ecology ofin Guizhou [J]. Guizhou Agric Sci, 2010, 38(11): 59–62. doi: 10.3969/j.issn.1001-3601.2010.11.018.
王君,刘海燕,邹天才. 贵州特有植物小黄花茶的种群生态学研究[J]. 贵州农业科学, 2010, 38(11): 59–62. doi: 10.3969/j.issn.1001- 3601.2010.11.018.
[27] ZHANG H Y, ZONG X H, WANG X, et al. Population structure and living community characteristics of endangeredLi ex H. T. Chang [J]. Plant Sci J, 2016, 34(4): 539–546. doi: 10.11913/ PSJ.2095-0837.2016.40539.
张华雨,宗秀虹,王鑫,等. 濒危植物小黄花茶种群结构和生存群落特征研究[J]. 植物科学学报, 2016, 34(4): 539–546. doi: 10. 11913/PSJ.2095-0837.2016.40539.
[28] YANG H Q, LIU Y Y, LIU F L, et al. Population structure and dynamic characteristics of an endangered and endemic speciessubsp.in southwest China [J]. Acta Bot Boreali-Occid Sin, 2020, 40(12): 2148–2156. doi: 10.7606/j.issn.1000-4025.2020.12.2148.
杨慧琴,刘圆缓,刘芳黎,等. 西南特有濒危植物大王杜鹃种群结构及动态特征[J]. 西北植物学报, 2020, 40(12): 2148–2156. doi: 10.7606/j.issn.1000-4025.2020.12.2148.
[29] LIU J Q, LI N, XIONG T S, et al. Structure and dynamic changes of different populations of rare plantvar.[J]. J Trop Subtrop Bot, 2014, 22(5): 479–485. doi: 10.3969/j.issn.1005-3395. 2014.05.009.
刘佳庆,李宁,熊天石,等. 濒危植物南方红豆杉不同种群的结构和动态变化[J]. 热带亚热带植物学报, 2014, 22(5): 479–485. doi: 10.3969/j.issn.1005-3395.2014.05.009.
[30] YANG H Y, FENG B X, YUAN M Q, et al. Analyses on population pattern and endangered causes of rare and endangered species[J]. Seed, 2013, 32(6): 52–54. doi: 10.3969/j.issn.1001- 4705.2013.06.013.
杨汉远,冯邦贤,袁茂琴,等. 珍稀濒危植物小叶红豆种群格局及濒危原因分析[J]. 种子, 2013, 32(6): 52–54. doi: 10.3969/j.issn. 1001-4705.2013.06.013.
[31] WANG L, GENG Y F, LI Y P, et al. Researches onand its population characteristics: A review [J]. J W China For Sci, 2016, 45(4): 166–173. doi: 10.16473/j.cnki.xblykx1972.2016.04.029.
王磊,耿云芬,李勇鹏,等. 云南红豆杉及其种群特性研究综述[J]. 西部林业科学, 2016, 45(4): 166–173. doi: 10.16473/j.cnki.xblykx 1972.2016.04.029.
[32] LEVINS R. Some demographic and genetic consequences of environ- mental heterogeneity for biological control [J]. Bull Entmol Soc Am, 1969, 15(3): 237–240. doi: 10.1093/besa/15.3.237.
[33] WU J G. What is metapopulation, really? [J]. Chin J Plant Ecol, 2000, 24(1): 123–126.
邬建国. Metapopulation(复合种群)究竟是什么? [J]. 植物生态学报, 2000, 24(1): 123–126.
[34] SONG W X, ZHANG F, LIU R T. The effect of habitat destruction pattern on the dynamics and persistence of metapopulation [J]. Acta Ecol Sin, 2009, 29(9): 4815–4819. doi: 10.3321/j.issn:1000-0933.2009. 09.027.
宋卫信,张锋,刘荣堂. 生境破坏的模式对集合种群动态和续存的影响[J]. 生态学报, 2009, 29(9): 4815–4819. doi: 10.3321/j.issn: 1000-0933.2009.09.027.
[35] ZHOU S R, WANG G. The influence of the number of reserves on population persistence at the metapopulation level [J]. J Lanzhou Univ (Nat Sci), 2002, 38(4): 109–113. doi: 10.3321/j.issn:0455-2059.2002. 04.021.
周淑荣,王刚. 保护区的数量和种群在集合种群水平上的续存[J]. 兰州大学学报, 2002, 38(4): 109–113. doi: 10.3321/j.issn:0455-2059. 2002.04.021.
[36] ZANG R G, DONG M, LI J Q, et al. Conservation and restoration for typical critically endangered wild plants with extremely small popu- lation [J]. Acta Ecol Sin, 2016, 36(22): 7130–7135. doi: 10.5846/stxb 201610082011.
臧润国,董鸣,李俊清,等. 典型极小种群野生植物保护与恢复技术研究[J]. 生态学报, 2016, 36(22): 7130–7135. doi: 10.5846/stxb 201610082011.
Population Structure and Survival Potentiality Analysis of Endangered
BAI Xiaojie1, SHEN Kaiping2, MU Jun2, WENG Tao2, ZANG Lipeng2, REN Wenda2, HAN Xu2, LI Qing2, TAN Qiyu2, HE Yuejun2*
(1. Management Bureau of Chishui Alsophila Spinulosa National Nature Reserve,Chishui 564700, Guizhou, China; 2. Forestry College, Forest Ecology Research Center, Guizhou University,Guiyang 550025, China)
In order to understand the growth characteristics and survival status of the endangered, an inventory ofresources was carried out in ChishuiNational Nature Reserve, Guizhou Province. The height and ground diameter of each plant were investigated by sampling method, and the population number and structural characteristics were statistically analyzed. The population was divided into specific age groups with 0.5 cm diameter interval and classified into 5 diameter class from I to XV. A static life table was drawn to analyze the survival potential and development trend of the population. The results showed that a total of 545 individuals ofwere found in the survey, the number varied greatly in different niches, ranging from 24 to 120. The number of individuals with ground diameter less than 0.5 cm and height less than 0.5 m were very few, and the maximum population was maintained with average ground diameter of 0.5-2 cm. The mortality and extinction rates of I-II age class (0-1.0 cm) population increased sharply, while the life expectancy decreased sharply. The mortality and life expectancy of II-XV age class (≥0.5 cm) population tended to be flat, while the extinction rates fluctuated greatly but generally tended to be flat. The population structure and survival curve showed that the population was close to Deevey-I type, belonging to the declining population. Therefore, the seedling number ofpopulation was seriously insufficient, the population self-renewal was difficult, the existing population interference pressure was large, showing obvious population decline trend, it was urgented to strengthen the population conservation.
;Population structure; Static life table; Survival curve; Endangered plant
10.11926/jtsb.4548
2021-10-25
2021-12-03
国家自然科学基金项目(31660156);贵州省计划项目([2021]455);贵州省高层次创新型人才平台项目([2020]6004,[2017]5788,[2018]5781);贵州省生态学重点学科建设项目(ZDXK[2016]7);赤水小黄花茶专项调查项目(2021)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 31660156); the Planning Project in Guizhou (Grant No. [2021]455), the Talent-platform Program of Guizhou (Grant No. [2020]6004, [2017]5788, [2018]5781); the Project for Key Subject Construction of Ecology in Guizhou (Grant No. ZDXK [2016]7); and the Special Project forSurvey in Chishui (Grant No. 2021).
白小节(1980生),男,工程师,主要从事自然保护区珍稀物种保护研究。E-mail: gzcsbxj@163.com
. E-mail: hyj1358@163.com
